不同來源山羊體細胞重編程效率比較
2017-05-30 10:35:43宋輝李卉王鋒
安徽農(nóng)業(yè)科學 2017年31期
宋輝 李卉 王鋒
摘要 [目的]檢測克隆奶山羊(♀12003)和轉基因克隆奶山羊(♂12001)作為供體細胞,對體細胞重編程效率的影響。[方法]采用慢病毒作為載體,攜帶多能因子來感染正常山羊(g2)、♀12003和♂12001體細胞,統(tǒng)計克隆形成率和堿性磷酸酶陽性克隆率。[結果] 與普通山羊(g2)相比,轉基因克隆奶山羊耳成纖維細胞的重編程效率低,出現(xiàn)的細胞克隆在傳代之前容易分化;克隆奶山羊體細胞與普通山羊體細胞的重編程效率接近,轉基因克隆奶山羊(♂12001)的重編程效率極顯著低于克隆奶山羊(♀12003)和普通山羊(P<0.01)。[結論]轉基因克隆奶山羊體細胞可以被重編程為誘導性多能干細胞(Induced pluripotent stem cells,iPSCs),但效率較低。
關鍵詞誘導性多能干細胞;重編程;克隆羊
中圖分類號S827文獻標識碼A文章編號0517-6611(2017)31-0115-03
Abstract[Objective] To study the effects of cloned dairy goat (♀12003) and transgenic cloned dairy goat (♂12001) as donor cells on somatic cell reprogramming efficiency. [Method]The lentivirus was used as carrier to carry the pluripotent factor to infect the somatic cells of normal goat (g2), ♀12003 and ♂12001. And the clonal formation rate and alkaline phosphatase positive clone rate were recorded. [Result] Compared with the normal goat (g2), the reprogramming efficiency of ear fibroblasts of the transgenic cloned dairy goat was lower, and the cell clones appeared to be easily differentiated before passage. The reprogramming efficiency of cloned dairy goat′s somatic cells was close to that of the normal goat′s somatic cell. The reprogramming efficiency of transgenic cloned dairy goat (♂ 12001) was significantly lower than that of cloned dairy goat (♀12003) and common goat (P<0.01). [Conclusion] Transgenic cloned dairy goat′s somatic cells could be reprogrammed into induced pluripotent stem cells (iPSCs), but the efficiency was low.
Key wordsInduced pluripotent stem cells;Reprogramming;Cloned goat
轉基因技術是研究基因功能的常用方法之一,在建立人類遺傳疾病模型、動物品種改良、藥用蛋白生產(chǎn)等領域也具有極大的開發(fā)價值。目前常用的生產(chǎn)轉基因動物的方法效率很低,工作繁鎖,周期長。以胚胎干細胞(Embryonic stem cells,ESCs)作為核供體細胞,通過核移植技術生產(chǎn)轉基因動物,可以極大地提高成功率。然而,由于開始使用的材料有各種問題,比如胚胎自身的原因、在體外對ESCs開始的處理以及對ESCs后期的培養(yǎng)條件等因素的影響,偶蹄類動物嚴格意義上的ESCs系一直無法成功建立。這些都制約了偶蹄類家畜ESCs的研究及應用。
誘導性多能干細胞(iPSCs)技術的產(chǎn)生為研究偶蹄類動物的多能干細胞提供了一個很好的思路。豬、牛、綿羊和山羊的iPSCs相繼建系成功[1-13]。南京農(nóng)業(yè)大學江蘇省肉羊產(chǎn)業(yè)工程技術研究中心已經(jīng)成功獲得批量轉基因克隆奶山羊,經(jīng)檢測發(fā)現(xiàn)轉人乳鐵蛋白(hLF)基因已經(jīng)很好地整合到這些克隆奶山羊體細胞的基因組中。筆者用克隆奶山羊和轉基因克隆奶山羊耳成纖維細胞作為體細胞,經(jīng)過慢病毒感染進行重編程,統(tǒng)計克隆形成率和堿性磷酸酶陽性克隆率,旨在為今后使用轉基因克隆奶山羊iPSCs作為供體細胞通過核移植技術高效率生產(chǎn)轉基因克隆奶山羊奠定基礎。
1材料與方法
1.1主要試劑
DMEM培養(yǎng)基、DMEM/F12培養(yǎng)基、胎牛血清(FBS)、非必需氨基酸(NEAA)、雙抗、無鈣鎂PBS緩沖液、胰蛋白酶、谷氨酰胺、β-巰基乙醇(β-ME)均購自美國Life公司;絲裂霉素-C(MIT-C)購自瑞士Roche公司;成纖維細胞生長因子(β-FGF)購自美國Peprotech公司;堿性磷酸酶染色試劑盒購自美國Millipore公司。
1.2方法
1.2.1hLF基因的檢測。提取山羊♂12001細胞DNA,采用PCR方法對其進行鑒定,PCR反應體系(50 μL)如下:10×PCR Buffer 5 μL,10 mmol/L dNTP Mixture 8 μL,上、下游引物各20 pmol,LA Taq DNA聚合酶0.5 μL,25 mmol/L Mg2+ 4 μL,cDNA產(chǎn)物2 μL,補加ddH2O至50 μL。PCR反應程序如下:94 ℃變性5 min;94 ℃變性15 s,60 ℃退火30 s,72 ℃延伸1 min,40個循環(huán);最后72 ℃延伸5 min。設計1對引物:F為5′-GAATGGCTGGCAGTGAAACA-3′,R為5′-CTCAATGGGCTCAGGTGGAC-3′。
1.2.2
hLF基因克隆至T載體。回收RT-PCR擴增產(chǎn)物,進行純化后,克隆在pMD-19T載體的T位點。純化后的PCR產(chǎn)物4 μL,Buffer 2 μL,pMD-19T 1 μL,T4 DNA連接酶1 μL,加水至10 μL,16 ℃過夜。將連接產(chǎn)物轉化感受態(tài)細胞TOP10,隨機挑取單克隆,擴增后進行PCR。將鑒定正確的重組質粒進行測序分析。
1.2.3堿性磷酸酶染色。堿性磷酸酶檢測使用密理博公司的堿性磷酸酶染色試劑盒,按照說明書操作,具體步驟如下:吸棄6孔板中的培養(yǎng)基,PBS清洗3次;加入4%多聚甲醛溶液固定2 min;1 × Rinsebuffer清洗2次;加入AP染色液避光孵育15 min(AP染色液現(xiàn)用現(xiàn)配,F(xiàn)astRed Violet∶Naphthol-BIphosphatesolution∶H2O=2∶1∶1)。吸棄染色液,1×Rinsebuffer 清洗2次。加入PBS,拍照并計算誘導效率。
1.2.4
比較不同來源山羊耳成纖維細胞重編程產(chǎn)生克隆效率和克隆質量。將病毒感染后的體細胞用胰蛋白酶消化鋪在鋪有飼養(yǎng)層的細胞培養(yǎng)皿上,每個皿鋪1×105個細胞,每組做3個重復孔。12~15 d后,在顯微鏡下統(tǒng)計每個皿出現(xiàn)的總克隆數(shù),隨后對每個皿進行AKP染色。統(tǒng)計每個皿中AKP陽性克隆數(shù),最后統(tǒng)計總克隆數(shù)和AKP陽性克隆數(shù)占鋪在對應培養(yǎng)皿上感染細胞數(shù)的比例。
2結果與分析
2.1細胞轉基因鑒定
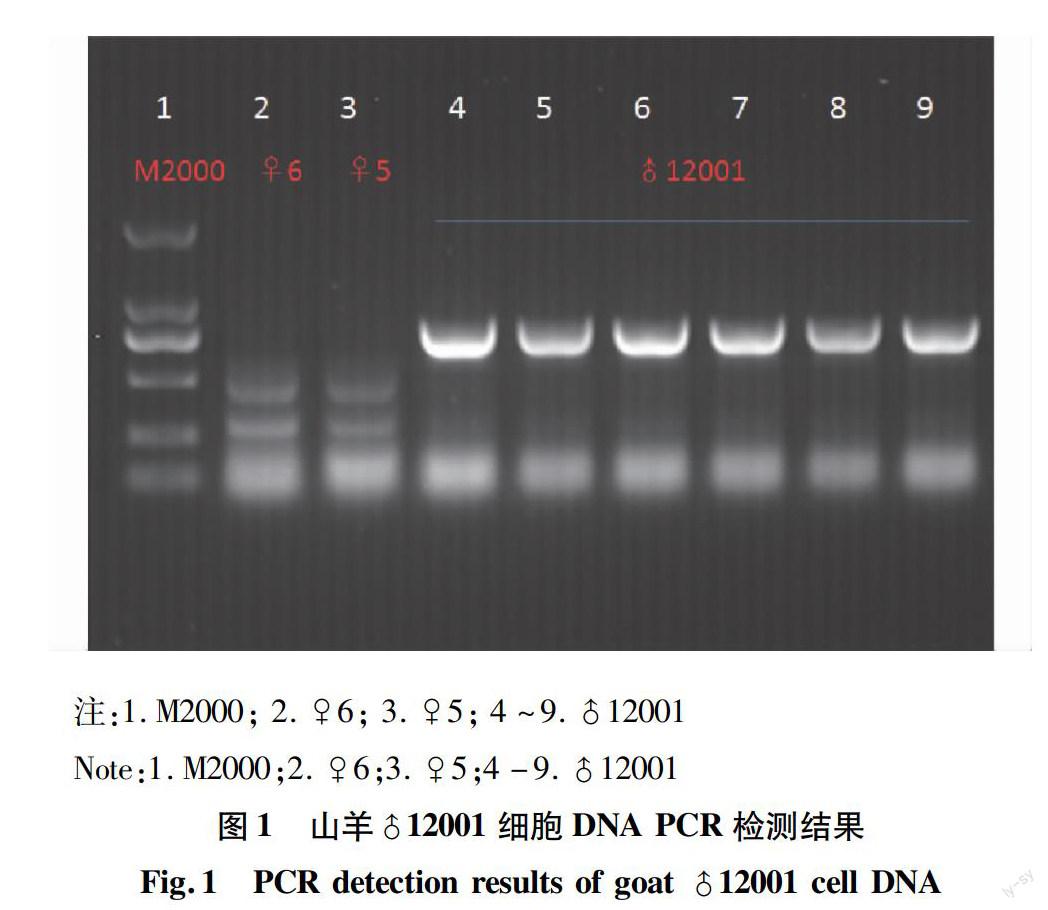
通過PCR檢測山羊♂12001細胞DNA,結果表明通過PCR可以檢測到目的基因hLF,♀5和♀6為普通奶山羊,作為陰性對照(圖1),測序結果表明PCR擴增的片段與hLF基因序列相一致。
2.2堿性磷酸酶(AKP)染色
通過AKP檢測,在顯微鏡下
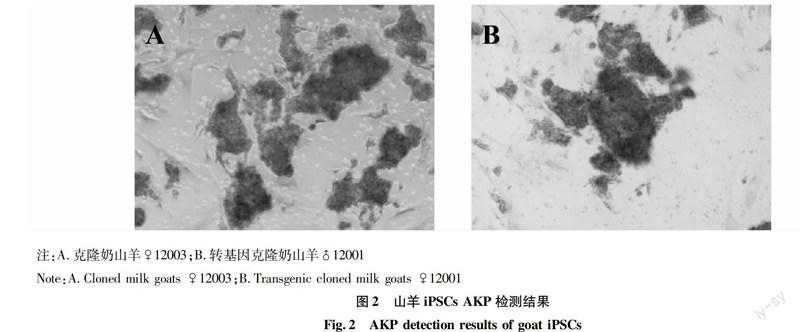
可觀察到細胞的胞漿內(nèi)有紅棕色顆粒,說明克隆奶山羊(♀12003)和轉基因克隆奶山羊(♂12001)iPSCs對AKP染色呈陽性(圖2)。
2.3不同來源山羊耳成纖維細胞重編程產(chǎn)生克隆效率和克隆質量的比較
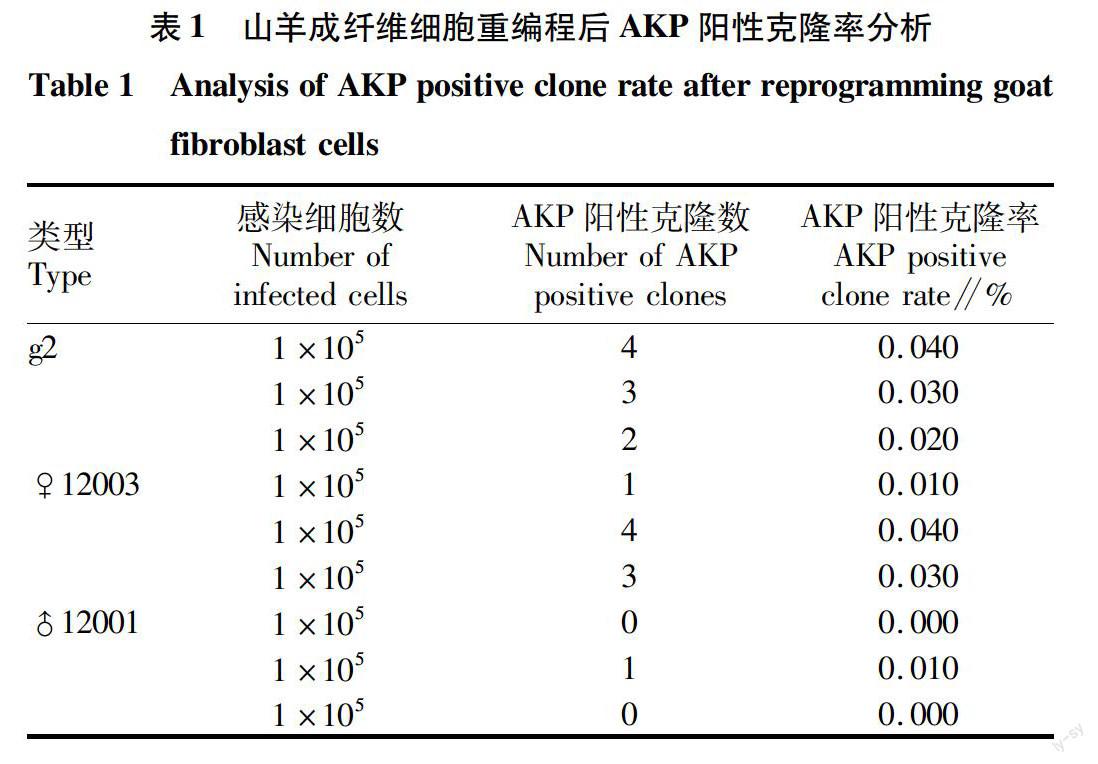
與普通山羊相比,轉基因克隆奶山羊耳成纖維細胞的誘導效率低,出現(xiàn)的細胞克隆在傳代之前容易分化(有的在克隆剛產(chǎn)生的時候就開始分化)。在誘導效率和克隆質量方面,轉基因克隆奶山羊(♂12001)3次重復試驗的平均陽性AKP克隆率為(0.003±0.006)%,顯著低于克隆奶山羊[(0.027±0.015)%]和地方品種山羊[(0.030±0010)%](表 1,圖3,P<0.01)。從圖3可以看出,普通山羊(g2)和克隆奶山羊(♀12003)重編程效率均極顯著高于轉基因克隆奶山羊(♂12001)(P< 0.01),普通山羊(g2)和克隆奶山羊(♀12003)重編程效率沒有顯著差異(P>0.05)。
3結論與討論
ESCs替代普通體細胞作為核供體細胞,可以有效提高SCNT的效率,從而提高偶蹄類家畜的生產(chǎn)特性[14-17]。但是,偶蹄類家畜ESCs一直沒有建系成功。iPSCs是一個建立偶蹄類家畜多能干細胞系的新方法。因此,iPSCs可以代替
ESCs作為供體細胞進行核移植,提高克隆率。許多家畜的
iPSCs已經(jīng)建立成功,包括山羊[1]、綿羊[2-4]、豬[8-9,11,13]和
牛[5-7]。研究人員已經(jīng)通過核移植技術,使用小鼠和豬的iPSCs作為供體細胞獲得相應的克隆動物[10,18-19]。由此可見,iPSCs為生產(chǎn)轉基因克隆動物提供了一個有用的工具。筆者所在實驗室已經(jīng)通過體細胞核移植技術,使用帶有人乳鐵蛋白(hLF)基因的奶山羊皮膚成纖維細胞作為供體細胞,成功獲得轉基因克隆奶山羊,但效率很低[20]。在轉基因過程中,外源基因整合到體細胞的基因組中;在體外,因為核移植的操作對細胞產(chǎn)生傷害和應激等,會導致轉基因克隆動物發(fā)生異常。因此,該研究使用慢病毒為載體將轉基因克隆奶山羊體細胞重編程為iPSCs,作為下一步生產(chǎn)克隆羊的供體細胞,以期提高克隆羊的克隆率及克隆質量。
iPSCs有多種方法來評估重編程的效率,從實際操作角度來看,大多數(shù)研究人員使用克隆計數(shù)這種方法統(tǒng)計重編程的效率[21]。該研究采用堿性磷酸酶活性染色與克隆計數(shù)相結合的方法統(tǒng)計重編程效率,獲得了更為準確的山羊成纖維細胞的重編程效率。使用轉基因克隆奶山羊耳成纖維細胞(tgFs)重編程后獲得的AKP陽性克隆數(shù)顯著低于使用奶山羊耳成纖維細胞重編程后獲得的AKP陽性克隆數(shù)(P<001)。與普通奶山羊耳成纖維細胞相比,使用tgFs進行重編程要困難的多。大多數(shù)tgFs重編程后產(chǎn)生克隆會發(fā)生快速的分化,這可能與轉基因羊的供體細胞經(jīng)過基因改造,外源基因整合到體細胞的基因組中,導致其重編程機制發(fā)生改變造成的。
參考文獻
[1] REN J T,PAK Y J,HE L X Z,et al.Generation of hircineinduced pluripotent stem cells by somatic cell reprogramming[J].Cell Res,2011,21:849-853.
[2] BAO L,HE L X Z,CHEN J J,et al.Reprogramming of ovine adult fibroblasts to pluripotency via druginducible expression of defined factors[J].Cell Res,2011,21(4):600-608.
[3] LI Y,CANG M,LEE A S,et al.Reprogramming of sheep fibroblasts into pluripotency under a druginducible expression of mousederived defined factors[J].PLoS One,2011,6:1-8.
[4] LIU J,BALEHOSUR D,MURRAY B,et al.Generation and characterization of reprogrammed sheep induced pluripotent stem cells[J].Theriogenology,2012,77(2):338-346.
[5] DENG Y F,LIU Q Y,LUO C,et al.Generation of induced pluripotent stem cells from buffalo(Bubalus bubalis)fetal fibroblasts with buffalo defined factors[J].Stem Cells Dev,2012,21(13):2485-2494.
[6] HAN X P,HAN J Y,DING F R,et al.Generation of induced pluripotent stem cells from bovine embryonic fibroblast cells[J].Cell Res,2011,21:1509-1512.
[7] HUANG B,LI T,ALONSOGONZALEZ L,et al.A virusfree polypromoter vector induces pluripotency in quiescent bovine cells under chemically defined conditions of dual kinase inhibition[J].PLoS One,2011,6(9):1-14.
[8] ESTEBAN M A,XU J Y,YANG J Y,et al.Generation of induced pluripotent stem cell lines from Tibetan miniature pig[J].J Biol Chem,2009,284(26):17634-17640.
[9] EZASHI T,TELUGU B P,ALEXENKO A P,et al.Derivation of induced pluripotent stem cells from pig somatic cells[J].Proc Natl Acad Sci USA,2009,106(27):10993-10998.
[10] FAN N N,CHEN J J,SHANG Z C,et al.Piglets cloned from induced pluripotent stem cells[J].Cell Res,2013,23:162-166.
[11] WU Z,CHEN J J,REN J T,et al.Generation of piginduced pluripotent stem cells with a druginducible system[J].J Mol Cell Biol,2009,1:46-54.
[12] LIU K,JI G Z,MAO J,et al.Generation of porcineinduced pluripotent stem cells by using OCT4 and KLF4 porcine factors[J].Cell reprogram,2012,14(6):505-513.
[13] MONTSERRAT N,DE OATE L,GARRETA E,et al.Generation of feederfree pig induced pluripotent stem cells without Pou5f1[J].Cell transplantation,2012,21(5):815-825.
[14] RIDEOUT W M Ⅲ,WAKAYAMA T,WUTZ A,et al.Generation of mice from wildtype and targeted ES cells by nuclear cloning[J].Nat Genet,2000,24:109-110.
[15] KEEFER C L.Production of bioproducts through the use of transgenic animal models[J].Anim Reprod Sci,2004,82/83:5-12.
[16] DONOVAN D M,KERR D E,WALL R J.Engineering disease resistant cattle[J].Transgenic Res,2005,14(5):563-567.
[17] WALL R J,POWELL A M,PAAPE M J,et al.Genetically enhanced cows resist intramammary Staphylococcus aureus infection[J].Nat biotechnol,2005,23(4):445-451.
[18] ZHOU S,DING C,ZHAO X,et al.Successful generation of cloned mice using nuclear transfer from induced pluripotent stem cells[J].Cell Res,2010,20:850-853.
[19] KOU Z H,KANG L,YUAN Y,et al.Mice cloned from induced pluripotent stem cells(iPSCs)[J].Biol Reprod,2010,83(2):238-243.
[20] WAN Y J,ZHANG Y L,ZHOU Z R,et al.Efficiency of donor cell preparation and recipient oocyte source for production of transgenic cloned dairy goats harboring human lactoferrin[J].Theriogenology,2012,78(3):583-592.
[21] MAHERALI N,AHFELDT T,RIGAMONTI A,et al.A highefficiency system for the generation and study of human induced pluripotent stem cells[J].Cell stem cell,2008,3(3):340-345.
