C型產(chǎn)氣莢膜梭菌實(shí)驗(yàn)感染仔豬腸道circRNA的表達(dá)特征
2022-11-29 13:40:06黃曉宇楊巧麗閆尊強(qiáng)王鵬飛石海仁滾雙寶
畜牧獸醫(yī)學(xué)報(bào) 2022年11期
關(guān)鍵詞:差異
黃曉宇,楊巧麗,閆尊強(qiáng),王鵬飛,石海仁,滾雙寶,3*
(1.甘肅農(nóng)業(yè)大學(xué)動(dòng)物科學(xué)技術(shù)學(xué)院,蘭州 730070; 2.西藏自治區(qū)農(nóng)牧科學(xué)院,拉薩 850000;3.甘肅省現(xiàn)代養(yǎng)豬工程技術(shù)研究中心,蘭州 730070)
產(chǎn)氣莢膜梭菌(Clostridiumperfringens,C.perfringens)是一種革蘭陰性厭氧芽孢桿菌,廣泛分布于自然環(huán)境、人和動(dòng)物胃腸道微生物群[1-2]。產(chǎn)氣莢膜梭菌能夠引起一系列人類和動(dòng)物腸道疾病(CAED),例如氣體壞疽、食物中毒、壞死性小腸結(jié)腸炎、炎癥性疾病、仔豬腹瀉、綿羊腸毒血癥等[3-4],逐漸成為引起人類和動(dòng)物組織中毒性疾病和腸道疾病的主要原因。按其所產(chǎn)4種主要致命毒素(α、β、ε和ι)類型不同,產(chǎn)氣莢膜梭菌被分為5種類型(A~E)[5]。
C型產(chǎn)氣莢膜梭菌(Clostridiumperfringenstype C,C.perfringenstype C, Cp)是新生仔豬腹瀉的重要感染性病原體之一[6],已成為仔豬高發(fā)病率和高死亡率的重要因素,給全球養(yǎng)豬業(yè)造成了巨大的經(jīng)濟(jì)損失[7]。C型產(chǎn)氣莢膜梭菌被認(rèn)為是引起哺乳動(dòng)物高致命性、暴發(fā)性壞死性腸炎、小腸結(jié)腸炎和全身性疾病的主要原因[8], 同時(shí)也是幼畜死亡的一個(gè)重要原因[9]。此外,人類因食用受污染的動(dòng)物產(chǎn)品或直接接觸受感染的動(dòng)物亦會(huì)感染該菌[9]。C型產(chǎn)氣莢膜梭菌產(chǎn)生的α和β毒素進(jìn)入腸道,吸附在腸道黏膜上,經(jīng)腸道屏障系統(tǒng)被吸收到機(jī)體血液循環(huán)中,進(jìn)而影響其他器官,如大腦[4]。目前,一些動(dòng)物模型已被用來(lái)研究產(chǎn)氣莢膜梭菌相關(guān)的腸道疾病[10-13], 如雞、兔和鼠。但是目前關(guān)于仔豬如何應(yīng)答C型產(chǎn)氣莢膜梭菌感染導(dǎo)致的腹瀉疾病尚不清楚。
circRNA是一種新型的非編碼RNA,由不含50-30極性和poly(A)尾孔的共價(jià)閉環(huán)結(jié)構(gòu),經(jīng)數(shù)千個(gè)基因的前體mRNA (pre-mRNA)反向剪接產(chǎn)生,在細(xì)胞中更穩(wěn)定[14-15]。諸多研究表明,circRNA不僅可以通過(guò)激活或抑制基因的轉(zhuǎn)錄參與調(diào)控其功能,還能通過(guò)靶向miRNA、轉(zhuǎn)錄和干擾pre-mRNA剪接在基因表達(dá)中發(fā)揮潛在的調(diào)節(jié)作用[16-17]。但是目前有關(guān)circRNA如何參與調(diào)控仔豬應(yīng)答C型產(chǎn)氣莢膜梭菌感染導(dǎo)致的腹瀉疾病尚不清楚。在本研究中,首先構(gòu)建一個(gè)C型產(chǎn)氣莢膜梭菌感染仔豬致腹瀉的動(dòng)物模型,并通過(guò)RNA測(cè)序全面描述C型產(chǎn)氣莢膜梭菌感染仔豬的回腸組織中circRNAs特異性表達(dá)譜,篩選并鑒定出差異表達(dá)的circRNAs,構(gòu)建與C型產(chǎn)氣莢膜梭菌性疾病密切相關(guān)的ceRNA互作網(wǎng)絡(luò),本研究不僅能為進(jìn)一步研究circRNA調(diào)控仔豬抵抗C型產(chǎn)氣莢膜菌性腹瀉提供基礎(chǔ),還可為今后豬抗腹瀉新品系的培育提供參考。
1 材料與方法
1.1 菌株培養(yǎng)
將C型產(chǎn)氣莢膜梭菌菌株CVCC 2032(購(gòu)自中國(guó)農(nóng)業(yè)微生物菌種保藏管理中心,北京)置于牛肉湯培養(yǎng)基中,37 ℃搖晃培養(yǎng)16 h (青島海博生物)。采用平板菌落計(jì)數(shù)法測(cè)定C型產(chǎn)氣莢膜梭菌的菌落形成單位(CFU)。
1.2 動(dòng)物試驗(yàn)
選擇6頭7日齡長(zhǎng)大二元仔豬,經(jīng)酶聯(lián)免疫吸附ELISA試劑盒(南京建成生物工程研究所,中國(guó))檢測(cè)血清中大腸桿菌、沙門菌和產(chǎn)氣莢膜梭菌均為陰性后作為試驗(yàn)仔豬,在適當(dāng)?shù)臍夂蚩刂坪屯耆綦x條件下單獨(dú)飼養(yǎng),自由采食。隨機(jī)選取3頭仔豬經(jīng)口灌服1 mL 1×109CFU·mL-1C型產(chǎn)氣莢膜梭菌培養(yǎng)液[18],連續(xù)5 d,作為處理組(TI組),剩余3頭仔豬灌服無(wú)菌培養(yǎng)液,作為對(duì)照組(CI組)。
接種后,每天監(jiān)測(cè)每只仔豬的健康狀況,包括行為、食欲、精神和毛發(fā)狀況。評(píng)估并記錄仔豬每天糞便黏稠度,按照腹瀉評(píng)分標(biāo)準(zhǔn)進(jìn)行評(píng)定:0 =正常、固體糞便;1=輕微腹瀉,糞便柔軟疏松;2=中度腹瀉,質(zhì)軟不成型糞便;3=嚴(yán)重腹瀉、水樣糞便[19]。試驗(yàn)結(jié)束后,采用巴比妥酸鹽麻醉法人道屠宰6頭仔豬,無(wú)菌采集TI組和CI組仔豬的回腸組織樣品,用無(wú)菌PBS緩沖液(pH 7.4)沖洗干凈,液氮速凍并保存于-80 ℃超低溫冰箱,以備RNA提取。
1.3 總RNA提取和 circRNA測(cè)序
提取TI組和CI組樣品中總RNA,經(jīng)1%瓊脂糖凝膠檢測(cè)后,利用Qubit?RNA Assay Kit測(cè)定RNA濃度。使用NanoPhotometer分光光度計(jì)(IMPLEN, CA, USA)和RNA Nano 6000檢測(cè)試劑盒(Agilent Technologies, CA, USA)分別評(píng)估總RNA的純度和完整性。
取TI組和CI組樣品RNA 5 μg,按照說(shuō)明書去除樣品核糖體RNA (Epicentre RibozeroTMrRNA 去除試劑盒,Epicentre, 美國(guó))和線性RNA(RNase R,Epicentre, USA)。利用Next?UltraTMDirectional RNA Library Prep Kit制備circRNA測(cè)序文庫(kù)(NEB, 美國(guó)),通過(guò)Agilent Bioanalyzer 2100系統(tǒng)進(jìn)行質(zhì)量評(píng)估后,在Illumina Hiseq 4000平臺(tái)(Illumina, San Diego, CA, USA)上進(jìn)行測(cè)序,獲得150 bp配對(duì)端(PE150)序列。
1.4 數(shù)據(jù)比對(duì)
去除包含接頭序列、ploy-N和低質(zhì)量(>50% of bases with Phred scores<5)的原始數(shù)據(jù)后,獲取clean數(shù)據(jù),通過(guò)計(jì)算clean數(shù)據(jù)的Q20、Q30和GC含量,使用Bowtie2[20]將clean reads與豬參考基因組進(jìn)行比對(duì)。
1.5 circRNA的鑒定和定量
使用clean reads中的find_circ[21]和CIRI2[22]檢測(cè)并識(shí)別circRNAs。首先使用TPM(Transcripts read count Per Kilobase Million)對(duì)潛在 circRNAs的原始計(jì)數(shù)進(jìn)行標(biāo)準(zhǔn)化,以評(píng)估表達(dá)水平[23]。
1.6 circRNAs差異表達(dá)分析
基于負(fù)二項(xiàng)分布,使用DESeq2 R包[24]對(duì)circRNAs進(jìn)行差異表達(dá)分析。采用Benjamini和Hochberg方法控制錯(cuò)誤發(fā)生率,以P值<0.05的circRNAs視為差異表達(dá)。
1.7 GO和KEGG富集分析
利用DAVID (http://david.abcc.ncifcrf.gov)分析和預(yù)測(cè)差異表達(dá)circRNA的線性轉(zhuǎn)錄本;利用GOseq R包[25]和 KOBAS 2軟件[26]對(duì)差異表達(dá)circRNA宿主基因進(jìn)行Gene Ontology (GO)功能和Kyoto Encyclopedia of Genes and Genomes (KEGG)信號(hào)通路富集分析。通過(guò)KOBAS軟件統(tǒng)計(jì)circRNA宿主基因的富集度[27],校正P值<0.05為顯著富集。
1.8 circRNA-miRNA-gene互作網(wǎng)絡(luò)的預(yù)測(cè)
通過(guò)miRanda軟件預(yù)測(cè)差異表達(dá)circRNA外顯子中的microRNA靶點(diǎn),構(gòu)建circRNA/miRNA相互作用關(guān)系,用Cytoscape生成circRNA-miRNA-mRNA互作網(wǎng)絡(luò)圖[28]。
1.9 circRNA-miRNA-gene基因網(wǎng)絡(luò)與CAED分析
為研究circRNA相關(guān)的ceRNA網(wǎng)絡(luò)與CAED之間潛在調(diào)控關(guān)系,首先篩選TI組和CI組仔豬回腸組織中顯著差異表達(dá)circRNAs、miRNAs及其靶mRNAs的數(shù)據(jù)集(校正P<0.05),同時(shí)確保circRNAs、miRNAs及其靶miRNAs的表達(dá)水平在一定的數(shù)量級(jí);其次,circRNA相關(guān)的ceRNA網(wǎng)絡(luò)與CAED相關(guān) (在Web of Science中搜索miRNA,在Genecards數(shù)據(jù)庫(kù)中搜索mRNA);最后,進(jìn)一步篩選參與炎癥、免疫或感染等相關(guān)信號(hào)通路的差異表達(dá)circRNAs。根據(jù)上述4個(gè)步驟篩選與CAED相關(guān)的ceRNA網(wǎng)絡(luò)。
1.10 qPCR驗(yàn)證及數(shù)據(jù)分析
采用熒光定量qPCR檢測(cè)ceRNA網(wǎng)絡(luò)中6個(gè)circRNAs、3個(gè)miRNAs和4個(gè)mRNAs的表達(dá)水平,分別以GAPDH基因和U6基因作為內(nèi)參基因,引物信息見(jiàn)表1。采用9.5 μL 2 × SYBR Green Real-time PCR Master Mix (TaKaRa,中國(guó)大連),正、反引物各1 μL,1 μL cDNA和7.5 μL游離RNase ddH2O的20 μL反應(yīng)體系,在Roche熒光定量PCR儀進(jìn)行檢測(cè),溶解曲線用于評(píng)價(jià)PCR產(chǎn)物的特異性。
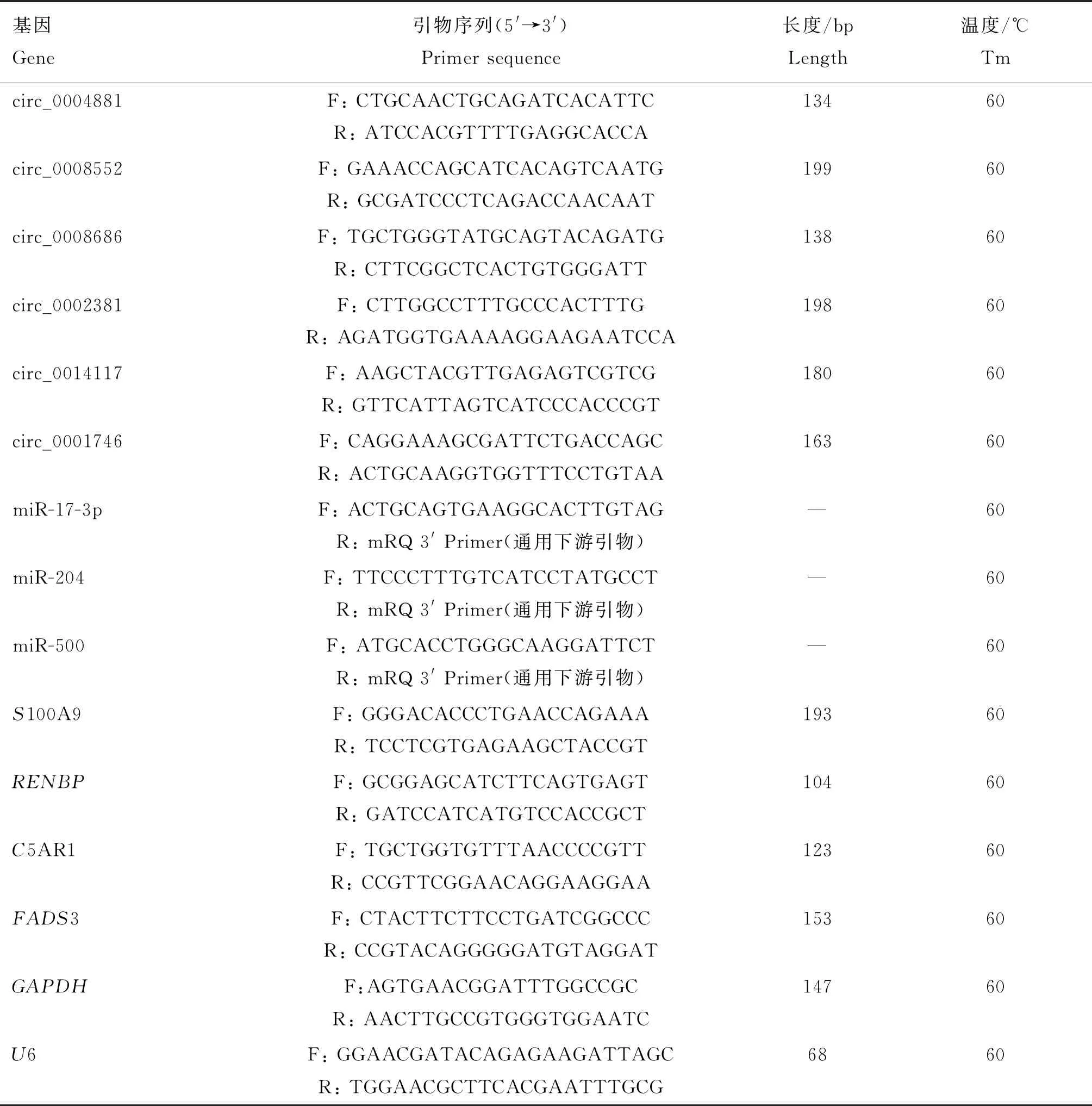
表1 circRNA、miRNA 和 mRNA熒光定量qPCR引物信息
1.11 數(shù)據(jù)統(tǒng)計(jì)分析
利用SPSS 21.0軟件進(jìn)行試驗(yàn)數(shù)據(jù)的統(tǒng)計(jì)學(xué)分析,采用獨(dú)立樣本T-test 分析和單因素方差分析(one-way ANOVA)檢測(cè)TI組和CI組仔豬糞便腹瀉評(píng)分;定量結(jié)果采用2-ΔΔCt法計(jì)算相對(duì)表達(dá)量,所得數(shù)值均用“平均值±標(biāo)準(zhǔn)誤(X±SE)”表示,每個(gè)樣品設(shè)置3個(gè)生物學(xué)重復(fù)。
2 結(jié) 果
2.1 仔豬腹瀉評(píng)分及糞便中C型產(chǎn)氣莢膜梭菌數(shù)量
Cp感染導(dǎo)致仔豬出現(xiàn)了不同程度的腹瀉,對(duì)TI組和CI組仔豬的平均腹瀉評(píng)分和總腹瀉評(píng)分進(jìn)行統(tǒng)計(jì)及差異顯著性分析。結(jié)果發(fā)現(xiàn),TI組仔豬的平均腹瀉評(píng)分和總腹瀉評(píng)分分別為1.51±0.29和37.33±2.51,均極顯著高于CI組仔豬的平均腹瀉評(píng)分0.49±0.03和總腹瀉評(píng)分12.00±1.00(P<0.01,表2)。

表2 TI組和CI組仔豬腹瀉評(píng)分統(tǒng)計(jì)分析
2.2 Cp感染仔豬回腸circRNA的鑒定
經(jīng)測(cè)序鑒定,C型產(chǎn)氣莢膜梭菌感染后,TI組和CI組仔豬回腸組織中共檢測(cè)到3 162個(gè)circRNAs(圖1A),這些circRNAs的長(zhǎng)度分布范圍為1~40 kb,大部分circRNA長(zhǎng)度小于3 kb(圖1B),其中最大長(zhǎng)度為2 135 nt,最小長(zhǎng)度為22 nt,平均長(zhǎng)度為298 nt(圖1C)。大多數(shù)circRNA含有1~3個(gè)外顯子、內(nèi)含子或基因間區(qū)域(圖1D)。在染色體分布方面,這些鑒定到的circRNAs主要來(lái)源于編碼蛋白質(zhì)的基因外顯子區(qū)域(占81.39%),部分來(lái)自基因間區(qū)域,少數(shù)來(lái)自內(nèi)含子區(qū)域(圖1E)。
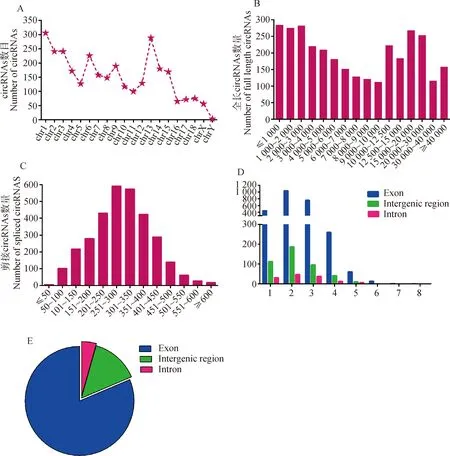
A. 仔豬染色體circRNAs的鑒定; B. 不同全長(zhǎng)的circRNAs數(shù)量;C. 不同剪接長(zhǎng)度的circRNAs數(shù)量;D. circRNAs外顯子區(qū)、基因間區(qū)和內(nèi)含子區(qū)3種亞型特征;E. 仔豬回腸circRNAs分類
將Cp感染仔豬回腸組織 circRNA原始數(shù)據(jù)提交至NCBI Sequence Read Archive (SRA)數(shù)據(jù)庫(kù),獲得登錄號(hào)PRJNA705036。
2.3 差異表達(dá)circRNA分析
對(duì)Cp感染的TI組和CI組鑒定到的3 162個(gè)circRNAs進(jìn)行分析,發(fā)現(xiàn)2 854個(gè)circRNAs在兩組間共同表達(dá),203個(gè)circRNAs和105個(gè)circRNAs僅在TI組和CI組中特異性表達(dá)(圖2A)。聚類結(jié)果顯示,TI組和CI組具有相同表達(dá)模式的circRNA聚類在一起(圖2B)。
以校正后P值 < 0.05為標(biāo)準(zhǔn)對(duì)鑒定到的circRNA進(jìn)行差異表達(dá)分析, 從TI組和CI組共鑒定到694個(gè)circRNAs顯著差異表達(dá), 其中上調(diào)表達(dá)404個(gè)circRNAs,下調(diào)表達(dá) 290個(gè)circRNAs(圖2C)。還對(duì)cirRNAs進(jìn)行了環(huán)狀基因組分析(圖3),TI組和CI組中差異表達(dá)cirRNAs主要分布在1、2、3、6、8、9、13、14、15和X染色體上,每條染色體上分布的差異表達(dá)cirRNAs數(shù)量和差異倍數(shù)差別明顯(圖3)。
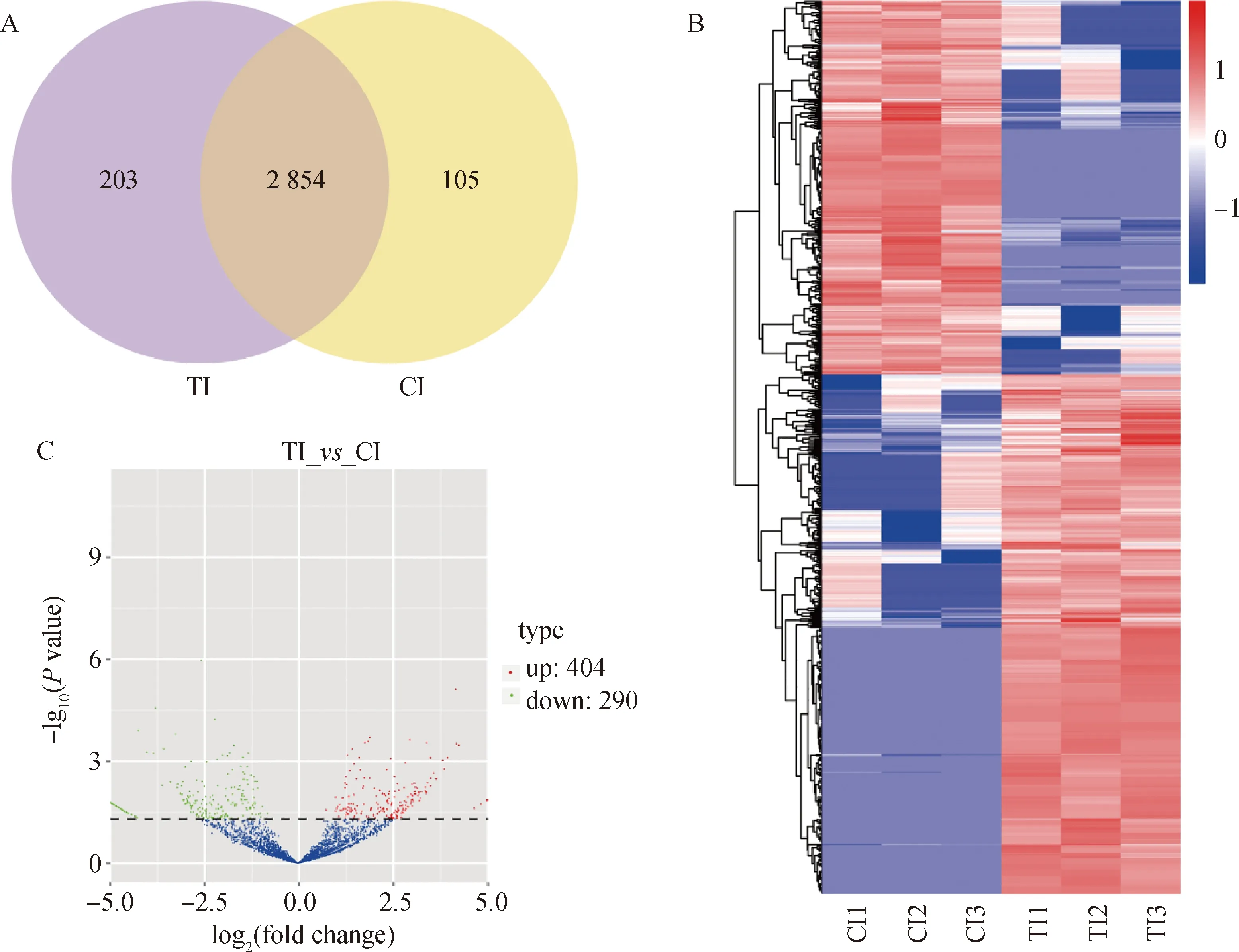
A. 維恩圖;B. 聚集熱圖,行表示差異表達(dá)circRNAs,列表示不同樣本;C. 火山圖,紅點(diǎn)和藍(lán)點(diǎn)分別代表上調(diào)和下調(diào)的circRNAs
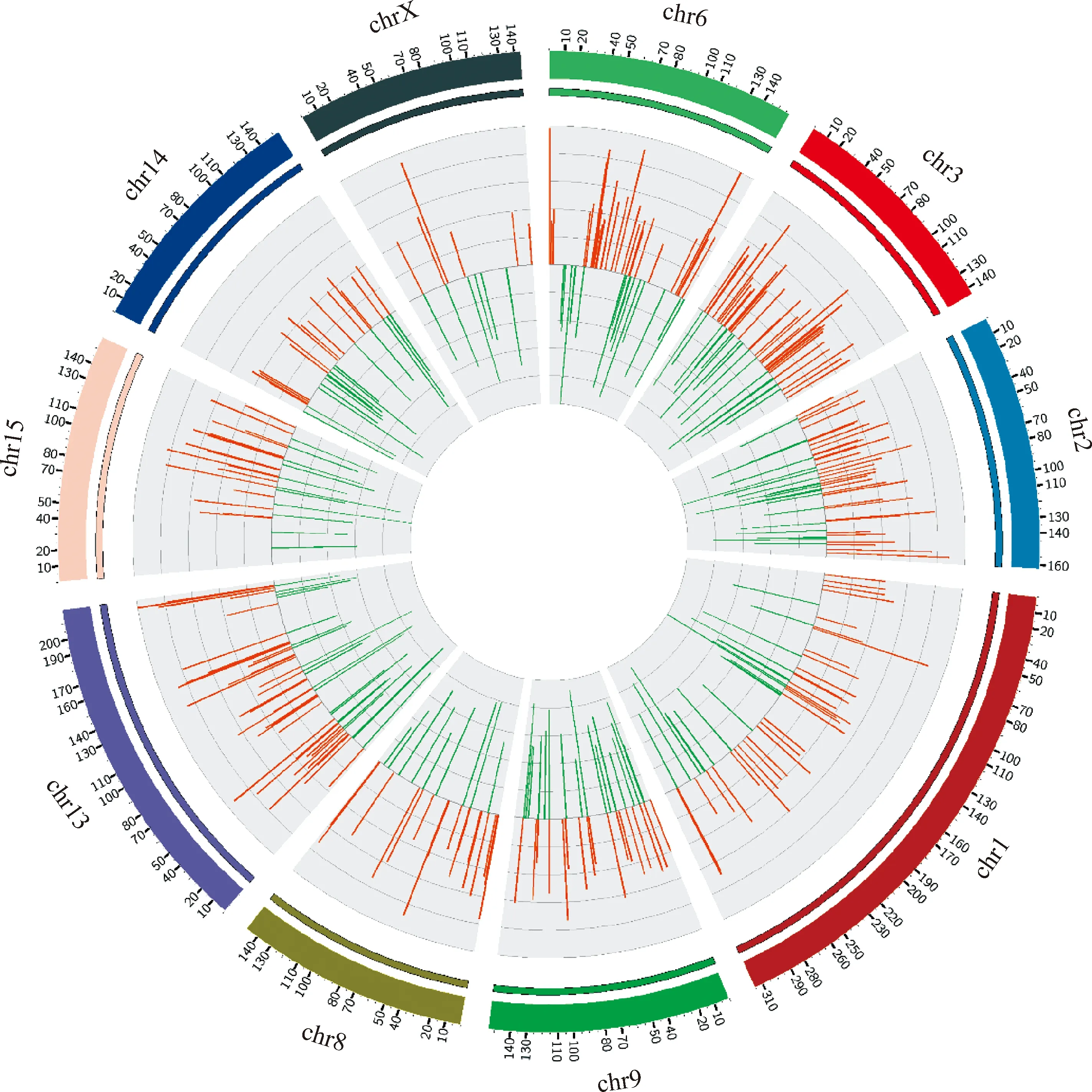
紅色代表上調(diào)circRNAs,綠色代表下調(diào)circRNAs
2.4 功能富集分析
circRNAs的功能與circRNA的親本基因的功能相關(guān)。對(duì)TI組與CI組間差異表達(dá)circRNAs的親本基因進(jìn)行GO功能富集分析,發(fā)現(xiàn)差異表達(dá)circRNAs主要富集在135個(gè)GO功能(校正P<0.05),其中包括52個(gè)極顯著富集的GO功能(校正P<0.01),包括26個(gè)生物過(guò)程、25個(gè)細(xì)胞組分和1個(gè)分子功能,如細(xì)胞器組織、細(xì)胞代謝過(guò)程、細(xì)胞大分子代謝過(guò)程等(圖4A)。KEGG信號(hào)通路富集分析鑒定出197條顯著富集的信號(hào)通路,top20富集的KEGG通路包括細(xì)胞周期、TGF-beta信號(hào)通路、賴氨酸降解、Wnt信號(hào)通路、T細(xì)胞受體信號(hào)通路、MAPK信號(hào)通路等(圖4B),表明差異表達(dá)circRNAs通過(guò)其親本基因參與了細(xì)胞、免疫和信號(hào)轉(zhuǎn)導(dǎo)過(guò)程,揭示了差異表達(dá)circRNA親本基因的潛在功能。
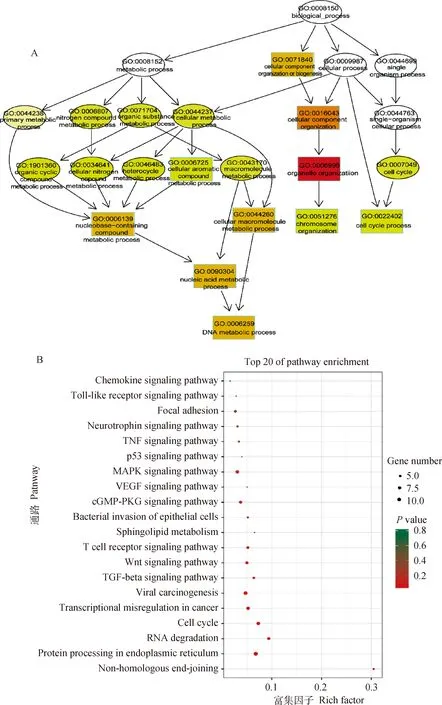
圖4 TI組和CI組之間差異表達(dá)circRNA 親本基因GO功能(A)和KEGG信號(hào)通路(B)富集分析
2.5 circRNA-miRNA-mRNA相互作用關(guān)系預(yù)測(cè)
通過(guò)circRNA轉(zhuǎn)錄組測(cè)序從Cp感染的仔豬回腸組織中鑒定出694個(gè)顯著差異表達(dá)circRNAs,結(jié)合課題組前期從Cp感染組和對(duì)照組仔豬回腸組織中鑒定出53個(gè)顯著表達(dá)miRNAs[29]和3 669個(gè)顯著表達(dá)mRNAs[30],構(gòu)建了circRNA-miRNA-mRNA交互網(wǎng)絡(luò)(包括349個(gè)circRNA、36個(gè)miRNAs和233個(gè)mRNAs)(圖5),如circ0006008- ssc-let-7i- TARBP2,circ0002897- miR-24-3p- ITM2C,circ0003611- miR-365-5p- NFKBID, circ0009977- miR-7134-3p- IL17RC等。這些ceRNA網(wǎng)絡(luò)基因可能在仔豬應(yīng)答Cp感染過(guò)程中發(fā)揮重要作用。
2.6 circRNA-miRNA-mRNA網(wǎng)絡(luò)與CAED關(guān)系分析
為進(jìn)一步分析circRNA相關(guān)的ceRNA網(wǎng)絡(luò)與CAED之間可能存在的關(guān)系,篩選出由8 circRNAs-5 miRNAs-12 mRNAs組成的關(guān)系網(wǎng)絡(luò),由圖6可知,novel-circ0004881能夠通過(guò)競(jìng)爭(zhēng)性結(jié)合miR-17-3p調(diào)控S100A9基因和ERBB2基因,novel-circ0014949能夠競(jìng)爭(zhēng)性結(jié)合miR-204調(diào)控FAM167基因的表達(dá),novel-circ0008552能夠競(jìng)爭(zhēng)性結(jié)合miR-500調(diào)控RENBP、C5AR1和FADS3基因的表達(dá)。
2.7 qPCR驗(yàn)證
對(duì)ceRNA網(wǎng)絡(luò)中的6個(gè)circRNAs,3個(gè)miRNAs和4個(gè)mRNAs的表達(dá)量進(jìn)行qPCR驗(yàn)證,檢測(cè)結(jié)果表明:circ0004881、circ0008552和circ0014117表達(dá)水平極顯著上調(diào)(P<0.01),circ0008686和circ0002381表達(dá)水平極顯著下調(diào)(P<0.01),miR-500、miR-17-3p、miR-204及S100A9、RENBP基因表達(dá)水平均顯著上調(diào)(P<0.05),但circ0001746、C5AR1和FADS3基因表達(dá)水平顯著下調(diào)(P<0.05),這些基因的表達(dá)水平與測(cè)序結(jié)果均一致(圖7)。
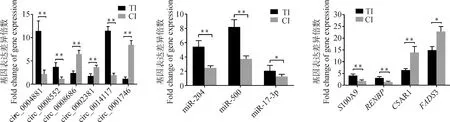
數(shù)據(jù)以“平均值±SEM”表示;*. P<0.05; **. P<0.01
3 討 論
近十年,C型產(chǎn)氣莢膜梭菌被認(rèn)為是引起仔豬急性高度傳染性腹瀉、腸道炎癥性疾病及壞死性腸炎的重要致病菌之一,嚴(yán)重威脅著世界養(yǎng)豬業(yè)的健康發(fā)展[3]。目前有關(guān)C型產(chǎn)氣莢膜梭菌感染引起仔豬腹瀉的研究主要集中在發(fā)病機(jī)制和疾病診斷、病理檢測(cè)等方面,由于人們對(duì)豬抵抗C型產(chǎn)氣莢膜梭菌感染性腹瀉疾病的調(diào)控機(jī)制研究不足,使得該類疾病目前尚無(wú)有效的治療方法。circRNAs是一種新發(fā)現(xiàn)的特殊非編碼RNA,廣泛存在于各類細(xì)胞中,具有充當(dāng)競(jìng)爭(zhēng)性內(nèi)源RNA(ceRNA),競(jìng)爭(zhēng)性結(jié)合miRNA,調(diào)控其親本基因的表達(dá)及轉(zhuǎn)錄等多種生物學(xué)功能[31]。circRNA相關(guān)的ceRNA網(wǎng)絡(luò)可能在許多免疫和炎性疾病過(guò)程中發(fā)揮關(guān)鍵作用[32-33]。研究報(bào)道腸道組織has-circ-001569的表達(dá)與miRNA-145水平呈負(fù)相關(guān),與miRNA-145靶基因E2F5、BAG4和FMNL2水平呈正相關(guān),has-circ-001569通過(guò)競(jìng)爭(zhēng)性結(jié)合miRNA-145,調(diào)控其靶基因E2F5、BAG4和FMNL2基因表達(dá),促進(jìn)細(xì)胞增殖和侵襲[34]。然而,circRNA如何調(diào)控仔豬應(yīng)答C型產(chǎn)氣莢膜梭菌感染引起的腹瀉疾病尚不清楚。
本研究報(bào)道了C型產(chǎn)氣莢膜梭菌感染的仔豬回腸circRNA的表達(dá)模式,經(jīng)分析,在TI組和CI組中共鑒定到3 162個(gè)circRNAs,其中694個(gè)circRNAs顯著差異表達(dá)(P<0.05),包括404個(gè)circRNAs上調(diào)表達(dá),290個(gè)circRNAs下調(diào)表達(dá)。為了進(jìn)一步了解差異表達(dá) circRNA的功能,本研究進(jìn)行了差異表達(dá)circRNA的功能富集分析。值得注意的是,差異表達(dá)的circRNAs主要富集在12個(gè)KEGG信號(hào)通路,如TGF-beta信號(hào)通路、T細(xì)胞受體和MAPK等免疫相關(guān)信號(hào)通路,這些通路被報(bào)道是與C型產(chǎn)氣莢膜梭菌感染性疾病密切相關(guān)的信號(hào)通路[29],該結(jié)果說(shuō)明,circRNA可通過(guò)免疫相關(guān)信號(hào)通路調(diào)控仔豬應(yīng)答C型產(chǎn)氣莢膜梭菌感染引起的腹瀉疾病。
本課題組前期報(bào)道了C型產(chǎn)氣莢膜梭菌感染仔豬回腸miRNA和mRNA的動(dòng)態(tài)變化[29-30],結(jié)合該數(shù)據(jù),在分析circRNA、miRNA和mRNA表達(dá)相關(guān)性的基礎(chǔ)上,本研究進(jìn)一步構(gòu)建了C型產(chǎn)氣莢膜感染仔豬回腸組織差異表達(dá)circRNA相關(guān)的ceRNA調(diào)控網(wǎng)絡(luò),共形成了836條circRNA、miRNA和mRNA相關(guān)關(guān)系(圖5)。在此ceRNA調(diào)控網(wǎng)絡(luò)中,發(fā)現(xiàn)一些circRNA可以與miR-15b、miR-21和miR-17-3p等競(jìng)爭(zhēng)性結(jié)合,miR-15b、let-7i、S100A9、IL7R、IL17RC、CD101和TARBP2等免疫相關(guān)分子均被報(bào)道與宿主免疫系統(tǒng)疾病密切相關(guān)。有研究報(bào)道,miR-15b、miR-17-3p、miR-29a和miR-21被認(rèn)為與結(jié)直腸癌(CRC)和炎癥性腸炎(IBD)等疾病的發(fā)生發(fā)展有關(guān),可作為疾病診斷及預(yù)后的生物標(biāo)志物[35-36]。miR-15b與宿主空腸和回腸中某些細(xì)菌的拷貝數(shù)呈正相關(guān),可作為免疫細(xì)胞發(fā)育的調(diào)節(jié)劑,對(duì)宿主腸道的發(fā)育具有潛在的調(diào)控功能[37]。因此推測(cè),circRNA可能通過(guò)競(jìng)爭(zhēng)性結(jié)合miRNA調(diào)控靶mRNA,在仔豬應(yīng)答C型產(chǎn)氣莢膜梭菌感染性腹瀉過(guò)程中發(fā)揮免疫調(diào)節(jié)作用。
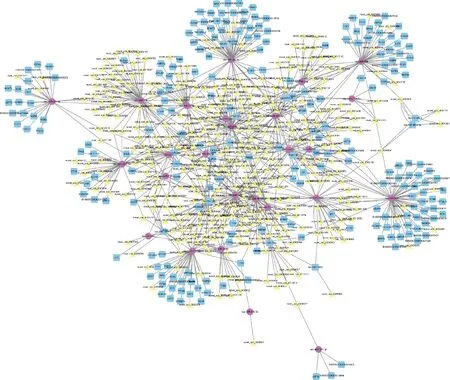
藍(lán)色正方形代表mRNA,紅色圓圈代表miRNA,黃色三角形代表circRNA
本研究篩選出了與CEAD相關(guān)的ceRNA調(diào)控網(wǎng)絡(luò),包括8個(gè)circRNAs,5個(gè)miRNAs和12個(gè)mRNAs,如circ-0004881-miR-17-3p-S100A9、circ-0002381-miR-338-FMVK、circ-0014949-miR-204-FAM167等,其中circ-0008552-miR-500與RENBP、C5AR1和FADS3均具有潛在的靶向關(guān)系(圖6A)。qPCR檢測(cè)結(jié)果表明,Cp感染導(dǎo)致仔豬回腸miR-17-3p的表達(dá)顯著上調(diào),其上游circ-0004881和下游靶基因S100A9的表達(dá)均顯著上調(diào)。miR-17-3p被circ-0004881競(jìng)爭(zhēng)性吸附,抑制3種線粒體抗氧化酶:錳超氧化物歧化酶、谷胱甘肽過(guò)氧化物酶2和硫氧還蛋白還原酶2,協(xié)同有效地去除細(xì)胞內(nèi)的活性氧,提高腫瘤的治療效率[38]。鈣結(jié)合蛋白基因S100A9具有調(diào)控細(xì)胞增殖、誘導(dǎo)細(xì)胞凋亡、參與炎性應(yīng)答、影響炎癥細(xì)胞的遷移、抵抗病原菌多種生物功能[39-40]。LPS通過(guò)抑制細(xì)胞活力、促進(jìn)細(xì)胞凋亡和促炎癥因子的產(chǎn)生,誘導(dǎo)細(xì)胞發(fā)生炎癥損傷。Zhao等[41]利用脂多糖(LPS)感染小鼠,發(fā)現(xiàn)LPS處理的小鼠肺組織中S100A9基因的表達(dá)上調(diào),說(shuō)明S100A9基因的表達(dá)水平與LPS誘導(dǎo)的小鼠肺組織的炎癥反應(yīng)密切相關(guān)。S100A9能顯著改善小鼠結(jié)腸組織的炎癥反應(yīng),減少免疫細(xì)胞(巨噬細(xì)胞、中性粒細(xì)胞)的浸潤(rùn)和促炎性細(xì)胞因子(TNF-α、IL-1β、IFN-γ、IL-6等)的產(chǎn)生[42]。本研究發(fā)現(xiàn),Cp感染導(dǎo)致仔豬回腸S100A9基因顯著上調(diào)表達(dá),說(shuō)明circ-0004881可能通過(guò)競(jìng)爭(zhēng)性結(jié)合miR-17-3p,影響S100A9基因的表達(dá),參與仔豬抵抗Cp感染過(guò)程。
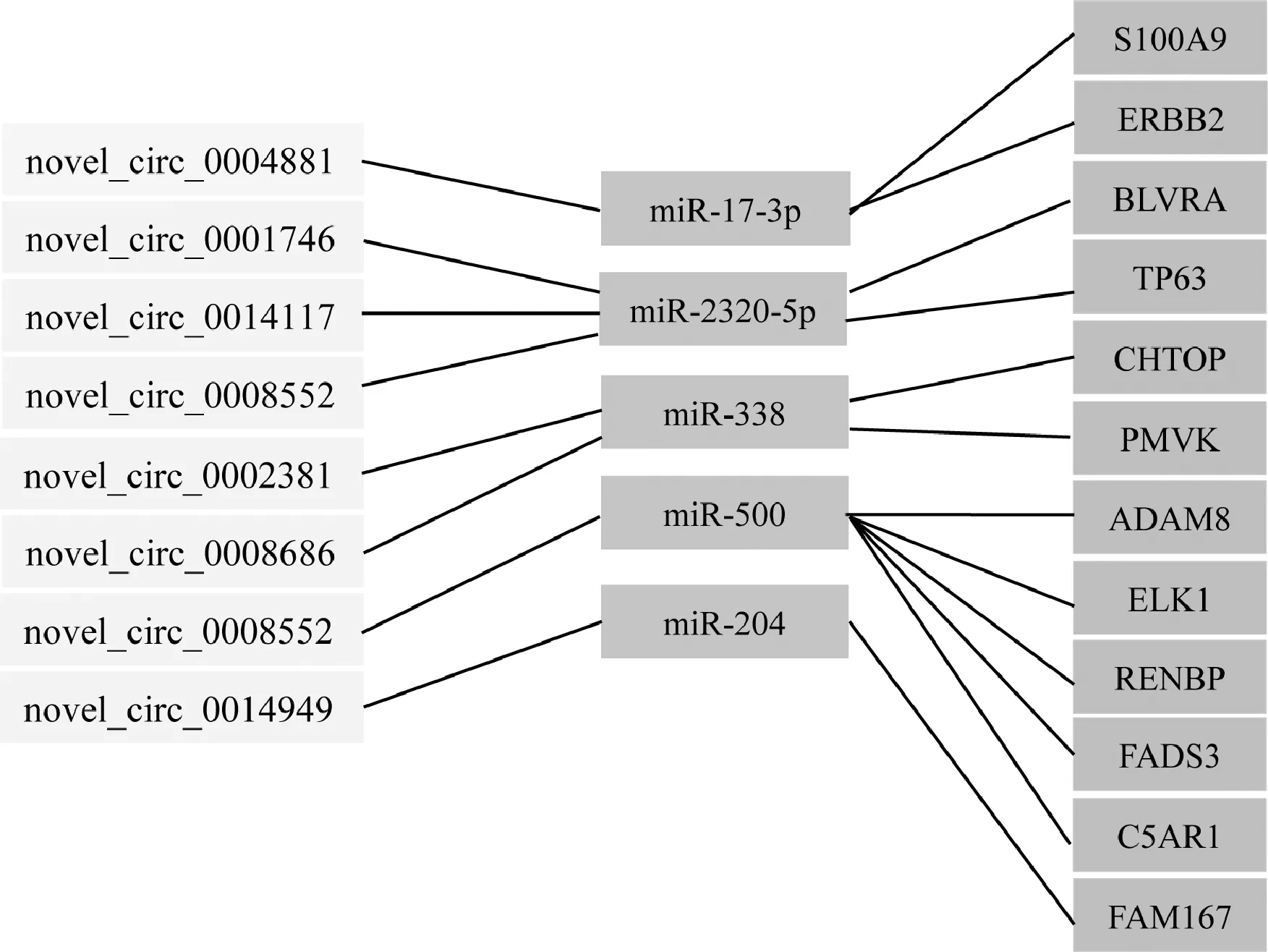
圖6 circRNA相關(guān)的ceRNA網(wǎng)絡(luò)與CAED之間的潛在關(guān)系
LPS刺激能夠激活巨噬細(xì)胞miR-500的表達(dá)上調(diào),通過(guò)靶向MFN2抑制TGF-β/Smad的激活,促進(jìn)細(xì)胞增殖和活化[43]。Wang等[29]研究發(fā)現(xiàn),miR-500通過(guò)靶向調(diào)控ELK1、HSPA2、IL7R基因參與抵制Cp感染引起的仔豬疾病。上調(diào)表達(dá)的miR-500能通過(guò)抑制靶基因的表達(dá),激活泛素偶聯(lián)受體相互作用蛋白1(RIP1)和NF-κB, 促進(jìn)癌細(xì)胞的增殖和存活[44]。C5AR1是一種炎癥驅(qū)動(dòng)因子,巨噬細(xì)胞能通過(guò)C5AR1的先天免疫反應(yīng),增加TNF-α炎癥反應(yīng),加劇機(jī)體炎癥因子的產(chǎn)生。補(bǔ)體在AOM/dss誘導(dǎo)的小鼠CRC模型的炎癥組織中被廣泛激活,導(dǎo)致多方面的后果。高表達(dá)的C5AR1可能通過(guò)調(diào)節(jié)某些關(guān)鍵細(xì)胞因子和趨化因子的產(chǎn)生,損傷CD8+T細(xì)胞,誘導(dǎo)腸道炎癥的發(fā)生,可能是腸道炎癥和腸癌發(fā)生的主控調(diào)控因子[45]。FADS3被認(rèn)為是B細(xì)胞特異性基因,能夠特異性結(jié)合NK-κB、p63轉(zhuǎn)錄因子,參與機(jī)體的細(xì)胞增殖、氧化應(yīng)激反應(yīng)[46]。本研究中,circ-0008552和miR-500在Cp感染導(dǎo)致仔豬回腸組織中的表達(dá)顯著上調(diào),C5AR1和FADS3基因的表達(dá)顯著下調(diào),因此推測(cè),circ-0008552可能競(jìng)爭(zhēng)性結(jié)合miR-500,負(fù)調(diào)控C5AR1和FADS3基因的表達(dá),參與Cp感染仔豬致腹瀉的過(guò)程。miR-204可以通過(guò)靶基因特異性抑制炎癥細(xì)胞的增殖、自噬并誘導(dǎo)細(xì)胞凋亡[47],被認(rèn)為是治療免疫、炎癥和癌癥相關(guān)疾病的潛在靶點(diǎn)。本課題組前期發(fā)現(xiàn),miR-204在Cp感染腹瀉仔豬回腸組織和C型產(chǎn)氣莢膜梭菌CPB2毒素處理的腸上皮細(xì)胞IPEC-J2中均顯著高表達(dá),此外過(guò)表達(dá)miR-204促進(jìn)了豬IPEC-J2的凋亡和炎癥反應(yīng),抑制miR-204 則減弱了細(xì)胞的炎性反應(yīng)[48]。本研究中,Cp感染導(dǎo)致仔豬回腸組織miR-204的表達(dá)水平顯著上調(diào),可能促進(jìn)了Cp感染導(dǎo)致的仔豬腸道炎癥及腹瀉癥狀。
4 結(jié) 論
本研究綜合分析了C型產(chǎn)氣莢膜梭菌感染的仔豬回腸組織 circRNA差異表達(dá)譜,篩選出694個(gè)差異表達(dá)circRNAs,主要富集在TGF-β、T細(xì)胞受體、MAPK等信號(hào)通路,構(gòu)建出由8 circRNAs-5 miRNAs-12 mRNAs組成的ceRNA網(wǎng)絡(luò)與C型產(chǎn)氣莢膜菌感染致仔豬腹瀉相關(guān),其中circ-0004881-miR-17-3p-S100A9可能在Cp感染引起的仔豬腹瀉過(guò)程中發(fā)揮潛在功能。
猜你喜歡
英語(yǔ)世界(2023年10期)2023-11-17 09:19:16
汽車實(shí)用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂(lè)探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國(guó)特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國(guó)非營(yíng)利評(píng)論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語(yǔ)文(2016年21期)2016-05-25 13:13:50

- 畜牧獸醫(yī)學(xué)報(bào)的其它文章
- 3株H3N2亞型禽流感病毒的基因組特征與演化分析
- 鼠傷寒沙門菌伴侶蛋白Hfq與小RNA GcvB結(jié)合位點(diǎn)的初步分析
- 基于網(wǎng)絡(luò)藥理學(xué)探討黃芩素對(duì)豬丁型冠狀病毒感染的潛在作用機(jī)制
- 阿司匹林丁香酚酯對(duì)體外脂多糖誘導(dǎo)小鼠巨噬細(xì)胞炎癥反應(yīng)的抑制效應(yīng)
- 硒缺乏通過(guò)TLR4/MyD88/NF-κB信號(hào)通路介導(dǎo)雞胸腺細(xì)胞凋亡
- 奶牛牧場(chǎng)養(yǎng)殖環(huán)境中產(chǎn)ESBL耐藥菌的流行特征