牛壞死桿菌43K OMP基因缺失株生物學特性分析
2022-11-29 13:20:26趙鵬宇王天碩于思雯肖佳薇賀顯晶郭東華
畜牧獸醫學報 2022年11期
蔣 凱,趙鵬宇,王天碩,于思雯,畢 欄,肖佳薇,賀顯晶,郭東華
(黑龍江八一農墾大學,大慶 163319)
壞死桿菌(Fusobacteriumnecrophorum)是一種無鞭毛、無芽孢、嚴格厭氧的多形性革蘭陰性菌,其常駐于人類或動物的口腔、消化道和泌尿生殖道[1-2]。壞死桿菌作為機會主義病原可以引起反芻動物的腐蹄病、犢牛白喉、肉牛肝膿腫、奶牛乳腺炎和馬的呼吸道感染等壞死性化膿性疾病,也會引起人化膿性血栓頸靜脈炎為主的Lemierre’s綜合征,同時可誘發結腸癌[3-5]。因此,壞死桿菌感染給畜牧業造成了巨大的經濟損失并威脅人類的健康與安全,對其致病機制的研究具有重要現實意義。
壞死桿菌的毒力因子主要包括白細胞毒素、內毒素、黏附因子和一些胞外酶等。其中白細胞毒素被認為是壞死桿菌的主要毒力因子,它對中性粒細胞、巨噬細胞和肝細胞有毒害作用,并且對人和反芻動物的中性粒細胞具有特異性損傷[6-7]。但針對壞死桿菌白細胞毒素制備的亞單位疫苗保護效果不佳,有必要對壞死桿菌其他毒力因子或功能蛋白進行拓展性研究。黏附因子也是壞死桿菌的主要毒力因子之一,外膜蛋白(out membrane proteins, OMPs)在革蘭陰性菌黏附過程中發揮重要作用,也參與營養介導、耐藥性和生物被膜形成等生物過程[8]。2013年,Sun等[9]首次在牛壞死桿菌H05菌株上鑒定43K OMP,并發現其與同菌屬的具核梭桿菌外膜蛋白FomA同源性達70.22%,且與其他梭菌屬的OMPs具有高度保守性。對比壞死桿菌的Fnn亞種和Fnf亞種的主要OMPs,發現Fnn亞種OMPs顯著性條帶為40 ku,Fnf亞種OMPs則為37.5 ku,這也解釋了Fnn亞種和Fnf亞種壞死桿菌毒性的差異[10-11]。同時,43K OMP在牛羊腐蹄病臨床分離株中廣泛存在[12]。利用抗體抑制試驗和蛋白競爭試驗發現,43K OMP在壞死桿菌黏附宿主細胞中可能發揮潛在作用,利用43K OMP截短表達制備的多抗,初步篩選了43K OMP的黏附功能區[13]。而有關于43K OMP在壞死桿菌致病中的其他生物學功能尚未可知,因此,本實驗室通過構建牛壞死桿菌43K OMP缺失株,對比其與壞死桿菌親本株的生物學特性,初步探究缺陷43K OMP后壞死桿菌的生物學特性的變化,從而確定43K OMP基因可能參與壞死桿菌致病的生物學功能,為牛壞死桿菌致病機制的研究提供理論基礎。
1 材料與方法
1.1 菌株、細胞與試驗動物
壞死桿菌A25菌株購自美國ATCC公司(Fusobacteriumnecrophorum,Fnn亞種,ATCC 25286),基因缺失株A25Δ43K OMP株通過同源重組技術制備獲得(構建同源重組手臂并通過自殺質粒導入壞死桿菌內部,從而打斷43K OMP基因)[14],小鼠單核巨噬細胞系(RAW264.7)與牛子宮內膜細胞系(BEND)由黑龍江八一農墾大學獸醫分子病理學實驗室保存;6周齡SPF級Balb/c雌性小鼠,體重18~22 g,購自長春億斯實驗動物技術有限責任公司。
1.2 主要試劑、儀器
苛養厭氧菌肉湯(FAB)培養基購自青島海博生物技術有限公司,藥敏紙片購自北京天壇生物制品股份有限公司,DMEM培養基和RPMI-1640培養基購自上海Sigma-Aldrich貿易有限公司,BeyoClickTMEdU-488細胞增殖檢測試劑盒購自上海碧云天生物技術有限公司,厭氧培養箱購自上海龍躍儀器設備有限公司,熒光顯微鏡購自上海徠卡科技有限公司。
1.3 細菌培養
將牛壞死桿菌親本株A25株與基因缺失株A25Δ43K OMP株甘油凍存菌復蘇接種于FAB培養基中(其中基因缺失株A25Δ43K OMP株培養基含有2 μg·mL-1甲砜霉素),37 ℃厭氧培養,氣體環境為:85% N2、10% H2和5% CO2,經革蘭染色鑒定后,按照1∶50傳代,傳3~4代可用于試驗。
1.4 細胞培養
RAW264.7細胞培養在含有10%胎牛血清的DMEM培養基中、BEND細胞培養在含有10%胎牛血清的RPMI-1640培養基中,當細胞密度達到80%即可進行傳代培養,細胞均在37 ℃,5% CO2細胞培養箱內培養。
1.5 生長曲線測定
將兩菌株按照1∶20比例分別接種到FAB液體培養基,于37 ℃厭氧培養,分別在培養0、2、4、6、8、10、12、18、24、30、36、42、48 h取菌液2 mL測定其600 nm的吸光度值,試驗進行3次重復,每次重復設3個平行組,記錄并繪制生長曲線。
1.6 藥敏試驗
根據CLSI標準,使用K-B紙片擴散法對兩菌株進行藥物敏感性試驗[15]。取50 μL處于對數生長期的親本株A25株和缺失株A25Δ43K OMP株菌液均勻涂布于固體培養基上,貼藥敏紙片,37 ℃厭氧培養48~72 h,觀察抑菌圈大小,測量抑菌圈直徑并記錄分析。
1.7 生物被膜形成能力測定
將親本株A25株和缺失株A25Δ43K OMP株接種至FAB培養基中,過夜培養后,將菌液用FAB培養基進行100倍稀釋,取200 μL菌液加入至96孔板內,37 ℃厭氧分別培養48、72和96 h,蒸餾水沖洗,隨后用1%結晶紫溶液染色30 min,蒸餾水沖洗后加入200 μL 95%乙醇溶液溶解30 min,檢測OD570 nm值[16],試驗進行3次重復,每次重復設3個平行組。
1.8 細胞黏附試驗
將親本株A25株和缺失株A25Δ43K OMP株接種至FAB培養基中,37 ℃厭氧培養至OD600 nm=0.6~0.8,收集細菌于室溫3 000×g離心5 min,無菌PBS洗3次,用RPMI-1640培養基重懸。同時將BEND細胞接種在12孔板中,每孔接種1×106個細胞,37 ℃,5% CO2細胞培養箱內培養12 h用于試驗,按照感染復數MOI為1∶100的比例加入細菌,共同孵育1 h。隨后用無菌PBS清洗3次除去未黏附的細菌,用1 mL無菌PBS收集細胞懸液并用連續10倍稀釋,涂布于固體培養基上37 ℃厭氧培養36~48 h,計算細菌的黏附數[17],試驗進行3次重復,每次重復設3個平行組。
1.9 細胞增殖抑制試驗
收集親本株A25株和缺失株A25Δ43K OMP株并分別用DMEM培養基重懸。同時將RAW264.7細胞接種在12孔板中,每孔接種1×106個細胞,37 ℃,5% CO2細胞培養箱內培養12 h用于試驗,按照感染復數MOI為1∶100的比例加入細菌,分別孵育2、4和6 h后,按照BeyoClickTMEdU-488細胞增殖檢測試劑盒說明書進行細胞增殖檢測,用熒光顯微鏡以400倍視野觀察,隨機取5個視野進行數據統計[18]。
1.10 缺失株對小鼠的致病性試驗
將6周齡Balb/c雌性小鼠平均分為3組,分別為:親本株A25、缺失株A25Δ43K OMP和對照組,每組10只。將菌液用無菌PBS洗3遍后重懸,200 μL腹腔接種至小鼠體內(5×108CFU·只-1)來測定存活曲線,對照組注射等量無菌PBS。從接種后開始計時,連續觀察14 d,并繪制小鼠的存活曲線。對死亡小鼠及存活14 d小鼠進行無菌剖檢,將肝組織研磨后,用無菌PBS對組織勻漿進行倍比稀釋,涂布于固體平板37 ℃厭氧培養48 h,平板計數并計算兩菌株在Balb/c雌性小鼠每克肝組織中的載菌量。
1.11 數據分析
每組試驗均重復3次,數據采用GraphPad Prism軟件One-way ANOVA進行分析,以“平均數±標準差”表示,P<0.05時判定結果差異顯著。
2 結 果
2.1 生長曲線
為了探究缺陷43K OMP基因是否會影響牛壞死桿菌的生長特性,分別檢測了兩菌株的生長曲線。結果如圖1所示,親本株A25株和缺失株A25Δ43K OMP株均在4~8 h進入對數生長期,12 h達到峰值,隨后進入平臺期,細菌密度逐漸減弱。結果顯示,缺陷43K OMP基因后牛壞死桿菌的生長特性沒有明顯差異(P>0.05)。
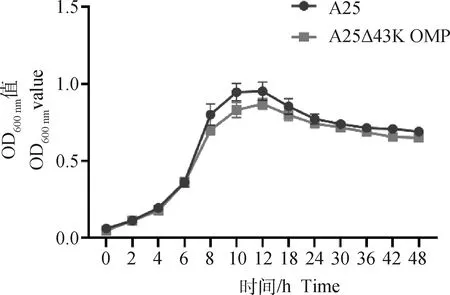
圖1 A25株和A25Δ43K OMP株生長曲線
2.2 藥敏試驗結果
為了探究缺陷43K OMP基因是否會對牛壞死桿菌的耐藥性產生影響,選用25種抗菌藥物對親本株A25株和缺失株A25Δ43K OMP株進行藥敏試驗。結果如表1所示,兩菌株對鏈霉素、桿菌肽和氟苯尼考高度耐藥,未出現抑菌圈。在缺陷牛壞死桿菌43K OMP基因后,缺失株A25Δ43K OMP株對氨芐西林、紅霉素、大觀霉素、磺胺甲噁唑和痢特靈產生的抑菌圈變小,表現出耐藥性增強;對克拉霉素、卡那霉素、新霉素、阿米卡星、奈替米星、呋喃妥因、多黏菌素B、利福平和環丙沙星產生的抑菌圈變大,表現出耐藥性減弱。
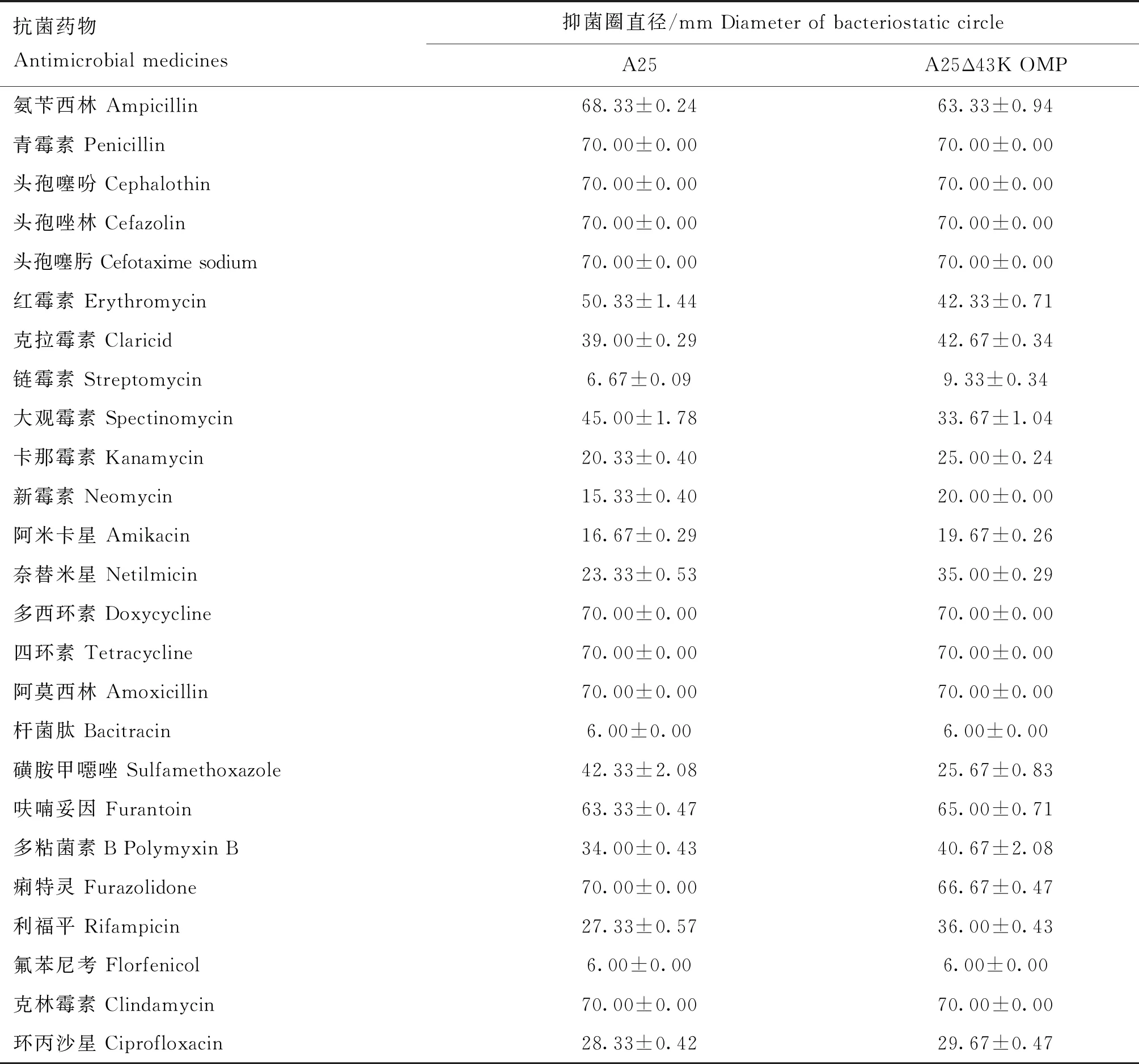
表1 A25和A25Δ43K OMP藥敏試驗結果
2.3 生物被膜檢測結果
為了探究缺陷43K OMP基因是否對牛壞死桿菌的生物被膜形成能力產生影響,用結晶紫染色法分別對兩菌株進行生物被膜測定。結果如圖2所示,接種48、72和96 h時,對照組OD570 nm分別為0.10、0.08和0.09,親本株A25 OD570 nm分別為0.40、0.49和0.50,缺失株A25Δ43K OMP OD570 nm分別為0.57、3.30和1.57。均表現為ODA25Δ43K OMP>ODA25>2×OD570 nm。結果顯示,兩菌株生物被膜形成能力均較強,且缺陷43K OMP基因后,牛壞死桿菌的生物被膜形成能力顯著增強。
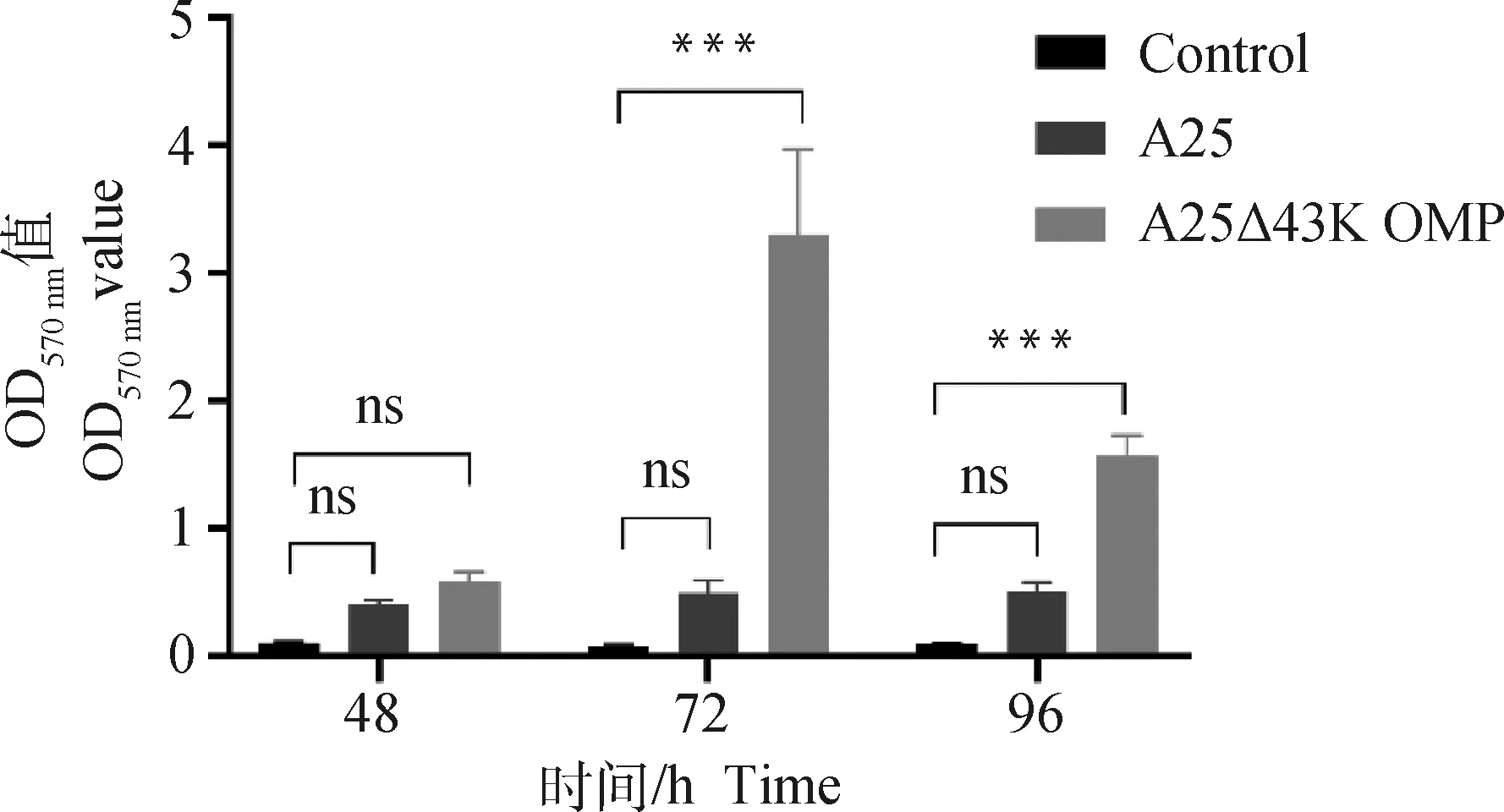
ns. P>0.05; ***. P<0.000 5。下同
2.4 細胞黏附能力測定結果
為了探究缺陷43K OMP基因是否對牛壞死桿菌的黏附能力產生影響,測定了兩菌株對BEND細胞的黏附能力。結果如圖3所示,親本株與缺失株在BEND細胞上黏附的細菌量分別為6.48×105和0.33×105CFU·mL-1。結果顯示,缺陷43K OMP基因后,可顯著降低牛壞死桿菌與BEND細胞的黏附。
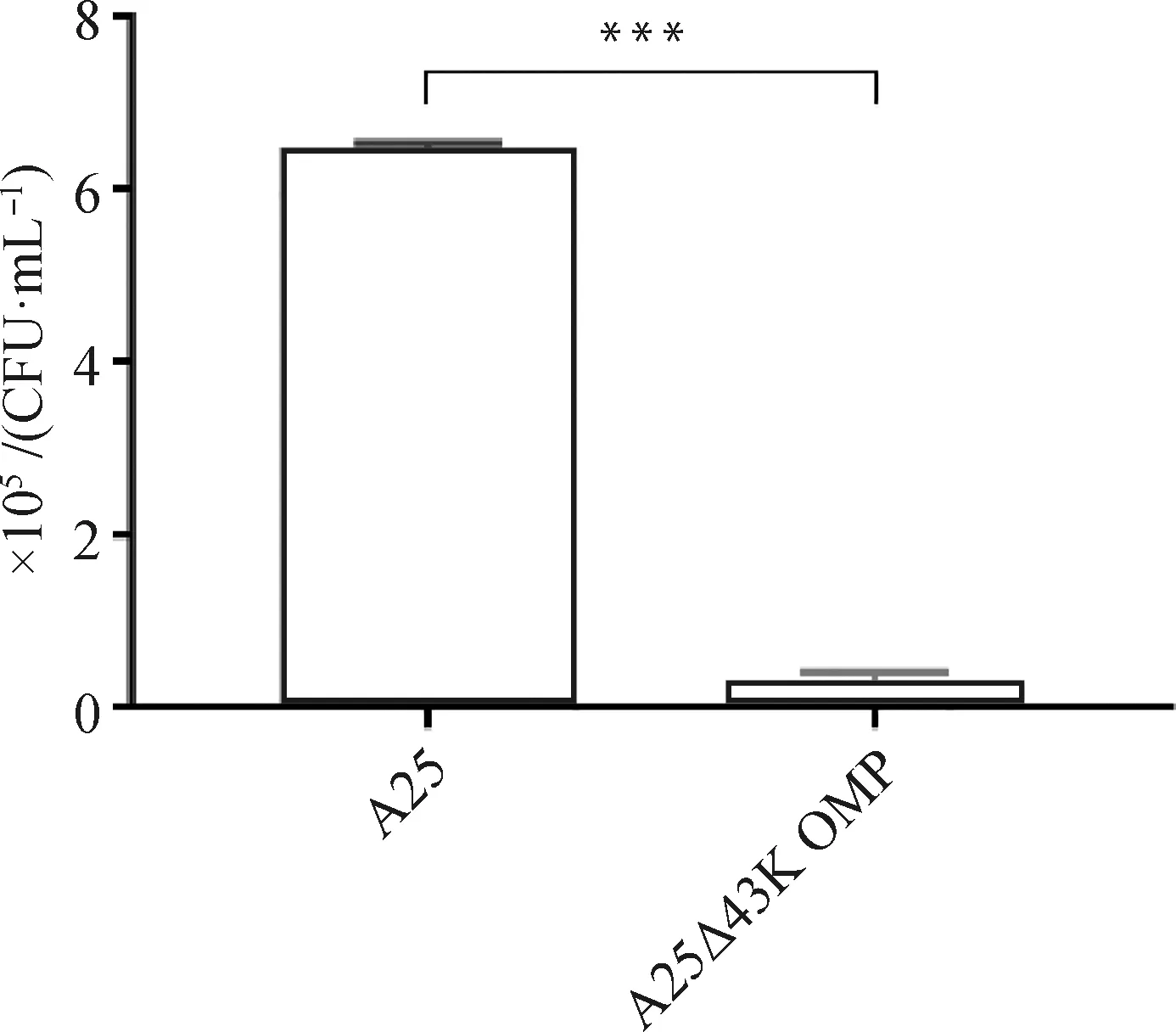
圖3 A25株和A25Δ43K OMP株對BEND細胞黏附能力
2.5 細胞毒性檢測結果
為了探究缺陷43K OMP基因是否對牛壞死桿菌的毒性產生影響,在體外分別測定兩菌株對RAW264.7細胞的增殖抑制能力。結果如圖4所示,對照組、親本株組和缺失株組的細胞增殖率在2 h分別為0.40、0.31和0.37,在4 h時分別為0.38、0.22和0.18,在6 h時分別為0.39、0.07和0.06(圖4B)。結果顯示,缺陷43K OMP基因,牛壞死桿菌對RAW264.7細胞毒性無顯著差異(P>0.05)。
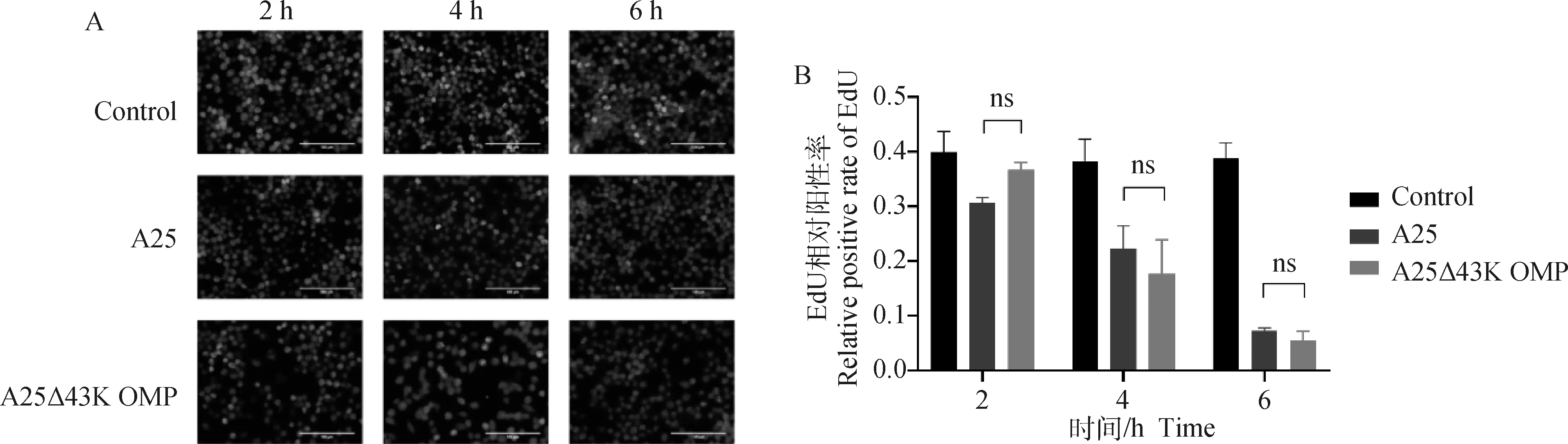
A. 從各組中隨機選擇視野并合并9張圖像(400×);B. 從熒光圖像中獲得EdU陽性細胞數,計算EdU相對陽性率
2.6 對Balb/c雌性小鼠致病力的影響
為了探究缺陷43K OMP基因后,牛壞死桿菌對Balb/c雌性小鼠的致病力是否會發生改變,將兩菌株用無菌PBS稀釋后腹腔注射試驗組小鼠,每只接種5×108CFU細菌,對照組小鼠腹腔注射等量無菌PBS溶液,觀察小鼠死亡時間情況及全身病理變化情況。結果如圖5所示,親本株A25株和缺失株A25Δ43K OMP株對Balb/c雌性小鼠的致死率分別為100%和80%。感染親本株A25株的小鼠死亡時間主要集中在前3 d,感染缺失株A25Δ43K OMP株的小鼠主要集中在5~7 d(圖5A)。對小鼠無菌解剖發現,感染A25株小鼠臟器無明顯變化,感染A25Δ43K OMP株小鼠心、腎無明顯變化,肺充血,脾充血腫大,肝表面出現散在的米粒大小的黃白色壞死灶,部分出現黃白色大片干酪樣壞死并與腹膜黏連。對Balb/c雌性小鼠肝組織進行細菌載量計數發現,親本株A25株與缺失株A25Δ43K OMP株的肝細菌載量分別為2.29×105和1.72×105CFU·g-1,缺失株A25Δ43K OMP株定植在肝的細菌數量顯著低于親本株A25株(P<0.05,圖5B)。結果表明,缺陷43K OMP基因后,牛壞死桿菌對Balb/c雌性小鼠的致病力降低。

A. 各菌株對小鼠的存活曲線;B. 各菌株在小鼠肝組織的細菌載量。*. P<0.05
3 討 論
近年來,壞死桿菌感染引起的牛肝膿腫、腐蹄病、乳腺炎和子宮內膜炎等壞死桿菌疾病,對我國畜牧養殖業造成了嚴重的經濟損失,嚴重制約養牛業發展;壞死桿菌病不僅影響經濟動物還威脅到人類的生命安全,多地均報道過人類感染壞死桿菌的病例[19-21]。壞死桿菌如何感染動物機體并導致動物發病的機制研究尚未明確。
OMPs作為革蘭陰性細菌的重要組成部分,在細菌致病中發揮重要作用。43K OMP作為壞死桿菌的重要OMPs,前期研究發現其與細菌黏附功能有關。當壞死桿菌與43K OMP多抗孵育或43K OMP與細胞孵育后,壞死桿菌黏附宿主細胞能力降低[10];同時將43K OMP基因克隆到pFLAG-CTS載體中,隨后在大腸桿菌BL21 DE3表達后,大腸桿菌的黏附能力增強[22]。這些結果顯示,43K OMP在壞死桿菌黏附宿主細胞中具有重要作用,但壞死桿菌黏附是否與43K OMP基因具有直接作用尚不可知。因此,本研究對比43K OMP基因缺失株和牛壞死桿菌親本株對BEND細胞的黏附能力,發現缺陷43K OMP的牛壞死桿菌對BEND細胞的黏附能力顯著降低,這與之前的研究結果相一致。表明43K OMP基因直接介導壞死桿菌對細胞的黏附功能。
生物被膜是細菌抵抗外界不利環境的一種生存形式,主要由細菌、蛋白質、胞外多糖和胞外DNA構成,是細菌產生耐藥性和造成動物疾病持續感染的重要機制。Ye等[23]研究發現,阪崎氏菌(Cronobactersakazakii)缺失OmpW后,阪崎氏菌的生物被膜含量顯著增強。Yonezawa等[24]研究發現,缺失幽門螺桿菌的外膜蛋白AlpB導致幽門螺桿菌形成生物被膜含量顯著下降。牛壞死桿菌缺陷43K OMP基因后,其生物被膜形成能力顯著增強,顯示牛壞死桿菌43K OMP基因介導生物被膜的產生,而43K OMP基因引起的生物被膜形成能力的變化是與抵抗外界不利因素有關還是直接參與到生物被膜的形成還需進一步研究。
細菌的耐藥機制有很多種,如外排泵排出、細胞膜通透性改變、藥物靶點基因突變等,外排泵多由孔狀外膜蛋白構成,而外排泵系統也是介導細菌固有耐藥和獲得耐藥的重要機制[25]。Jeanteur等[26]將大腸桿菌的OmpF突變后,大腸桿菌對頭孢菌素的耐藥性降低;權衡等[27]敲除鴨疫里默氏桿菌的rant基因后,該菌對四環素的耐藥性降低。牛壞死桿菌43K OMP是由β-折疊平行排列構成的桶裝蛋白,其中包含9個跨膜區域,與梭菌屬的多種外膜成孔蛋白具有較高的相似性[9]。本研究中,壞死桿菌缺陷43K OMP基因導致其對部分抗菌藥物耐藥性發生變化,說明43K OMP基因參與到細菌的耐藥功能中,但其究竟是以外排泵的形式還是以改變細胞膜通透性或介導改變藥物靶基因突變從而導致耐藥性發生變化還需進一步研究。
4 結 論
牛壞死桿菌43K OMP基因被缺陷后,牛壞死桿菌的黏附能力顯著下降,生物被膜形成能力顯著提升,耐藥性產生變化,對小鼠致病力顯著下降。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
航空世界(2020年10期)2020-01-19 14:36:20
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
