3株H3N2亞型禽流感病毒的基因組特征與演化分析
2022-11-29 13:40:10崔明仙王星博黃彥銘卞希一馮夢珂董偉仁周繼勇
畜牧獸醫學報 2022年11期
崔明仙,王星博,黃彥銘,卞希一,馮夢珂,顏 焰,董偉仁,周繼勇
(浙江大學動物醫學中心,農業農村部動物病毒學重點實驗室,杭州 310058)
禽流感是由禽流感病毒(avian influenza virus, AIV)引起的一種從呼吸系統病變到嚴重全身敗血癥等多種癥狀的禽類傳染病[1]。AIV屬于正黏病毒科,流感病毒屬,根據其對雞的致病性,可分為低致病性AIV(low pathogenic avian influenza virus, LPAIV)和高致病性AIV(high pathogenic avian influenza virus, HPAIV)[2]。H3亞型AIV是LPAIV中分離率較高的一種亞型[3]。H3亞型流感病毒宿主范圍較廣,除感染禽類外,陸續發現該亞型流感病毒還能感染馬、犬、貓等哺乳動物[4]。H3亞型流感病毒易發生基因重組[5],如豬H3N2病毒在豬這個“混合器”中通過與人流感病毒發生基因重排,可形成感染人的新型豬流感病毒[6]。近三年來,趙婉宸等[7]在中國華東地區(江蘇、山東和安徽)共分離到6株鴨或鵝源的H3N2亞型AIV,但未見浙江省H3N2亞型AIV的報道。因此,對浙江地區開展家禽H3N2亞型AIV 分子流行病學檢測,密切關注其傳播和進化情況,不僅對畜牧業生產十分重要,還具有重要的公共衛生學意義。
本試驗以2021年浙江地區分離到的3株H3N2亞型AIV為研究對象,對其開展分子特征及遺傳進化分析,以期為浙江乃至華東地區H3N2亞型AIV的流行病學監測提供數據參考。
1 材料與方法
1.1 實驗材料和試劑
反轉錄試劑盒購自賽默飛世爾科技公司;Trizol試劑、Phanta Max Super-fidelity DNA Polymerase等購自南京諾唯贊生物科技有限公司;9~12日齡SPF雞胚購自寧波純派農業科技有限公司;氯仿、異丙醇、乙醇購自國藥集團化學試劑有限公司;引物由浙江尚亞生物技術有限公司合成。
1.2 樣品檢測與分離
2021年,于浙江省四地(嘉興、湖州、舟山、臺州)采集禽類共923份咽肛拭子。采用RT-PCR技術對樣品進行AIV的檢測和亞型的鑒定。采用雞胚分離法分離病毒[8]。采用雞胚終點稀釋法純化病毒[9],采用血凝試驗(HA)和PCR法對分離到的病毒進行鑒定[10]。經PCR流感病毒HA亞型檢測,只檢測到單一亞型的樣品用Trizol法抽提病毒RNA,并送晶能生物技術(上海)有限公司進行二代測序(Illumina NovaSeq 6000 測序平臺),對測序結果利用Burrows-Wheeler Aligner(版本 0.7.17)[11]和 SAMtools(版本 1.10)[12]進行序列的比對。
1.3 病毒全基因組序列分析
將測序結果用BLAST工具進行同源序列搜索,采用DNAStar軟件中MegAlign模塊進行同源性分析;對NCBI和GISAID數據庫中的相關序列進行篩選,篩選標準:1)刪除兩個數據庫中的重復序列;2)須具有完整開放閱讀框。從NCBI和GISAID數據庫中最終獲得H3N2HA基因(717條)、H3N6HA基因(21條)、H3N2NA基因(605條)和H6N2NA基因(331條)。利用軟件RAxML(版本8.2.4)[13]繪制HA和NA基因進化樹,并用iTOL v4[14]進行可視化分析。
2 結 果
2.1 樣品檢測與病毒分離鑒定
2021年,采自嘉興、湖州、舟山和臺州的小型養殖場的雞源咽肛拭子分別為386、294、100和30份,采自嘉興和臺州的鴨源咽肛拭子分別為3和110份,共計923份樣品。AIV總陽性率為7.69%(71/923),其中,采自嘉興市、湖州市、舟山市和臺州市樣品的AIV陽性率分別為4.88%(19/389)、8.16%(24/294)、28%(28/100)和0(0/140)。雞源樣品AIV陽性率為8.40%(68/810),鴨源樣品AIV陽性率為2.65%(3/113)。從嘉興市樣品中分離到1株鴨源H3N2亞型AIV(A/duck/Zhejiang/Y1/2021, ZJ/Y1),從湖州市樣品中分離到2株雞源H3N2亞型AIV(A/chicken/Zhejiang/57/2021, ZJ57和A/chicken/Zhejiang/64/2021, ZJ64)。
2.2 病毒全基因組同源分析
BLAST比對結果顯示,2株雞源ZJ57和ZJ64的HA基因與A/duck/Hunan/161/2015(H3N6)相似性高達94.42%,鴨源ZJ/Y1的HA基因與A/duck/Hubei/ZYSYF18/2015(H3N6)相似性為97.25%,它們均與H3N6相似度較高;3株分離株的NA基因與H6N2相似性均在96.24%以上;PB2、M基因與H1Nx相似性為97.24%和97.66%以上;ZJ57和ZJ64的PB1基因、ZJ57的NS基因、ZJ/Y1的PA基因均與H10N7對應基因具有較高相似性(表1)。3株H3N2亞型分離株之間的HA基因核苷酸相似性為93.4%~100.0%,NA基因之間的核苷酸相似性為94.0%~99.9%。
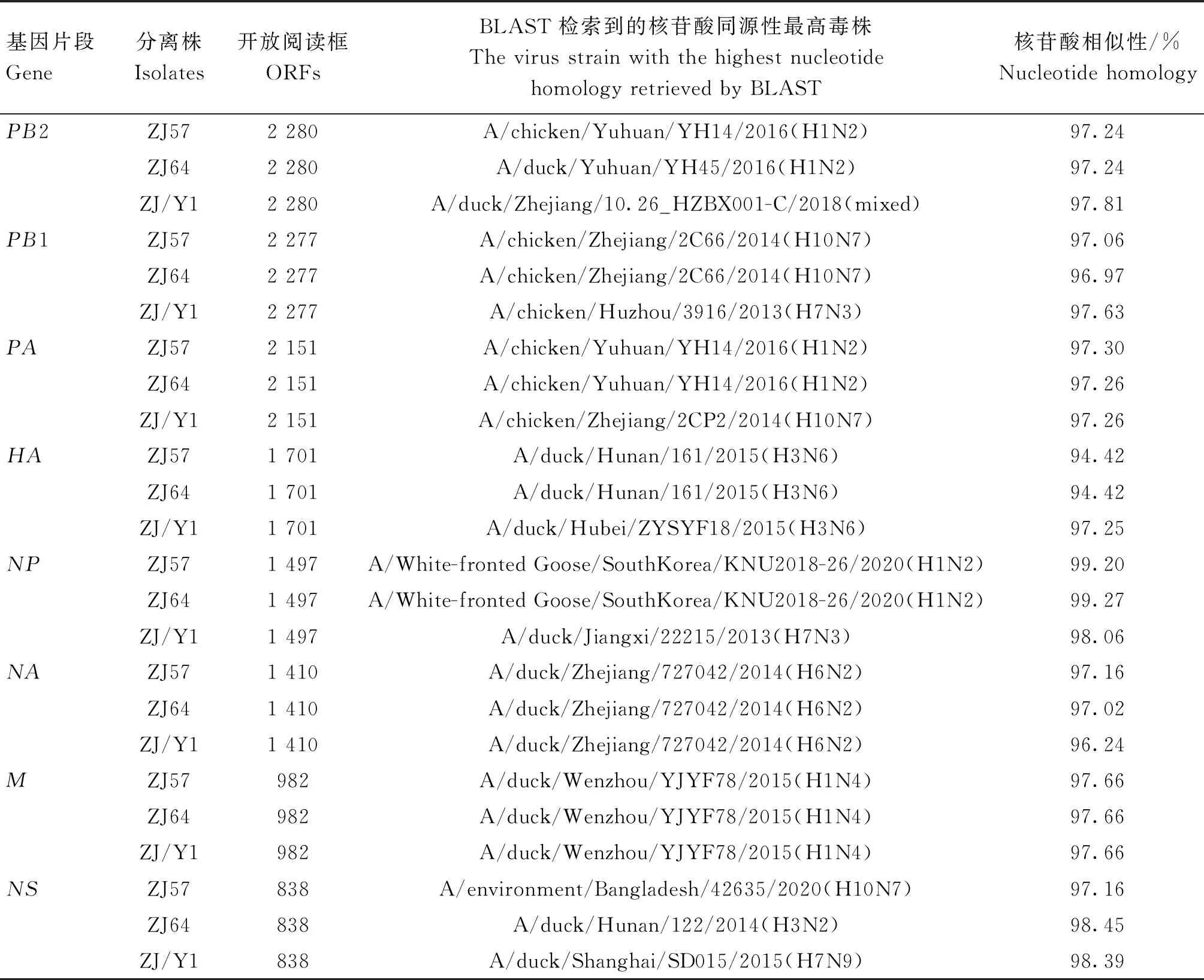
表1 3株H3N2亞型AIV分離株的BLAST分析
2.3 病毒HA和NA基因遺傳進化分析
遺傳進化分析表明:從宿主范圍來看,H3N2亞型流感病毒的宿主范圍可分為不同分支,禽源分支在遺傳進化上與豬源、人源和犬/貓源均不在同一分支;H3N2亞型AIV的主要宿主為鴨(圖1);從地區分布來看,H3N2亞型流感病毒在全國均有分布,其中,H3N2亞型AIV主要流行于華東地區;從時間分布來看,AIV主要流行于2010年以后。3株H3N2亞型分離株的HA和NA基因均屬于禽源分支,雞源ZJ57和ZJ64的HA基因與A/duck/Hunan/161/2015(H3N6)遺傳距離最近,而鴨源ZJ/Y1與A/duck/Hubei/ZYSYF9/2015(H3N6)遺傳距離最近,雞源ZJ57和ZJ64的HA基因與NA基因與A/duck/Guangxi/S31299/2015(H6N2)遺傳距離最近,而鴨源ZJ/Y1與A/duck/Guangdong/F1172/2018(H3N6)遺傳距離最近(圖1)。
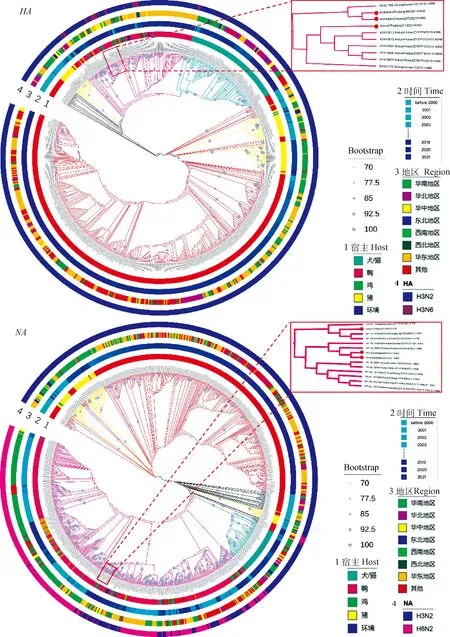
圖1 3株H3N2亞型流感病毒HA和NA基因遺傳進化樹
2.4 病毒分子特征分析
2.4.1HA和NA基因分子特征分析 3株H3N2亞型AIV的HA基因編碼區均由1 701個核苷酸組成,編碼566個氨基酸。HA1區編碼344個氨基酸,HA2區編碼221個氨基酸。分離株的HA蛋白裂解位點序列均為PEKQTR↓GLF,具有典型的低致病性禽流感病毒特征。3株分離株的HA中影響流感病毒與受體特異性結合的226Q、228G、190E和225G未發生突變,位于結合位點周圍的4個高度保守的殘基(Y98、W153、H183、Y195)也未發生突變,提示分離株跨物種傳播潛力較低。分離株HA蛋白的潛在N型糖基化位點均為6個,與浙江省禽源和人源分離株相比,存在糖基化位點增添和缺失的情況(表2)。3株H3N2亞型AIV的NA基因編碼區均由1 410個核苷酸組成,編碼469個氨基酸,頸部區無氨基酸缺失。NA蛋白共有5個預測的N型糖基化位點,分別為第61位(NITE)、69位(NNTI)、86位(NWSK)、146位(NGTI)和234位(NGTC)。

表2 3株H3N2亞型禽流感病毒HA蛋白N型糖基化位點預測
2.4.2 內部片段基因分子特征分析 3株H3N2亞型禽流感分離株PB2蛋白中與哺乳動物適應性相關的第627位關鍵位點均為Glu,符合禽流感特征,而人源A/Homosapien/China/LS340/2019(H3N2)株為Lys。PB1蛋白中與哺乳動物致病性相關的第66位氨基酸位為Ser(人源H3N2為Leu),而PB2中在跨物種傳播起關鍵作用的701位氨基酸位點、PB1蛋白中與小鼠致病性相關的第198和317位氨基酸位點、PA蛋白中能使毒力增強的第97位氨基酸位點、M1蛋白中能增強病毒對小鼠致病性的第30和215位氨基酸位點均未發生突變。
3 討 論
AIV為單股負鏈RNA病毒[15],由于其在RNA復制過程中缺乏校對機制,導致點突變頻率很高。除通過快速突變產生遺傳多樣性外,同源重組是另外一種重要的進化機制,它驅動病毒基因變異的形成,使AIV能夠克服選擇壓力而適應新的環境和宿主[16]。本研究結果顯示,已報道的H3N2亞型AIV主要流行于2010年后,集中分布在華東地區,鴨是其主要宿主。分離株各基因片段來源復雜,說明它們可能是經過長時間進化而發生自然重排后形成的重組病毒:分離株的HA基因與湖南和湖北兩地H3N6亞型毒株距離較近,ZJ57和ZJ64的NA基因與廣西的H6N2毒株遺傳關系較近,ZJ/Y1與廣東的H3N6毒株遺傳距離最近,進一步說明3株H3N2亞型AIVs片段來源較復雜,可能發生了基因重組。
流感病毒受體結合特異性會影響其跨物種傳播。人H3流感病毒HA蛋白226L和228S特征使其優先與人細胞表面的α2-6半乳糖苷唾液酸(SAα2-6Gal)受體結合,而禽源優先與α2-3半乳糖苷唾液酸(SAα2-3Gal)受體結合[17],PB2中的627E和701K氨基酸位點也在流感病毒跨種間傳播中起關鍵性作用[18]。本研究中3株H3N2亞型禽流感病毒分離株HA蛋白的226Q和228G特征表明其更傾向于結合禽源的SAα2-3Gal受體,提示分離株的跨物種傳播潛力較低。PB2蛋白序列中與哺乳動物適應性相關的第627位關鍵位點均為Glu,符合禽流感病毒特征。PB1中的66S、198K和701M氨基酸位點,M1中的30D和215A能增強流感病毒致病性[19-22]。但PB1蛋白中第66位氨基酸位點均由Leu突變為Ser,但是否會增強對哺乳動物的致病性還需進一步驗證。
禽流感病毒HA和NA蛋白都修飾有N-連接寡糖(N型糖基化位點)[23],流感病毒HA蛋白頭部的糖基化位點的結構和數量不相同,是導致病毒抗原多樣性的一個重要原因,它能夠掩蓋或者修飾抗原位點,減少先天免疫系統給病毒的生存壓力[24]。本研究發現浙江人源A/Homosapien/China/LS340/2019(H3N2)株HA蛋白和NA蛋白的糖基化位點較豐富,而禽源A/duck/Zhejiang/D13/2013(H3N2)[25]糖基化位點相對較少,3株分離株的糖基化位點均存在不同情況的缺失和增加,其是否會對病毒抗原多樣性造成影響還需進一步探究。
4 結 論
2021年,浙江地區禽流感陽性率為7.70%,分離到3株H3N2亞型AIVs,其基因片段來源復雜,推測可能由不同亞型毒株經過長時間進化而發生自然重排后形成。因未發生與哺乳動物適應性位點相關的突變,表明其跨種傳播潛力較低,但PB1中66位氨基酸突變為S,其是否會增加哺乳動物的致病性仍需進一步探究。
