齊墩果酸抑制骨關節炎大鼠肌肉氧化損傷及軟骨下骨異常骨重建
2022-11-29 13:20:34馬天文呂良鈺賈麗娜湯繼浪趙明超王新宇張建濤
畜牧獸醫學報 2022年11期
馬天文,呂良鈺,于 躍,賈麗娜,陳 鴻,湯繼浪,趙明超,王新宇,張建濤,高 利
(東北農業大學動物醫學學院,黑龍江省動物疾病致病機制與比較醫學重點實驗室,哈爾濱 150030)
骨關節炎(osteoarthritis,OA)是一種退行性全關節疾病。早期階段動物表現為關節疼痛、腫脹或跛行等臨床癥狀,病情加重后可出現關節僵硬、肌肉無力,功能障礙甚至致殘[1]。作為動物臨床高發的關節疾病,OA導致的跛行嚴重影響家畜使役,促使家畜過早淘汰,也影響動物健康和生活質量[2]。目前,OA發病機制不明確且治療手段十分有限,獸醫臨床上使用非甾體抗炎藥、COX-2抑制劑等藥物僅能夠緩解疼痛。而且長期使用會導致胃腸道和心血管不良反應以及肝功能損傷[3]。因此迫切需要一種有效、安全的策略來預防或治療OA。
齊墩果酸(oleanolic acid,OLA)是從女貞子果實等植物中分離的五環三萜類化合物。OLA具有抗炎、抗氧化和護肝等藥理作用。有研究表明,OLA通過miR-148-3p/FGF2通路減輕IL-1β誘導的軟骨細胞功能障礙[5]。OLA可以激活SIRT3通過抑制成纖維細胞樣滑膜細胞中的NF-κB信號通路來抑制滑膜炎癥[6],并且抑制OA大鼠軟骨MMP-3的表達。這些研究表明,OLA對OA軟骨和滑膜炎癥具有積極作用,但其對于OA導致的肌肉功能障礙和軟骨下骨異常骨重建的作用還有待于挖掘。
軟骨下骨異常骨重建是OA軟骨下骨的特征性病變,現有研究表明,其可能是OA發病的始發因素[8-9],表現為軟骨下骨板厚度減少、孔隙率增加、軟骨下骨小梁退化、軟骨下骨囊腫、骨贅形成等特征。此外,OA患畜臨床表現的肌肉無力,主要影響股四頭肌[10]。股四頭肌功能障礙可能早于關節疼痛和影像學形態改變的發生[11],并直接參與OA發病機制[12]。藏紅花素通過抑制肌肉氧化損傷和炎癥反應,減輕大鼠關節疼痛和肌肉功能障礙[13]。同時大量報道顯示,靶向調控Nrf2/NQO1/HO-1通路抑制肌肉氧化損傷是治療OA的潛在策略[14-15]。因此本研究通過ACLT+PMMx方法建立大鼠OA模型,以闡述OLA對OA大鼠肌肉功能障礙和軟骨下骨異常骨重建的作用及可能的分子機制,為下一步的藥物研發和臨床應用提供理論支持。
1 材料與方法
1.1 實驗動物
Sprague-Dawley大鼠購自長春易思實驗動物技術有限公司。動物飼養管理和試驗操作均遵循東北農業大學動物實驗倫理審查相關規定。
1.2 主要試劑
齊墩果酸(純度≥98%,CAS:508-02-1)購自成都曼思特生物科技有限公司。手術縫線購自上海浦東金環醫療用品股份有限公司。異氟烷購自北京友誠盛達生物科技有限公司。大鼠CTX-Ⅰ、MHC、CS、OCN、IL-1和IL-6 ELISA試劑盒購于上海酶聯生物科技有限公司。GAPDH抗體和NQO1抗體購于武漢愛博泰克生物科技有限公司,HO-1抗體購于沈陽萬類生物科技有限公司,Nrf2抗體購于Affinity公司,高敏型ECL化學發光檢測試劑盒購于南京諾唯贊生物科技股份有限公司。
1.3 主要儀器
外科顯微鏡(科奧達,中國);Epoch酶標儀(BioTek,美國);大鼠足底觸敏痛覺測試儀(UGO,意大利);Micro-CT(Bruker,德國)。
1.4 大鼠OA模型的建立
將18只大鼠隨機分為3組:對照組(CON組)、模型組(OA組)和給藥組(OLA組),每組6只。大鼠適應1周后進行試驗,OA組和OLA組的大鼠通過ACLT+PMMx方法[16]建立OA模型:大鼠異氟烷吸入麻醉,在右膝內側切開2~4 cm切口,髕骨外側脫位后膝關節完全屈曲,外科顯微鏡下切斷前十字韌帶,進行抽屜試驗確保韌帶完全切斷。并以前十字韌帶脛骨起點為圓點外旋45°切除內側半月板的1/3,無菌生理鹽水沖洗關節腔。可吸收線縫合傷口,消毒后放入籠內自由活動。CON組進行假手術,只切開關節囊后縫合,避免損傷其他韌帶和半月板。術后大鼠連續3 d給予適量抗生素。
1.5 給藥及樣本采集
OLA組大鼠手術后每日灌胃50 mg·(kg·d)-1OLA,連續灌胃4周。OLA溶于含有20 mL·L-1Tween 80的無菌生理鹽水中,OLA劑量的選擇參考之前的研究[17]。CON組與OA組灌胃等體積的含20 mL·L-1Tween 80的無菌生理鹽水。所有大鼠分別于造模后第0、7、14和28天進行疼痛行為學測試。試驗第28天,采集大鼠血液樣本后1 000g離心20 min,收集上清液進行血清ELISA試驗。試驗第28天所有大鼠實施安樂死,收集各組大鼠右側包括股直肌在內的股四頭肌樣本,注意避免包含任何脂肪組織或其他肌肉,進行Western blot和肌肉ELISA試驗。收集膝關節樣本,進行Micro-CT掃描。
1.6 疼痛行為學
1.6.1 伸膝發聲試驗 根據前人的方法[18],通過大鼠發聲次數反映膝關節疼痛情況。從靜止姿勢、膝略微彎曲開始測試。首先保定大鼠,伸展膝關節,保證膝關節完全伸展,將膝蓋夾在大拇指和食指之間,在內側方向對膝關節擠壓,上下左右伸膝。大鼠發聲記“X”,不發聲記“O”,每次間隔5 min,共測5次,記錄發聲次數之和。
1.6.2 熱敏感試驗 根據前人的方法[19]。在每次試驗開始前,將大鼠放在玻璃板上方的不透明塑料室中15 min以適應環境。將大鼠足底觸敏痛覺測試儀上的“十”字形標記置于大鼠左后跖足底中央,但避開足墊。接下來開啟儀器,從開始至大鼠出現抬腿回避的時間作為大鼠的足熱痛敏潛伏期(paw withdrawal latency, PWL)。每5 min重復1次,每只大鼠檢測3次取平均值。大鼠每次接受輻射熱刺激最長時間為20 s,以避免損傷其他組織。
1.7 ELISA試驗
采用ELISA試劑盒測定各組大鼠肌肉中檸檬酸合酶(citrate synthase, CS)、肌球蛋白重鏈(myosin heavy chain, MHC)、白細胞介素1(interleukin 1, IL-1)和IL-6的水平,以及血清中骨鈣素(osteocalcin, OCN)和Ⅰ型膠原羧基末端肽(C-terminal typeⅠ collagen telopeptide, CTX-Ⅰ)的水平,嚴格按照ELISA試劑盒的說明書操作。
1.8 Micro-CT對軟骨下骨影像學的檢測與分析
分別對各組大鼠膝關節進行Micro-CT掃描,包括脛骨和股骨各5 mm,射線管電流200 μA,電壓85 kV,掃描分辨率10 μm,曝光時間384 ms。使用三維重建軟件NRecon(V1.7.4.2版本,德國Bruker公司)對原始圖像進行選擇區域的重建。使用CT Analyser(1.18.8.0版本,德國Bruker公司)對感興趣區域ROI進行分析。設定統一參數,由CT Analyser軟件計算出骨體積分數(BV/TV)、骨小梁數(Tb.N)、骨小梁厚度(Tb.Th)和骨礦物質密度(BMD)參數。
1.9 Western blot檢測目的蛋白
使用含蛋白酶抑制劑的RIPA裂解緩沖液提取各組肌肉的總蛋白。用BCA蛋白分析試劑盒測定蛋白濃度。將蛋白樣品加入凝膠泳道電泳,然后使用300 mA恒流,濕轉法轉移到PVDF膜上。用50 g·L-1牛血清白蛋白(BSA)封閉1 h后,在4 ℃下與一抗孵育過夜,一抗稀釋比:GAPDH(1∶2 000)、Nrf2(1∶2 000)、HO-1(1∶1 000)和NQO1(1∶2 000)。將膜與二抗(1∶3 000)室溫孵育1 h,使用高敏型ECL化學發光試劑顯影,用自動凝膠圖像分析系統顯示蛋白條帶,Image J軟件分析灰度值。
1.10 統計學分析
本試驗數據使用SPSS軟件(Windows版本19.0,USA)進行分析。數據均采用“平均值±標準偏差(SD)”表示。組間采用單因素方差分析(ANOVA),然后進行Tukey檢驗,P<0.05為差異具有統計學意義,制圖軟件為GraphPad Prism 8。P<0.05表示差異顯著,P<0.01表示差異極顯著。
2 結 果
2.1 OLA抑制OA大鼠的關節疼痛
如圖1A伸膝發聲行為學檢測結果顯示,與CON組比較,OA組造模28 d內發聲次數極顯著增加(P<0.01),與OA組比較,OLA組大鼠發生次數極顯著減少(P<0.01)。如圖1B所示,與CON組比較,OA組大鼠的PWL極顯著縮短(P<0.01),與OA組比較,OLA組大鼠的PWL極顯著增加(P<0.01)。
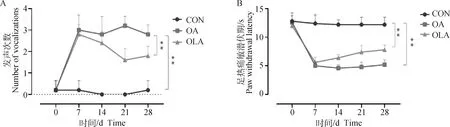
A.伸膝發聲試驗;B.熱敏感試驗;*.P<0.05,**.P<0.01,下同
2.2 OLA通過Nrf2/NQO1/HO-1通路抑制OA大鼠肌肉氧化損傷
如圖2結果顯示,與CON組比較,OA組大鼠股四頭肌組織中Nrf2和NQO1蛋白表達極顯著下降(P<0.01),OLA組中Nrf2和NQO1蛋白表達顯著(P<0.05)或極顯著(P<0.01)升高。與OA組比較,OLA組中Nrf2、NQO1和HO-1蛋白表達水平極顯著(P<0.01)或顯著(P<0.05)升高。綜上所述,OLA通過調控Nrf2/NQO1/HO-1通路抑制OA大鼠肌肉氧化損傷。
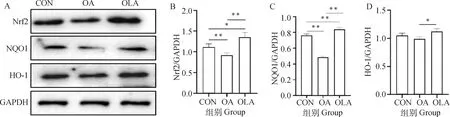
A.OLA干預對OA大鼠Nrf2/NQO1/HO-1蛋白表達的影響;B~D.各蛋白表達情況灰度分析
2.3 OLA能夠改善OA大鼠肌肉功能障礙
通過ELISA方法檢測大鼠股四頭肌組織中CS、MHC以及炎癥因子IL-1和IL-6含量變化。如圖3A~D結果顯示,與CON組比較,OA組大鼠肌肉組織中MHC和CS含量極顯著(P<0.01)或顯著(P<0.05)降低,OLA組中CS和MHC含量無明顯變化。與OA組比較,OLA組中MHC和CS含量顯著增加(P<0.05)。此外,與CON組比較,OA組大鼠肌肉組織中IL-1和IL-6含量極顯著增加(P<0.01),OLA組中IL-6含量極顯著降低(P<0.01)。與OA組比較,OLA組肌肉組織中IL-1和IL-6含量顯著(P<0.05)或極顯著(P<0.01)降低。
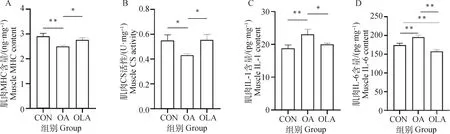
圖3 OLA能夠改善OA大鼠肌肉功能障礙
2.4 OLA抑制OA大鼠軟骨下骨的異常骨重建
如圖4所示,與CON相比,OA組中Tb.Th、BMD和BV/TV顯著(P<0.05)或極顯著(P<0.01)降低,OLA組中BV/TV極顯著降低(P<0.01)。與OA組相比,OLA組中Tb.Th、BMD和BV/TV顯著(P<0.05)或極顯著(P<0.01)增加。Tb.N在CON組、OA組和OLA組的組間變化均不顯著。
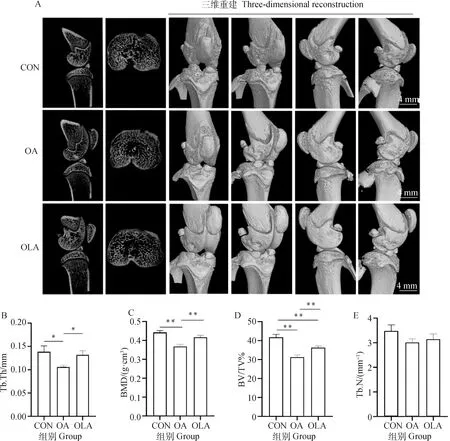
A.各組大鼠右膝關節Micro-CT掃描代表性圖像;B. Tb.Th(骨小梁厚度);C. BMD(骨礦物質密度);D. BV/TV(骨體積分數);E. Tb.N(骨小梁數)
2.5 OLA抑制OA大鼠血清中骨代謝標志物OCN和CTX-Ⅰ的表達
如圖5所示,與CON組比較,OA組大鼠血清中OCN含量顯著升高(P<0.05),CTX-Ⅰ濃度呈極顯著升高(P<0.01),OLA組中OCN和CTX-Ⅰ無顯著變化。與OA組比較,OLA組大鼠血清中OCN和CTX-Ⅰ含量顯著降低(P<0.05),其中CTX-Ⅰ濃度呈極顯著降低(P<0.01)。
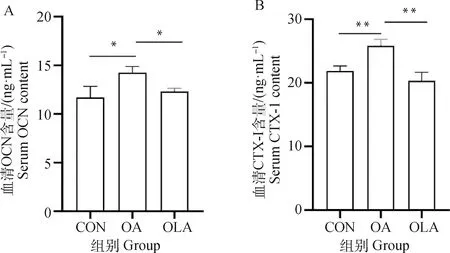
圖5 OLA抑制骨關節炎大鼠血清中骨代謝標志物OCN和CTX-Ⅰ的表達
3 討 論
OLA是一種廣泛使用的植物三萜類化合物,具有很強的抗氧化活性。以往研究表明,OLA對OA具有積極作用,但具體作用機制不明確。肌肉功能障礙和軟骨下骨異常骨重建與OA病理有關,本研究發現OLA能夠有效減輕OA大鼠疼痛反應,通過Nrf2/NQO1/HO-1通路抑制炎癥反應和肌肉氧化損傷,并且抑制軟骨下骨的異常骨重建,這為OLA治療OA機制提供了新思路。
氧化應激能夠參與OA肌肉功能障礙和炎癥的發病機制。肌肉氧化應激會使Nrf2在細胞質中積累,然后最終轉移到細胞核,啟動醌氧化還原酶-1(NQO1)和血紅素加氧酶1(HO-1)的基因轉錄。有研究表明,OA的肌肉Nrf2水平降低,而芝麻油能夠顯著增加肌肉Nrf2表達,抑制氧化應激從而減輕早期關節疼痛。本研究發現OLA干預后大鼠股四頭肌中Nrf2、HO-1和NQO1蛋白水平較OA組顯著上升,從而誘導促炎因子IL-1和IL-6的產生,這說明OLA能夠調控Nrf2/NQO1/HO-1通路抑制OA大鼠肌肉氧化損傷。此外,與OA相關的股四頭肌無力的潛在機制已被闡明[24],其中,MHC的改變在肌力中起主導作用[11]。細胞因子IL-1和IL-6以及CS與肌肉質量和肌力損失有關[25]。本研究發現ACLT+PMMx大鼠肌肉中IL-1和IL-6增加,CS活性和MHC表達降低,表明OA大鼠肌無力,持續的氧化應激導致細胞膜的脂質過氧化,損害細胞器的功能和細胞破壞最終導致炎癥反應,而OLA通過恢復CS活性和MHC含量可以改善OA中股四頭肌功能障礙。
OA是軟骨、軟骨下骨、韌帶和滑膜等所有關節結構的系統性疾病[26]。在OA中,軟骨下骨和關節軟骨之間進行信號交互。在OA早期,軟骨下骨板變薄、疏松,軟骨下骨小梁退化。在OA晚期,鈣化的軟骨和軟骨下骨板變厚,并伴有軟骨下骨小梁硬化等特征[28]。不同造模方法和造模時長軟骨下骨的變化時期有所不同。既往對OA軟骨下骨的研究大多通過切除前十字韌帶或半月板造成關節不穩而建立OA模型[31]。本研究通過ACLT+PMMx方法誘導大鼠創傷性OA,以探討軟骨下骨力學結構改變在OLA干預后的變化。有研究表明,在ACLT+MMx模型后兩周表現出顯著的軟骨下骨丟失,BV/TV下降,而在造模后6周BV/TV較對照組上升,呈現骨硬化。因此本試驗采取ACLT+PMMx模型造模4周,模擬OA早期階段,結果發現,造模后軟骨下骨BV/TV、BMD和Tb.Th均較CON組下降,而通過OLA干預后,BV/TV、BMD和Tb.Th參數顯著上升。此外,骨代謝標志物CTX-Ⅰ和CON分別是骨吸收過程中Ⅰ型膠原釋放入血的產物和成骨細胞分泌的非膠原蛋白[33]。在本研究中,OA大鼠血清中OCN和CTX-Ⅰ含量均顯著上升,OLA干預顯著降低了CTX-Ⅰ和OCN表達。這表明OLA參于OA導致的軟骨下骨異常骨重建,不僅可以改善OA發病早期階段關節軟骨下骨生物學損傷,還能夠調節其生物力學特性。OLA調節OA軟骨下骨代謝,抑制骨小梁三維結構的破壞,并緩解OA大鼠的骨侵蝕,從而有效阻止OA病理過程中軟骨下骨異常骨重建和微結構改變的可能性。但OLA如何參與軟骨下骨代謝的分子機制,以及對破骨細胞和成骨細胞的調控作用仍需進一步研究。
4 結 論
OLA可能夠抑制OA大鼠關節疼痛反應,并通過調控Nrf2/NQO1/HO-1通路抑制OA大鼠肌肉功能障礙,改善OA早期軟骨下骨生物力學性能和微結構改變,從而延緩OA大鼠軟骨下骨異常骨重建的發生。
