鼠傷寒沙門菌伴侶蛋白Hfq與小RNA GcvB結合位點的初步分析
2022-11-29 13:40:10令狐遠鳳段世宇張家莉張寶太
畜牧獸醫(yī)學報 2022年11期
令狐遠鳳,潘 永,楊 陽,段世宇,張家莉,張寶太,楊 琦*
(1.貴州大學動物科學學院, 貴陽 550025;2.貴州大學動物疫病研究所,貴陽 550025;3.貴州省農(nóng)業(yè)科學院畜牧獸醫(yī)研究所,貴陽550025)
STM sRNA GcvB是一類重要的RNA分子,其無法編碼相應的蛋白質,而是通過與Hfq協(xié)同作用,轉錄后調控STM有關靶mRNA[1]。現(xiàn)已經(jīng)驗證確認受GcvB直接調控的靶mRNA在STM中近30個,其中,大部分靶mRNA與GcvB的結合位點也已闡明[2]。sRNA-Hfq -mRNA三聯(lián)體在細菌中普遍存在,Hfq不僅能夠維持sRNA在細菌中轉錄后的穩(wěn)定性,而且可重塑sRNA與其靶mRNA的二級結構構象,使sRNA易于與其靶mRNA結合[3]。Hfq六聚體通常利用其近端面與sRNA富含U的序列結合并作用[4]。
STM Hfq具有維持GcvB在體內穩(wěn)定性的作用,但Hfq與GcvB具體的作用結合位點尚未探究透徹。鑒于此,本研究將利用RNA二級結構預測軟件分析GcvB二級結構并篩查Hfq結合偏好型的富U基序;利用λ-Red同源重組酶系統(tǒng)構建相應序列的突變或截短菌株;利用噬菌體轉導技術構建相應的hfq或gcvB基因缺失菌株以及l(fā)acZ基因融合菌株;通過qRT-PCR檢測不同重組菌株gcvB基因的轉錄水平;通過β-半乳糖苷酶試驗檢測不同重組菌株oppA基因的蛋白水平,最后分析Hfq與GcvB富U基序的作用關系,為闡明STM Hfq與GcvB的作用機理提供理論依據(jù)。
1 材料與方法
1.1 主要實驗材料
MA7455菌株(STM LT2/pKD46),MA10241菌株(STM LT2 ΔgcvB::cat),MA10675菌株(STM LT2 Δhfq::tetRA),P22噬菌體均由法國國家科學研究中心(CNRS)分子遺傳學Bossi實驗室惠贈。STM LT2oppA::lacZ菌株由本實驗室構建。氯霉素、鹽酸四環(huán)素、卡那霉素、dNTPs Mix及ONPG均購自北京索萊寶科技有限公司。AceQ Universal SYBR qPCR Master Mix和膠回收試劑盒均購自南京諾唯贊生物科技有限公司。NovoScript?Plus All-in-one 1 st Strand cDNA Synthesis SuperMix (gDNA Purge)購自近岸蛋白生物科技有限公司。
1.2 引物設計與合成
登錄NCBI網(wǎng)站(https://www.ncbi.nlm.nih.gov/),獲取STM LT2株的全基因序列(GeneID:AE006468.2), 根據(jù)基因序列設計引物(表1),引物由Primer select軟件設計,并由上海生工生物工程有限公司合成。

表1 PCR引物序列
1.3 GcvB二級結構預測和富U基序篩查
使用Mfold軟件預測GcvB二級結構。登錄網(wǎng)站(http://unafold.rna.albany.edu/?q=mfold),打開RNA Folding Form (Version 2.3 energies)選項,輸入gcvB基因完整序列,其余參數(shù)按默認方案設置,導出并分析結果。篩查GcvB中Hfq結合偏好型富U基序。
1.4 GcvB富U基序突變或截短菌株的構建
利用λ-Red同源重組酶系統(tǒng),以P1/P2為引物,MA10241菌株總DNA為模板,PCR擴增后產(chǎn)物回收作為模板,P3/P10為引物,產(chǎn)物經(jīng)純化后與pKD46感受態(tài)細胞進行電轉,在平板上培養(yǎng)后以P8/P9為引物鑒定其菌落,篩選出STM LT2gcvB::cat;以P4/P10為引物,PCR擴增用于替換gcvB基因內部為T的目的片段,分別以P5/P10、P6/P10、P7/P10和P8/P10為引物,PCR擴增用于截短gcvB基因T8的目的片段,用上述相同方法將鑒定正確的菌株分別命名為STM LT2gcvB(130—134TTTTT>GGGGG)::cat、STM LT2gcvBΔ(206)::cat、STM LT2gcvBΔ(205—206)::cat、STM LT2gcvBΔ(204—206)::cat和STM LT2gcvBΔ(203—206)::cat;以STM LT2gcvBΔ(204—206)::cat菌株總DNA為模板,P1/P2為引物,其產(chǎn)物為模板,P9/ P10為引物,以相同方法鑒定正確的菌株命名為STM LT2gcvB[Δ(203—206),(184—185)::CCCC, (192—193)::GGGG]::cat。
1.5 P22噬菌體的轉導
將鼠傷寒沙門菌MA10675和STM LT2oppA::lacZ缺失菌株作為供體菌株,GcvB 富U基序突變或截短菌株為受體菌,分別取受體菌100、50 μL稀釋后供體菌borth至2.0 mL EP 管中混勻置于37 ℃搖床孵育1 h后涂布篩選,以引物P10/P11、P6/P7分別對hfq基因的缺失、oppA-lacZ融合基因進行鑒定。
1.6 qRT-PCR反應
根據(jù)R6950 Bacterial RNA Kit,提取鼠傷寒沙門菌LT2的RNA,以反轉錄得到的cDNA作為模板,P11/P12(gcvB基因)和 P13/P14(16S rDNA基因)為引物,按照SYBR?Premix Ex TaqTMII(Tli RNaseH Plus)試劑盒說明書進行qPCR,以16S rDNA基因為內參基因,以STM LT2gcvB::cat菌株為對照。最終采用相對比較法即Ct法(Qr=2-ΔΔCt)計算分析數(shù)據(jù)[5]。
1.7 β-半乳糖苷酶活性的檢測
取50 μL待測菌株置于5 mL LB中,培養(yǎng)至 OD600 nm=0.4后,取0.1 mL至新試管中,以Z buffer定容到1 mL。滴加3滴甲苯后在42 ℃水浴搖床中振蕩2 h,將試管轉于28 ℃ 水浴鍋加入0.2 mL ONPG,記錄加入時間(t0)。當試管中呈現(xiàn)出黃色時,加入0.5 mL Na2CO3終止反應,記錄終止時間(tf)。將試管中終止所得液體離心后測其OD420nm,記錄數(shù)值。通過公式計算β-半乳糖苷酶活性。
β-半乳糖苷酶活性=(1 000×OD420 nm)/[(tf-t0)×OD600 nm]
2 結 果
2.1 GcvB二級結構預測及富U基序篩查結果
使用Mfold軟件預測STM LT2 GcvB的二級結構。結果顯示,GcvB堿基可退火形成6個莖環(huán)結構,查找到的兩個Hfq結合偏好型基序,U5位于第4個與第5個莖環(huán)之間,U8位于GcvB 3′末端,且均處于單鏈狀態(tài)。
2.2 GcvB U5基序突變菌株/U8基序截短菌株及其hfq基因缺失菌株的構建結果
利用λ-Red同源重組酶系統(tǒng)和噬菌體轉導技術構建相應的hfq基因缺失菌株。以STM LT2gcvB::cat菌株DNA為模板,利用λ-Red同源重組酶系統(tǒng)將gcvB基因T5(130—134 nt)突變?yōu)镚5,將gcvB基因T8(199—207 nt)截短。PCR鑒定結果顯示,所有重組菌株均出現(xiàn)與預期相符的918、1 964 bp目的條帶,將PCR測序后進行峰圖比對,STM LT2gcvB基因T5(130—134 nt)已突變?yōu)镚5,STM LT2gcvB基因T8(199—207 nt)已分別被截短突變?yōu)門7、T6、T5和T4。
2.3 gcvB T5基序突變對gcvB轉錄水平的影響
經(jīng)過qRT-PCR試驗結果顯示,與野生型菌株(WT)相比,T5基序突變后gcvB基因轉錄水平未產(chǎn)生明顯變化,敲除hfq基因后gcvB基因轉錄水平下調52%,敲除hfq基因的基礎上T5基序的突變使gcvB基因轉錄水平下調51%。以上結果表明,hfq與gcvB基因轉錄水平呈正相關,但gcvBT5基序與gcvB基因的轉錄水平無明顯關聯(lián)。
2.4 gcvB T8基序截短對GcvB轉錄水平的影響
對gcvBT8基序截短菌株(T7、T6、T5和T4)及相應的hfq基因缺失菌株的gcvB轉錄水平進行檢測。qRT-PCR檢測結果顯示,與WT相比,T7和T6菌株gcvB基因轉錄水平未產(chǎn)生明顯變化,而T5和T4菌株分別下調40.5%和37.5%。敲除hfq基因的基礎上截短T8基序,與WT相比,T7、T6、T5和T4菌株分別下調58.5%、57.8%、75.2%和77.8%(圖1)。以上結果表明,gcvB基因T8基序的截短對其轉錄水平產(chǎn)生了抑制作用,截短為T5和T4時作用最為明顯。
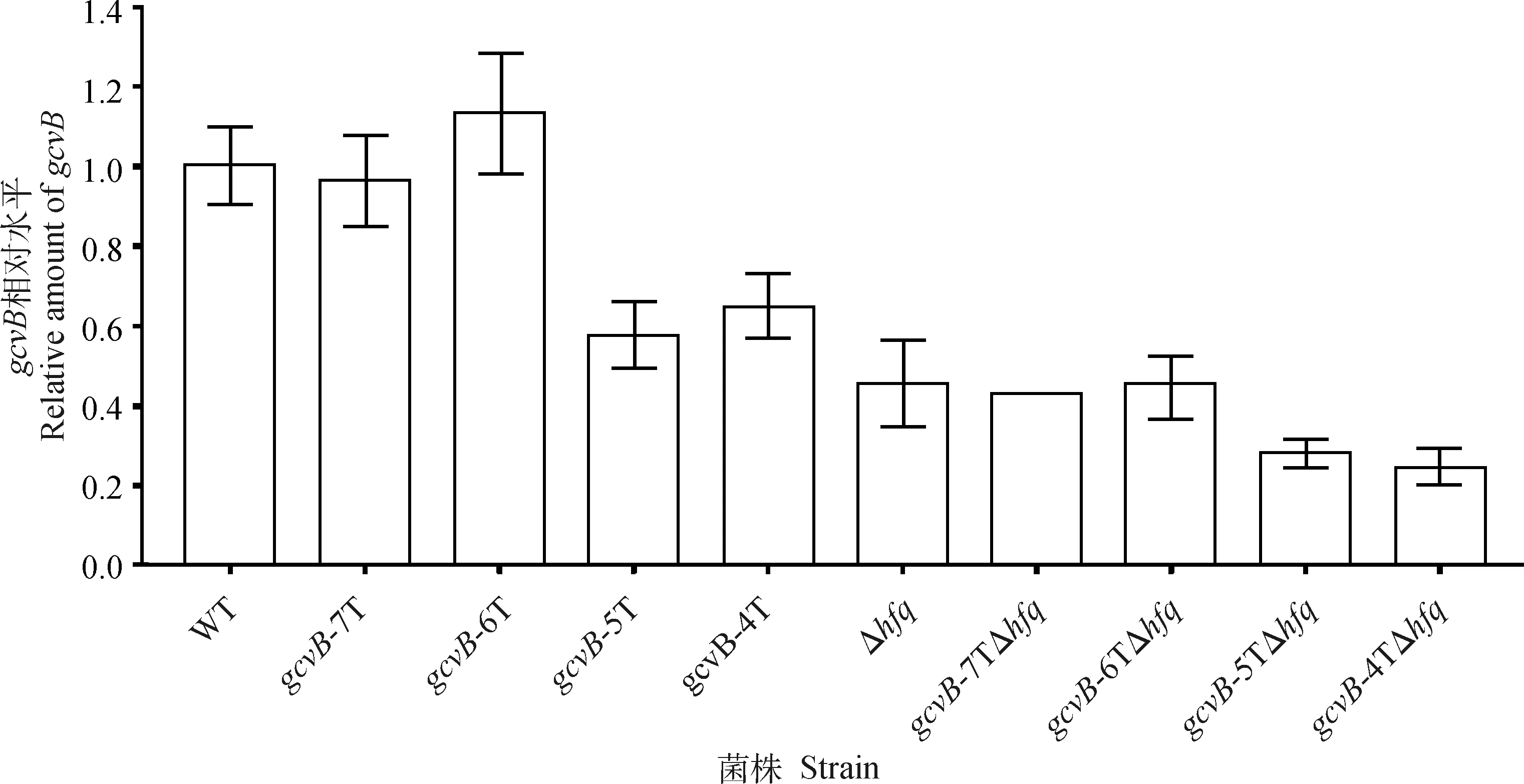
WT. STM LT2 gcvB::cat; gcvB-7T. STM LT2 gcvB Δ(206)::cat; gcvB-6T. STM LT2 gcvB Δ(205—206)::cat; gcvB-5T. STM LT2 gcvB Δ(204—206)::cat; gcvB-4T. STM LT2 gcvB Δ(203—206)::cat; Δhfq. STM LT2 gcvB::cat Δhfq::tetRA; gcvB-7TΔhfq. STM LT2 gcvB Δ(206)::cat Δhfq::tetRA; gcvB-6TΔhfq. STM LT2 gcvB Δ(205—206)::cat Δhfq::tetRA; gcvB-5TΔhfq. STM LT2 gcvB Δ(204—206)::cat Δhfq::tetRA; gcvB-4TΔhfq. STM LT2 gcvB Δ(203—206)::cat Δhfq::tetRA
2.5 GcvB終止子莖延伸菌株及其hfq基因缺失菌株的構建結果
利用λ-Red同源重組酶系統(tǒng)在gcvB基因185—186位堿基之間插入C4序列,191—192位堿基之間插入G4序列,構建GcvB終止子莖延伸菌株,在此基礎上,利用噬菌體轉導技術構建hfq基因缺失菌株。PCR產(chǎn)物測序峰圖比對表明目的片段已正確構建。
2.6 終止子莖延伸對GcvB轉錄水平的影響
對STM LT2gcvBΔ(203—206)::cat菌株GcvB終止子莖延伸菌株及其hfq基因缺失菌株的gcvB轉錄水平進行檢測。qRT-PCR檢測結果顯示,與WT相比,gcvB-LS4T終止子莖延伸菌株gcvB基因轉錄水平略微下調19.6%,而在此基礎上敲除hfq基因后,gcvB基因轉錄水平下調29%。與僅敲除hfq基因時相比,gcvB-LS4TgcvBΔhfq基因轉錄水平上調為原來的1.6倍(圖2)。以上結果表明,終止子莖的延伸與gcvB基因的轉錄水平呈正相關。
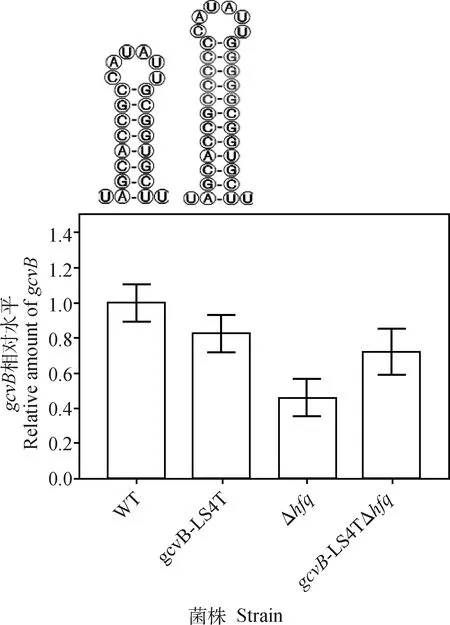
WT. STM LT2 gcvB::cat; gcvB-LS4T. STM LT2 gcvB[Δ(203—206),(184—185)::CCCC, (192—193)::GGGG]::cat; Δhfq. STM LT2 gcvB::cat Δhfq::tetRA; gcvB-LS4TΔhfq. STM LT2 gcvB[ Δ(203—206),(184—185)::CCCC, (192—193)::GGGG]::cat Δhfq::tetRA
2.7 oppA::lacZ基因融合菌株的構建結果
以STM LT2oppA::lacZ菌株作為供體菌,STM LT2gcvB::cat、STM LT2gcvBΔ(203—206)::cat和STM LT2gcvB[Δ(203—206),(184—185)::CCCC, (192—193)::GGGG]::cat分別為受體菌,運用噬菌體轉導技術構建相應的oppA::lacZ基因融合菌株。PCR鑒定結果顯示,所有重組菌株均出現(xiàn)與預期相符的918、914、922、250 bp目的條帶(圖3)。
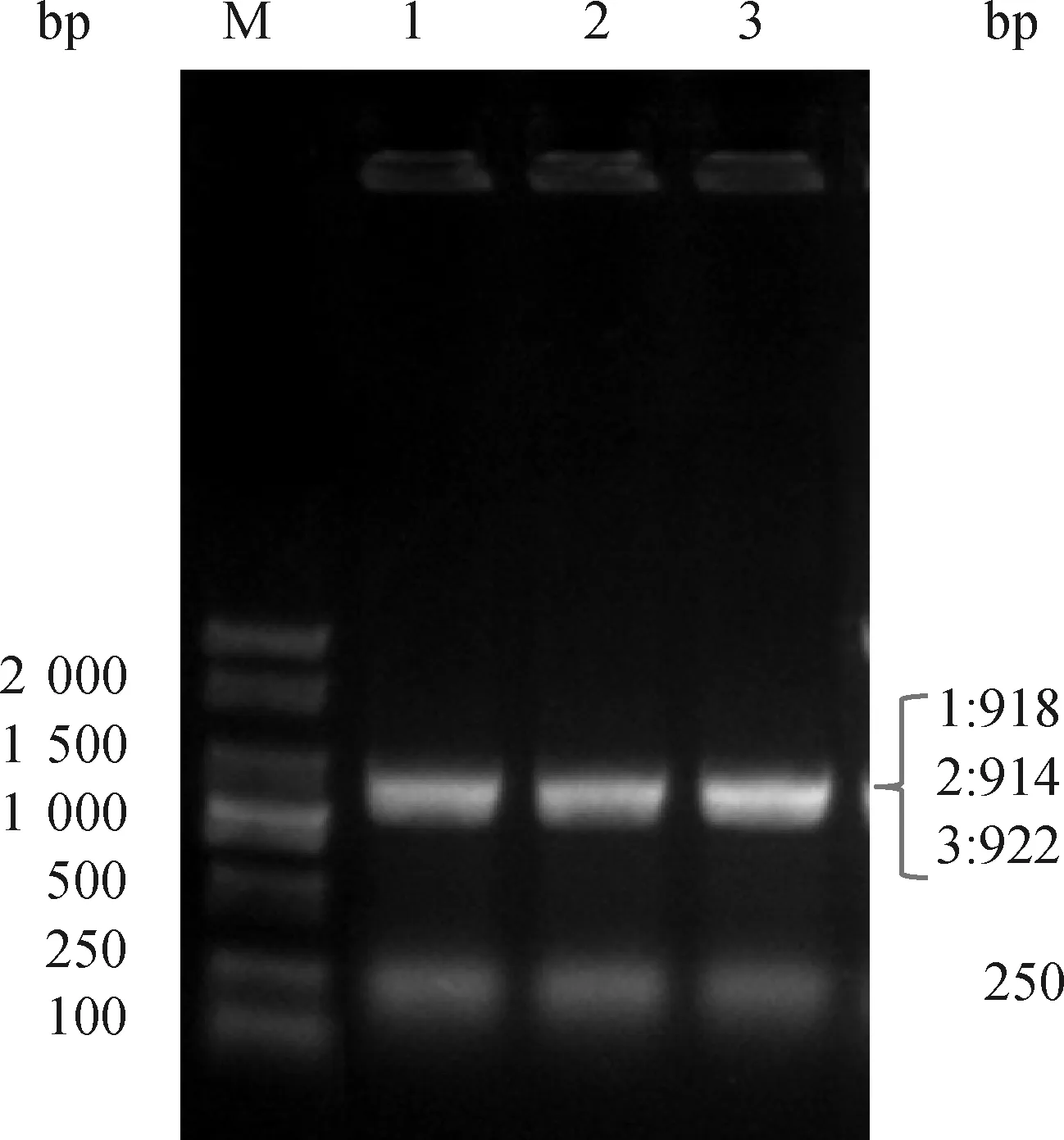
M. DL2000 marker; 1. STM LT2 gcvB::cat oppA::lacZ; 2. STM LT2 gcvB Δ(203—206)::cat oppA::lacZ; 3. STM LT2 gcvB[Δ(203—206),(184—185)::CCCC, (192—193)::GGGG]::cat oppA::lacZ
2.8 GcvB終止子莖延伸對oppA蛋白水平的影響
通過檢測oppA::lacZ基因融合菌株β-半乳糖苷酶活性,探究gcvBT8截短為T4以及延伸終止子莖對OppA蛋白表達的影響。結果顯示,與WT相比,gcvB基因 T8截短為T4后,oppA基因蛋白水平上調為原來的1.7倍。延伸該菌株的GcvB終止子莖后oppA基因蛋白水平上調為原來的1.7倍。以上結果表明,GcvB終止子莖的延伸雖恢復了因gcvB基因T8基序截短為T4時gcvB基因轉錄水平的下調,卻未恢復下調OppA蛋白表達的能力。
3 討 論
Sauer和Weichenrieder[6]通過分析尺寸排阻色譜法證實了大腸桿菌Hfq與sRNA RybB的3′末端U6具有很高的親和力,由于大多數(shù)Hfq依賴型sRNA 3′末端都具有4~8個尿嘧啶,該研究揭示了一種重要的Hfq與sRNA作用模式。Otaka等[4]發(fā)現(xiàn)截短大腸桿菌sRNA SgrS 3′末端U8時,SgrS喪失了與Hfq結合并使ptsG mRNA沉默的能力,再次證實了sRNA 3′末端polyU對Hfq結合的重要性。本研究中,對沙門菌sRNA GcvB二級結構預測,考慮到U5處于GcvB內部且在莖環(huán)結構附近,缺失將嚴重引起二級結構的改變從而增加了變量,不利于試驗分析GcvB與Hfq之間的作用關系,因此對U5進行了突變處理,對U8進行截短處理。先前的研究已表明,Hfq主要與SgrS 3′末端U8結合,從而增強了SgrS的穩(wěn)定性。試驗檢測了突變的U5及截短的U8菌株gcvB基因的轉錄水平,結果顯示,U5的突變并未引起gcvB基因轉錄水平的顯著變化,而U8的截短導致gcvB基因轉錄水平的明顯變化,當GcvB 3′末端尿嘧啶數(shù)量縮減為5個時,GcvB轉錄水平開始表現(xiàn)出明顯的下調。經(jīng)初步驗證,U8基序對于Hfq維持GcvB的穩(wěn)定性以及協(xié)助GcvB負調控OppA的表達是必要的。
由于懷疑GcvB U8基序的縮短破壞了gcvB的轉錄終止進程,本研究在GcvB U8基序截短為U4的基礎上延伸了U8基序前的終止子莖,與未延伸終止子莖相比,GcvB轉錄水平上調,幾乎恢復至野生型菌株的水平,在此基礎上敲除hfq基因也并未使GcvB轉錄水平發(fā)生明顯變化,僅略微下調。但結果仍不能充分說明GcvB U8基序截短為U4GcvB轉錄水平下調的原因是破壞了gcvB的轉錄終止進程或是破壞了GcvB與Hfq結合的關鍵基序,研究推測兩種方式均存在。而通過對oppA基因蛋白水平的檢測,發(fā)現(xiàn)GcvB U4菌株和其終止子莖延伸菌株oppA基因蛋白水平均較野生型菌株上調,該結果提示盡管終止子莖的延伸恢復了GcvB轉錄水平,但其抑制oppAmRNA的翻譯的能力已然喪失,而Hfq是GcvB發(fā)揮調控作用依賴的伴侶蛋白。結果進一步說明GcvB U8基序對于Hfq維持GcvB的穩(wěn)定性是重要的。
4 結 論
U5基序與Hfq維持GcvB穩(wěn)定性無明顯關聯(lián)。U8基序對Hfq協(xié)助GcvB轉錄后負調控oppAmRNA以及維持GcvB穩(wěn)定性來說是重要的,推測其為Hfq與GcvB的作用結合位點。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
學苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
