飼料中類胡蘿卜素在大黃魚皮膚中的沉積及其分離鑒定
2019-01-02 01:18:02申豪豪易新文張文兵麥康森
飼料工業 2018年10期
■申豪豪 牟 華 李 俊 易新文 張文兵麥康森
(水產動物營養與飼料農業部重點實驗室海水養殖教育部重點實驗室中國海洋大學水產學院,山東青島266003)
體色不僅是養殖魚類品質的一個重要指標,同時也反映了魚類的健康狀況,影響魚類的商品價值和市場認可度。大黃魚(Larimichthys crocea)是我國重要的海水養殖魚類,2016年我國人工養殖大黃魚產量已達16.54萬噸,居海水養殖魚類第一。野生大黃魚皮膚金黃,唇部為紅色,深受消費者的喜愛。然而,高密度養殖大黃魚的體色嚴重退化,降低了消費者的認可度,其市場售價與野生大黃魚相差幾十倍甚至上百倍。
影響魚類體色的因素有很多,如光照、溫度、背景色及營養物質等,其中類胡蘿卜素是影響魚類體色的主要因素之一[1-4]。魚類像其它脊椎動物一樣,不能從頭合成類胡蘿卜素,必需從食物中攝取[5]。目前,在魚類體內發現多種類胡蘿卜素,如蝦青素、黃體素、玉米黃質、金槍魚黃素等[6-7]。飼料中添加適宜的類胡蘿卜素,對于改善魚類體色有著重要作用。不同魚類體內的類胡蘿卜素組成存在差異,因而飼料中添加的有效呈色作用的類胡蘿卜素有所不同。研究發現蝦青素是鮭鱒魚體內的主要色素,如大馬哈魚(Salmosalar)[8]、細鱗大麻哈魚(Oncorhynchusgorbuscha)[9]和大西洋鮭(Salmo salar)[10]等。因此,蝦青素和各種蝦青素源被廣泛用于提升鮭鱒魚肌肉的紅色值[11-14]。
魚類類胡蘿卜素組成不同,這可能是由于類胡蘿卜素代謝差異所造成的。Schiedt等[15]報道在虹鱒(Oncorhynchus mykiss)中部分蝦青素雙酯被代謝為玉米黃質。Schiedt[16]發現鯉科魚類能夠通過氧化修飾作用將玉米黃質轉化為蝦青素。Tsushima等[17]發現在日本方頭魚(Branchiostegus japonicas)中,蝦青素被代謝為金槍魚黃素。Gomes等[18]也報道發現在金頭鯛(Sparusaurata)中,蝦青素被代謝為黃體素酯和表黃體素酯。Yi等[19-20]研究了飼料中添加不同類胡蘿卜素(蝦青素、黃體素、角黃素)對大黃魚體色的影響,結果發現,這些類胡蘿卜素的使用都可改善大黃魚的體色,然而效果存在差異,其中黃體素具有更好的著色效果,說明黃體素可能是大黃魚皮膚中的主要色素。但有關大黃魚皮膚中類胡蘿卜素的組成及代謝情況卻仍然不清楚。
本研究選擇蝦青素和黃體素為實驗類胡蘿卜素添加到配合飼料,對大黃魚進行養殖實驗,研究其對大黃魚皮膚呈色的影響,并對皮膚中的類胡蘿卜素各組分進行分離和鑒定,為通過飼料營養途徑改善養殖大黃魚的體色提供基礎數據。
1 材料與方法
1.1 試劑與儀器
甲醇和甲基叔丁基醚(MTBE)為色譜純,正己烷、丙酮、乙醚、氫氧化鉀和二叔丁基對甲酚(BHT)均為國產分析純,水為超純水。
安捷倫1100高效液相色譜儀(配備G1313A二元泵,G1313A高效自動進樣器,G1316A柱溫箱,G1315ADAD檢測器),美國安捷倫科技有限公司。
1.2 養殖實驗
以無類胡蘿卜素添加的飼料為對照組(CD組),在此基礎上分別添加80 mg/kg的蝦青素(帝斯曼,荷蘭)和黃體素(廣州智特奇生物科技有限公司,中國),分別命名為AST80和X80飼料組。飼料配方及營養水平見表1。

表1 飼料配方及營養水平(干重)
養殖實驗用大黃魚購自寧德市富發水產有限公司,于4.0 m×4.0 m×4.0 m的海水網箱中暫養2周,暫養期投喂對照組飼料。養殖實驗開始前,所有大黃魚饑餓24 h,使用1∶10 000稀釋的丁香酚麻醉。選取規格一致的大黃魚,稱重并隨機分配到9個2.0 m×2.0 m×2.5 m的網箱中,每個網箱放養50尾。養殖實驗期間,每天飽食投喂兩次(6:00和18:00),養殖周期9周,水溫18~28 ℃,鹽度28~32 g/l,溶氧6.2~8.3 mg/l。
1.3 樣品采集與體色測定
養殖實驗結束時,所有大黃魚饑餓24 h,并用1∶10 000稀釋的丁香酚麻醉,然后取樣。每個網箱中隨機抽10尾魚取皮膚,皮膚樣品用鋁箔包裹保存于-80℃冰箱中。
大黃魚體色按照國際發光照明委員會CIE(CIE,1976)的規定測定,L*代表亮度值、a*代表紅色值、b*代表黃色值。使用CR-400型色彩色差計(柯尼卡-美能達,日本)于夜間20:00至24:00進行測定,避免外源性光照對大黃魚體色的影響。每個網箱隨機取5尾魚,在背部和腹部皮膚分別選取2個點測定體色值,盡量保證檢測部位的一致性。
1.4 大黃魚皮膚中類胡蘿卜素的分析
大黃魚皮膚中類胡蘿卜素的分析參考相關方法并進行改良[18,21-24]。準確稱取0.25 g大黃魚皮膚于離心管中,加入10 ml丙酮,使用勻漿機勻漿1 min。將離心管在4℃、5 000×g轉速的離心機中離心5 min,吸取上層提取液,再加入丙酮,反復萃取直到無色。合并萃取液,加入4 ml甲醇-正己烷(1∶1,v/v)混合提取液和2 ml超純水,震蕩離心管,靜置分層后吸取上層正己烷層。在正己烷層萃取液加入4 ml 30%KOH-甲醇(w/v)溶液,充入氮氣,于40℃避光皂化反應1 h。皂化結束后加入4 ml 3%NaCl(w/v)溶液,將離心管在4℃、轉速為5 000×g的離心機中離心5 min,吸取上層正己烷層萃取液,用超純水清洗,直到清洗液為中性;在剩余提取液中加入4 ml乙醚,將離心管在4℃、5 000×g轉速的離心機中離心5 min,吸取上層乙醚提取液,用超純水清洗,直到清洗液為中性;合并正己烷和乙醚萃取液,氮氣吹干,溶于1 ml甲醇(0.05%BHT)中,然后進行HPLC分析。
1.5 統計分析
所有數據采用SPSS19.0軟件進行單因素ANOVA分析,每個處理3個重復,結果以“平均值±標準誤”來表示。當P<0.05時,認為有顯著性差異,并進行Tukey's多重比較。
2 結果
飼料中蝦青素和黃體素對大黃魚皮膚呈色的影響結果見表2。各處理組在背部皮膚亮度值、腹部皮膚亮度值和背部皮膚紅色值上沒有顯著性差異(P>0.05)。在腹部皮膚紅色值方面,類胡蘿卜素添加組高于對照組,最高值出現在蝦青素組,然而無顯著性差異(P>0.05)。在背部皮膚黃色值方面,黃體素組顯著高于對照組和蝦青素組(P<0.05),蝦青素組和對照組間無顯著性差異(P>0.05)。在腹部皮膚黃色值方面,黃體素組和蝦青素組顯著高于對照組(P<0.05),然而黃體素組和蝦青素組間無顯著性差異(P>0.05)。
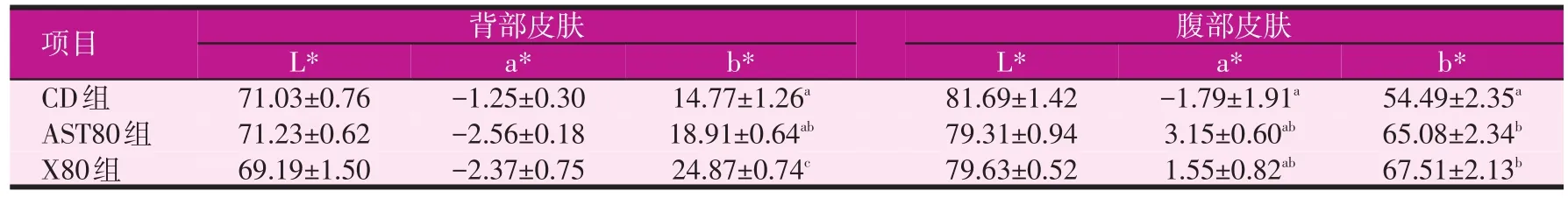
表2 飼料中蝦青素和黃體素對大黃魚體色的影響
飼料中蝦青素和黃體素對大黃魚皮膚類胡蘿卜素含量和各組分比例的影響結果見表3。類胡蘿卜素添加組大黃魚皮膚中類胡蘿卜素的總含量顯著高于對照組(P<0.05),黃體素添加組顯著高于蝦青素添加組(P<0.05)。在所有處理組中,黃體素占大黃魚皮膚中類胡蘿卜素比例最高,在53.04%~67.55%之間,金槍魚黃素次之(30.87%~43.76%);全反式玉米黃質所占比例最低(1.01%~3.20%)。
飼料中蝦青素和黃體素對大黃魚皮膚類胡蘿卜素組分的影響見圖1。對照組和色素添加組間類胡蘿卜素組分無差異,共有7種類胡蘿卜素被檢測到。雖然AST80組添加了蝦青素,然而卻在所有處理組的大黃魚皮膚中均沒有檢測到蝦青素。
3 討論
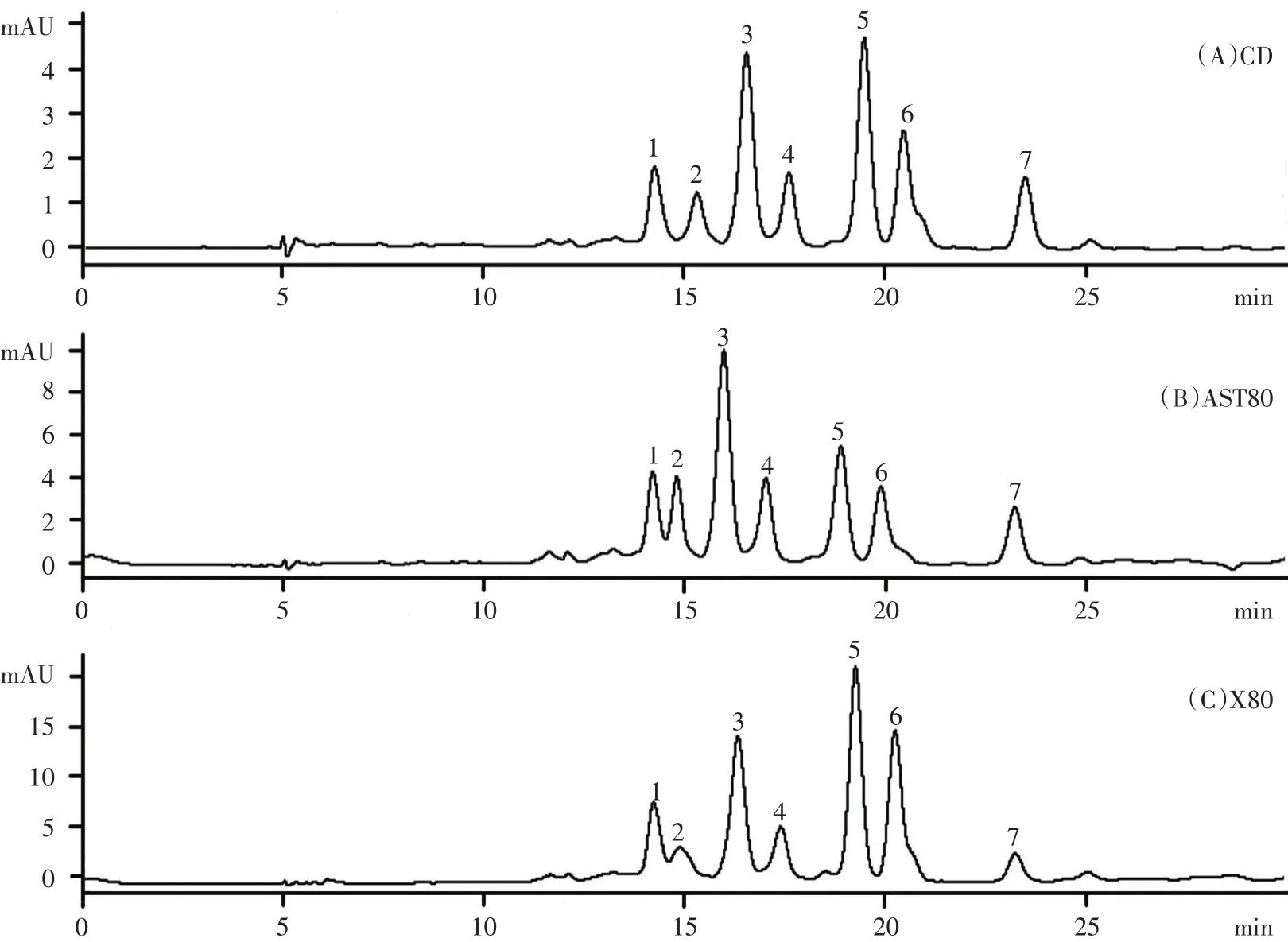
圖1 不同處理組大黃魚皮膚中類胡蘿卜素組分的液相色譜圖
在本研究中,各處理組大黃魚在背部皮膚亮度值和腹部皮膚亮度值上沒有顯著性差異。Kalinowski等[25]報道真鯛皮膚亮度值不受飼料中類胡蘿卜素含量的影響。Yi等[19-20]在大黃魚中也有類似發現。通常情況下,蝦青素被認為是紅色色素用于魚類的色素沉積。Chatzifotis等[26]發現蝦青素能夠提高真鯛皮膚的紅色值,相似的結果在牙鲆(Paralichthysolivaceus)[27]和玫瑰鯽(Pethiaconchonius)[28]中也被發現。然而,在本研究中,各處理組大黃魚背部皮膚紅色值和腹部皮膚紅色值沒有顯著性差異,可能的原因是蝦青素被轉化為其它的色素。黃體素是黃色的色素,被廣泛用于提高魚類皮膚的黃色值。在本研究中,黃體素組大黃魚背部皮膚黃色值顯著高于對照組和蝦青素組。在腹部皮膚黃色值方面,黃體素組和蝦青素組顯著高于對照組。Chatzifotis等[26]報道黃色的類胡蘿卜素(黃體素和玉米黃質)能夠顯著的提高真鯛的黃色值。本研究顯示黃體素是大黃魚皮膚中的主要色素(見表3),然而,在腹部黃色值方面,蝦青素組和黃體素組間沒有顯著的差異,這表明在腹部黃色值方面可能存在一個飽和度。
在本研究中,黃體素組大黃魚皮膚中的類胡蘿卜素含量比對照組和蝦青素組高,這表明與蝦青素相比,大黃魚能夠更好的利用黃體素,這個結果與Yi等[19]的研究結果一致。Li等[29]也發現斑點叉尾鮰(IetalurusPunetaus)皮膚中主要沉積黃體素和玉米黃質而不是蝦青素和角黃素。然而,相關研究發現,與角黃素相比,大西洋鮭能夠更好的利用蝦青素,相似的研究結果在銀大馬哈魚(Oncorhynchuskisutch)上也被發現[30]。可能的原因是魚類對類胡蘿卜素的利用上具有物種差異性。蝦青素是鮭魚的主要色素,然而,本研究表明黃體素是大黃魚皮膚中的主要色素,大黃魚更傾向于沉積黃體素在皮膚中。
在本研究中,各個處理組的大黃魚皮膚中類胡蘿卜素的組分沒有差異,且均沒有檢測到蝦青素,包括蝦青素飼料添加組,這顯示大黃魚不沉積蝦青素,并可能將吸收的蝦青素轉化為其它類胡蘿卜素。魚類含有各種各樣的類胡蘿卜素[6,15,31],并能通過對雙鍵的氧化、還原及裂解作用等修飾它們[7]。Tsushima等[17]報道日本方頭魚通過還原作用將蝦青素經4-酮基玉米黃質、玉米黃質、4-酮基黃體素和黃體素轉化為金槍魚黃素。Gomes等[21]發現在金頭鯛中,蝦青素被代謝為黃體素酯和表黃體素酯。同樣地,Maoka[7]也報道了鱸形目魚類可以將蝦青素經金盞花黃素、玉米黃質和ε-ε-胡蘿卜素-3,3'-二酮最終轉化為金槍魚黃素。蝦青素是鮭魚中的主要色素,然而,Matsuno[6]報道鮭魚不能通過代謝修飾作用從其它類胡蘿卜素合成蝦青素。與此相反,Schiedt[16]發現鯉科魚類能夠通過氧化修飾作用將玉米黃質轉化為蝦青素。這顯示魚類在類胡蘿卜素代謝上存在差異,可能是由于物種不同導致。在本研究中,黃體素占大黃魚皮膚中類胡蘿卜素的比例最高(53.04%~67.55%),其次是金槍魚黃素(30.87%~43.76%),玉米黃質含量最低(1.01%~3.20%)(見表3)。Yi等[19]發現大黃魚皮膚的黃色值隨著飼料中黃體素/蝦青素比值的降低而降低,推斷大黃魚能夠在一定程度上將蝦青素轉化為黃色的色素。綜合本研究和已發表的相關數據,可以推測在大黃魚中蝦青素可能經黃體素和玉米黃質最終被轉化為金槍魚黃素。然而,本研究的結果只是來自大黃魚的皮膚,還缺乏肌肉或其他組織色素沉積的數據,有關上述推測還需要進一步的研究。
4 結論
本研究表明,蝦青素和黃體素均能有效改善大黃魚的體色,大黃魚能夠更好的利用黃體素,并可能將蝦青素經黃體素和玉米黃質最終轉化為金槍魚黃素。
