基于轉(zhuǎn)錄組測序分析細(xì)粒棘球絳蟲原頭蚴與巨噬細(xì)胞RAW264.7免疫互作相關(guān)基因
2022-11-29 13:39:56王正榮張艷艷孟季蒙薄新文
畜牧獸醫(yī)學(xué)報(bào) 2022年11期
關(guān)鍵詞:差異
王正榮,馬 勛,張艷艷,孟季蒙,薄新文*
(1.省部共建綿羊遺傳改良與健康養(yǎng)殖國家重點(diǎn)實(shí)驗(yàn)室,石河子 832000;2.新疆農(nóng)墾科學(xué)院畜牧獸醫(yī)研究所,石河子 832000;3.石河子大學(xué)動(dòng)物科技學(xué)院,石河子 832000)
細(xì)粒棘球絳蟲(Echinococcusgranulosus)是囊型包蟲病的主要病原,該病是一種危害嚴(yán)重的人獸共患寄生蟲病,是一種被世界衛(wèi)生組織嚴(yán)重忽視的疾病之一。這種疾病與貧窮和不良的衛(wèi)生習(xí)慣有關(guān),特別是在畜牧業(yè)養(yǎng)殖地區(qū)[1]。同時(shí),包蟲病也是導(dǎo)致我國西部農(nóng)牧區(qū)牧民因病致貧、因病返貧的主要原因之一。目前,該病已在全國20個(gè)省份發(fā)生,主要分布在西北部分牧區(qū),包括青海、西藏、新疆等地。據(jù)估計(jì),我國包蟲病患者人數(shù)約為100萬,受威脅人口近6 600萬,患者數(shù)量巨大。與現(xiàn)有的全球包蟲病流行數(shù)據(jù)相比,中國包蟲病受威脅人口數(shù)和患者人數(shù)仍居世界首位[1-2]。然而,包蟲病病原與宿主的免疫互作的發(fā)病機(jī)制尚未明確,終末宿主疫苗的研究也相對滯后,這些都是影響該病徹底根除的主要因素和瓶頸問題。
巨噬細(xì)胞是一種重要的免疫細(xì)胞,是主要的抗原遞呈細(xì)胞。在機(jī)體先天免疫和適應(yīng)性免疫過程中具有重要的作用。現(xiàn)有的研究表明,巨噬細(xì)胞在宿主抗蠕蟲感染中具有重要的調(diào)控作用[3]。現(xiàn)已有很多有關(guān)巨噬細(xì)胞在抗棘球絳蟲感染中免疫功能研究的文獻(xiàn)報(bào)道,研究的內(nèi)容涉及巨噬細(xì)胞表達(dá)的一些功能分子[4-7]以及非編碼的miRNA分子[8-10]和lncRNA分子[11-12]等。同時(shí),也有一些研究棘球絳蟲功能分子在調(diào)節(jié)宿主免疫反應(yīng)方面的研究報(bào)道[13-15],但截至目前有關(guān)棘球絳蟲應(yīng)對宿主免疫反應(yīng)時(shí),反映蟲體整體基因差異表達(dá)的研究報(bào)道依然較少,致使人們對其缺乏系統(tǒng)的認(rèn)識。基于此,本研究采用轉(zhuǎn)錄組學(xué)技術(shù)分析了細(xì)粒棘球絳蟲原頭蚴應(yīng)對巨噬細(xì)胞免疫壓力時(shí),其基因的差異表達(dá)情況,以期系統(tǒng)解析其應(yīng)對宿主細(xì)胞免疫反應(yīng)時(shí)潛在的免疫調(diào)控機(jī)制,同時(shí)為疫苗和藥物的研發(fā)挖掘新的靶標(biāo)分子。
1 材料與方法
1.1 試驗(yàn)材料、試劑及主要儀器
在屠宰場收集被細(xì)粒棘球絳蟲感染的帶有包囊的羊肝,帶回實(shí)驗(yàn)室,無菌條件下抽取含有原頭蚴的包囊液,離心收集原頭蚴,用加有雙抗的PBS洗滌3~5次,最后將收集的原頭蚴置于含有10%胎牛血清以及雙抗的RPMI 1640培養(yǎng)液中,在37 ℃,5% CO2培養(yǎng)箱中培養(yǎng)備用。于細(xì)胞培養(yǎng)瓶中培養(yǎng)RAW 264.7細(xì)胞,待細(xì)胞生長穩(wěn)定時(shí)將其傳代培養(yǎng)至新的培養(yǎng)瓶,當(dāng)培養(yǎng)瓶中細(xì)胞交匯率達(dá)到80%時(shí),接種細(xì)粒棘球絳蟲原頭蚴,接種濃度為每毫升2 000個(gè),設(shè)PBS為對照組,每組3個(gè)生物學(xué)重復(fù),共培養(yǎng)24 h后,分別收集培養(yǎng)上清液中原頭蚴以及RAW264.7細(xì)胞,用PBS洗滌3次,最后將收集的原頭蚴保存于-80 ℃冰柜備用,建庫測序時(shí)采用混池測序。
胎牛血清購自Gibco公司(美國);RPMI 1640細(xì)胞培養(yǎng)液購自Sigma公司(美國);小鼠單核巨噬細(xì)胞RAW 264.7購自ATCC細(xì)胞庫(中國);SMARTer Ultra Low RNA Kit for Illumina Sequencing試劑盒購自Clontech公司(美國);AMPure XP beads試劑盒購自Beckman公司(美國);Invitrogen Trizol Reagent購自Invitrogen公司(美國);CW2582 cDNA合成試劑盒購自康為世紀(jì)生物技術(shù)有限公司(中國);SYBR PremixExTaqMix購自TaKaRa公司(日本);實(shí)時(shí)熒光定量PCR儀Lightcycle 2.0購自羅氏公司(德國)。
1.2 總RNA的提取、轉(zhuǎn)錄組文庫的建立以及測序
使用SMARTer Ultra Low RNA Kit for Illumina Sequencing試劑盒進(jìn)行細(xì)粒棘球絳蟲原頭蚴和RAW264.7細(xì)胞的裂解和第一條cDNA鏈的合成,使用AMPure XP beads試劑盒純化后,用Advantage 2 PCR kit試劑盒擴(kuò)增第一條cDNA鏈,并再次純化,最終得到雙鏈cDNA。Qubit 2.0定量檢測,合格后,用Covaris系統(tǒng)超聲打斷雙鏈cDNA短片段進(jìn)行末端修復(fù),加A尾并連接測序接頭后,純化,選擇片段大小在200 bp左右的文庫進(jìn)行PCR富集得到最終的cDNA文庫。先使用Qubit 2.0進(jìn)行文庫的初步定量,稀釋文庫至2 ng·μL-1,隨后用Agilent 2100對文庫的insert size進(jìn)行檢測,符合預(yù)期后,使用qPCR方法對文庫的有效濃度進(jìn)行準(zhǔn)確定量(文庫有效濃度>2 nmol·μL-1),以保證文庫質(zhì)量。文庫檢驗(yàn)合格后,把不同文庫按照有效濃度及目標(biāo)數(shù)據(jù)量的需求,進(jìn)行Hiseq測序。
1.3 差異基因的表達(dá)分析
使用Stringtie軟件對基因在轉(zhuǎn)錄水平上進(jìn)行表達(dá)量分析,皮爾遜相關(guān)系數(shù)表示樣品間基因的表達(dá)水平相關(guān)性,使用R語言的procmp函數(shù),根據(jù)表達(dá)量對各樣品進(jìn)行PCA主成分分析。采用DESeq對基因表達(dá)進(jìn)行差異分析,篩選條件為表達(dá)差異倍數(shù)|log2(Fold Change)|>1,顯著性Pvalue<0.05。
使用R語言ggplots 2軟件包繪制差異表達(dá)基因的火山圖,Pheatmap軟件包根據(jù)同一基因在不同樣品中的表達(dá)水平和同一樣品中不同基因的表達(dá)模式進(jìn)行聚類,采用Euclidan方法計(jì)算距離,使用層次聚類最長距離法進(jìn)行聚類。
1.4 差異表達(dá)基因的GO分析
利用GO(gene ontology)對差異表達(dá)基因進(jìn)行功能聚類分析。GO分為分子功能(molecular function)、生物過程(biological process)和細(xì)胞組成(cellular component)3個(gè)部分。差異表達(dá)的基因可以通過ID對應(yīng)或者序列注釋的方法找到與之對應(yīng)的GO編號,而GO編號可用于對應(yīng)到TERM,即功能類別或者細(xì)胞定位。如P-value≤0.05,則表示顯著性富集。
1.5 差異表達(dá)基因的KEGG分析
采用KEGG(kyoto encyclopedie of genes and genomes)數(shù)據(jù)庫對差異表達(dá)的基因進(jìn)行了信號通路分析,通過Pathway顯著性富集能確定特異基因參與的最主要生化代謝途徑和信號轉(zhuǎn)導(dǎo)途徑。具體方法以KEGG Pathway為單位,應(yīng)用超幾何檢驗(yàn),找出與整個(gè)基因組背景相比,在特異基因中顯著性富集的Pathway,最終將FDR≤0.05的Pathway定義為在差異表達(dá)基因中顯著富集的Pathway。
1.6 部分差異表達(dá)基因的轉(zhuǎn)錄水平分析
將無菌采集的細(xì)粒棘球絳蟲原頭蚴保存于液氮中,備用。總RNA提取采用Invitrogen Trizol Reagent試劑,提取方法參照試劑說明書步驟進(jìn)行。反轉(zhuǎn)錄采用康為世紀(jì)公司10 μL反轉(zhuǎn)錄體系的CW2582 cDNA合成試劑盒。
以上述cDNA為模板,以GAPDH為內(nèi)參基因,對隨機(jī)篩選的差異表達(dá)基因(引物見表1)進(jìn)行qRT-PCR擴(kuò)增,反應(yīng)體系:SYBR PremixExTaqMix(2×) 10 μL,上下游引物(10 mmol·L-1)各0.4 μL,cDNA 1 μL,加ddH2O 至20 μL。反應(yīng)條件:95 ℃預(yù)變性30 s;95 ℃ 5 s,60 ℃ 20 s,共40個(gè)循環(huán)。每個(gè)基因做3個(gè)生物學(xué)重復(fù),采用相對比較ΔCt(Qr=2-ΔΔCt)法計(jì)算目的基因的相對表達(dá)量。

表1 實(shí)時(shí)定量PCR試驗(yàn)相關(guān)引物
2 結(jié) 果
2.1 細(xì)粒棘球絳蟲原頭蚴差異基因表達(dá)分析
當(dāng)細(xì)粒棘球絳蟲原頭蚴與巨噬細(xì)胞RAW264.7共培養(yǎng)24 h后,與0 h相比,共培養(yǎng)24 h后的原頭蚴共有435個(gè)基因的表達(dá)出現(xiàn)顯著變化,其中,227個(gè)基因呈上調(diào)表達(dá),208個(gè)基因呈下調(diào)表達(dá)。對差異表達(dá)基因進(jìn)行火山圖以及前20位(top20)聚類熱圖可視化分析,本研究將-lg(FDR)>1的差異表達(dá)定義為顯著差異表達(dá)基因(圖1)。
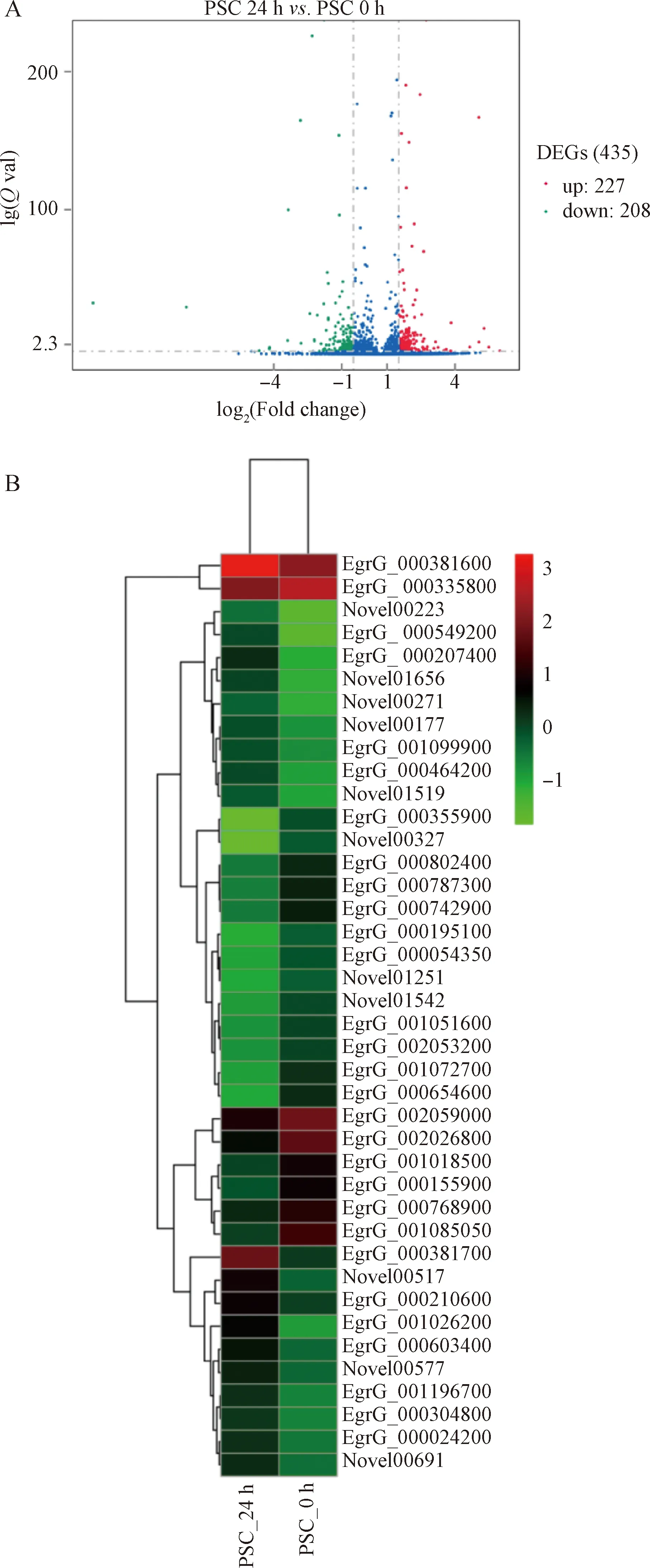
圖1 原頭蚴與RAW264.7共培養(yǎng)24 h后差異表達(dá)基因的火山圖(A)、top20熱圖(B)分析
其中,上調(diào)表達(dá)前10的基因包括Novel00223、EgrG_000549200、EgrG_001026200(NHRF3)、EgrG_000207400、EgrG_000381700、Novel01656、Novel00517、Novel00271(EGR_00497)、EgrG_000464200和Novel01519(EG95 protein precursor)。下調(diào)表達(dá)前10的基因包括EgrG_000155900(ALDR)、EgrG_001072700(DPYD)、EgrG_002026800(SYNC)、EgrG_001085050(HSP70)、EgrG_000654600(CATL)、EgrG_000355900、Novel00327(EF-hand domain-containing protein)、EgrG_002320300、pathogen_EgG_scaffold_0325_ncRNA_4499-4601(LORF2)和EgrG_002059100(表2)。
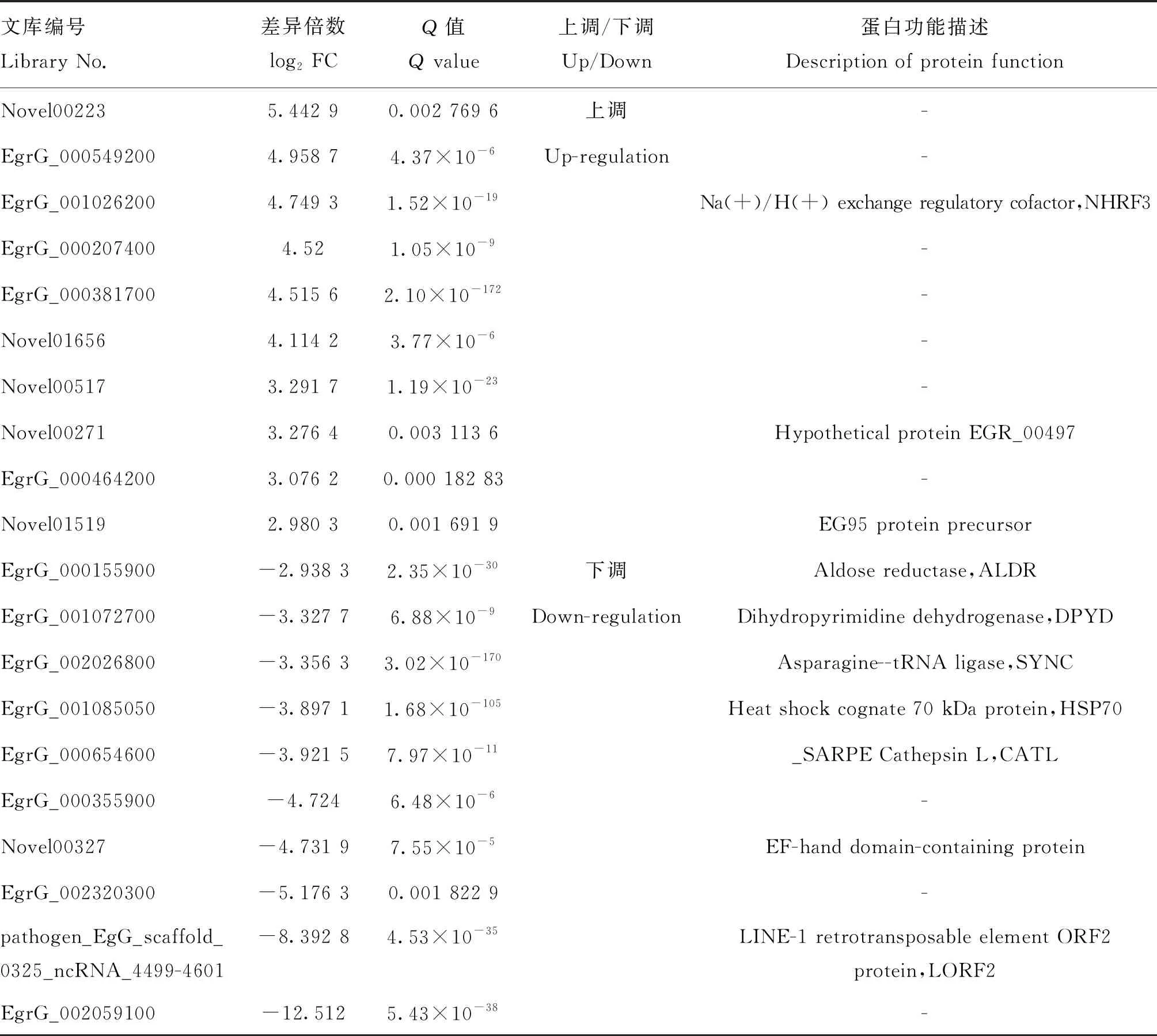
表2 感染后24 h Top10差異表達(dá)基因
此外,在上調(diào)表達(dá)的基因中還有EgrG_000320800(HSP10)、EgrG_000515900(AgB)、EgrG_000550000(FABP1)、EgrG_000253300(囊泡運(yùn)輸?shù)鞍譙C22B)、EgrG_000460300(鈣結(jié)合蛋白SM16)和EgrG_000715700(PDCD6)等。下調(diào)表達(dá)的基因中還包括EgrG_001018500(跨膜蛋白TM144)、EgrG_000802400(內(nèi)固醇類受體)、Novel01542(MAPK7)、EgrG_000317500(丙氨酸氨基轉(zhuǎn)移酶2,ALAT2)、EgrG_000589100(谷氨酸脫氫酶2,GLUD2)和EgrG_000644000(谷氨酸合酶,NADH)等。
2.2 細(xì)粒棘球絳蟲原頭蚴差異表達(dá)基因的GO分析
GO分析的結(jié)果顯示,當(dāng)細(xì)粒棘球絳蟲原頭蚴與巨噬細(xì)胞RAW264.7共培養(yǎng)24 h后,其差異表達(dá)基因的功能主要富集在生物學(xué)過程(BP)的血紅素轉(zhuǎn)運(yùn)、鐵配位實(shí)體運(yùn)輸、輔因子輸送、α-氨基酸生物合成過程以及細(xì)胞形態(tài)發(fā)生等。細(xì)胞組分(CC)的細(xì)胞外間質(zhì)、核糖核酸酶MRP復(fù)合物、核糖核酸酶P復(fù)合物以及胞內(nèi)核糖核酸復(fù)合體等。分子功能(MF)的絲氨酸型內(nèi)肽酶抑制劑活性、α-半乳糖苷酶活性、輔因子結(jié)合以及磷脂酰肌醇乙酰氨基葡萄糖轉(zhuǎn)移酶活性等過程(圖2,表3)。
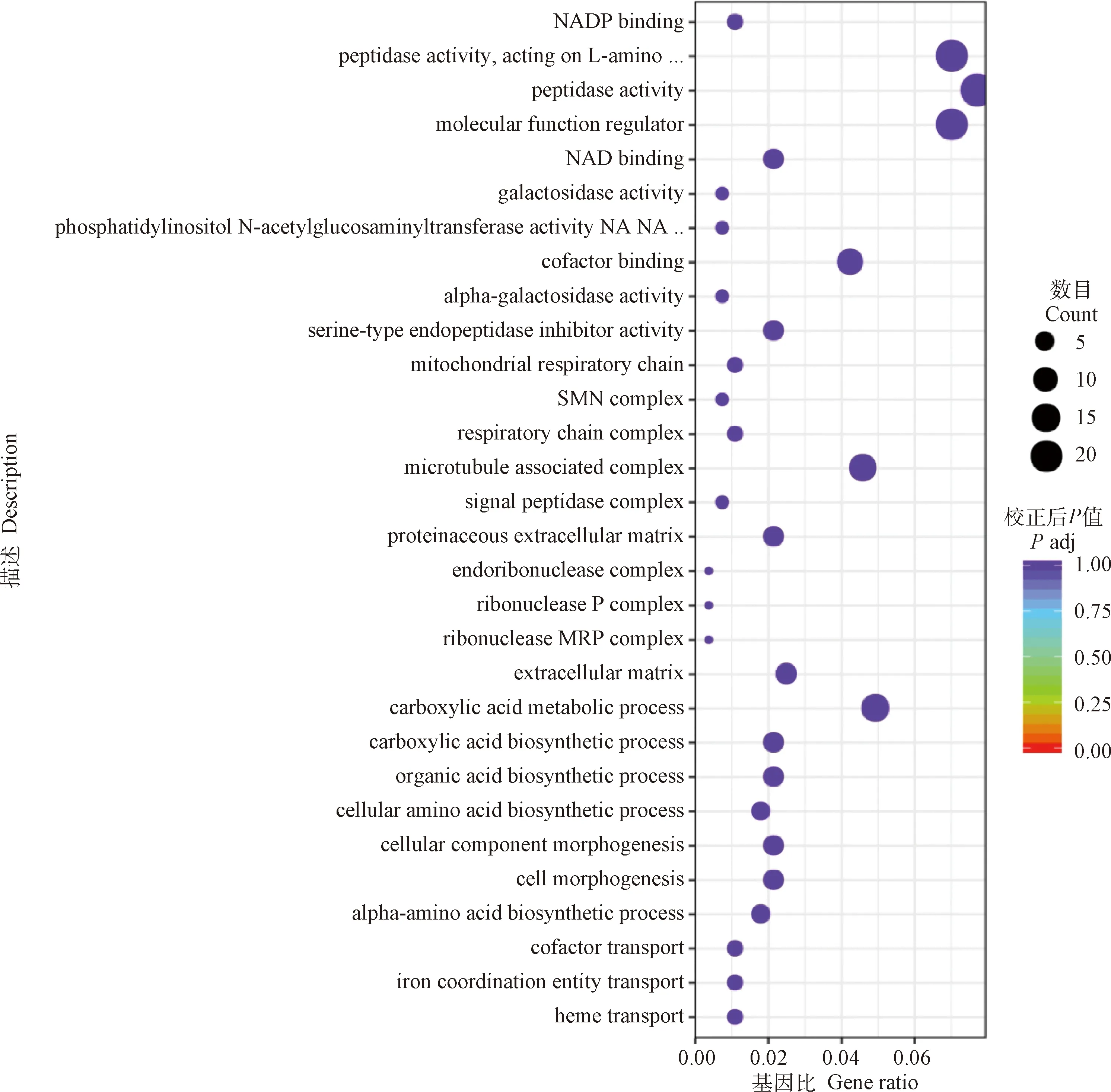
圖2 與RAW264.7共培養(yǎng)24 h后原頭蚴中差異表達(dá)基因的GO分析
2.3 細(xì)粒棘球絳蟲原頭蚴差異表達(dá)基因的KEGG分析
KEGG分析的結(jié)果顯示,當(dāng)細(xì)粒棘球絳蟲原頭蚴與巨噬細(xì)胞RAW264.7共培養(yǎng)24 h后,原頭蚴中差異表達(dá)的基因主要參與剪接體、內(nèi)吞作用、內(nèi)質(zhì)網(wǎng)的蛋白質(zhì)處理、吞噬體、MAPK信號通路以及鈣信號通路等。其中,上調(diào)表達(dá)的基因主要參與剪接體、內(nèi)質(zhì)網(wǎng)的蛋白質(zhì)處理、核糖體、吞噬體以及MAPK等信號通路。下調(diào)表達(dá)的基因主要參與丙氨酸、天冬氨酸和谷氨酸代謝、鈣信號通路、氨基酸生物合成以及內(nèi)吞作用等信號通路(圖3,表4)。
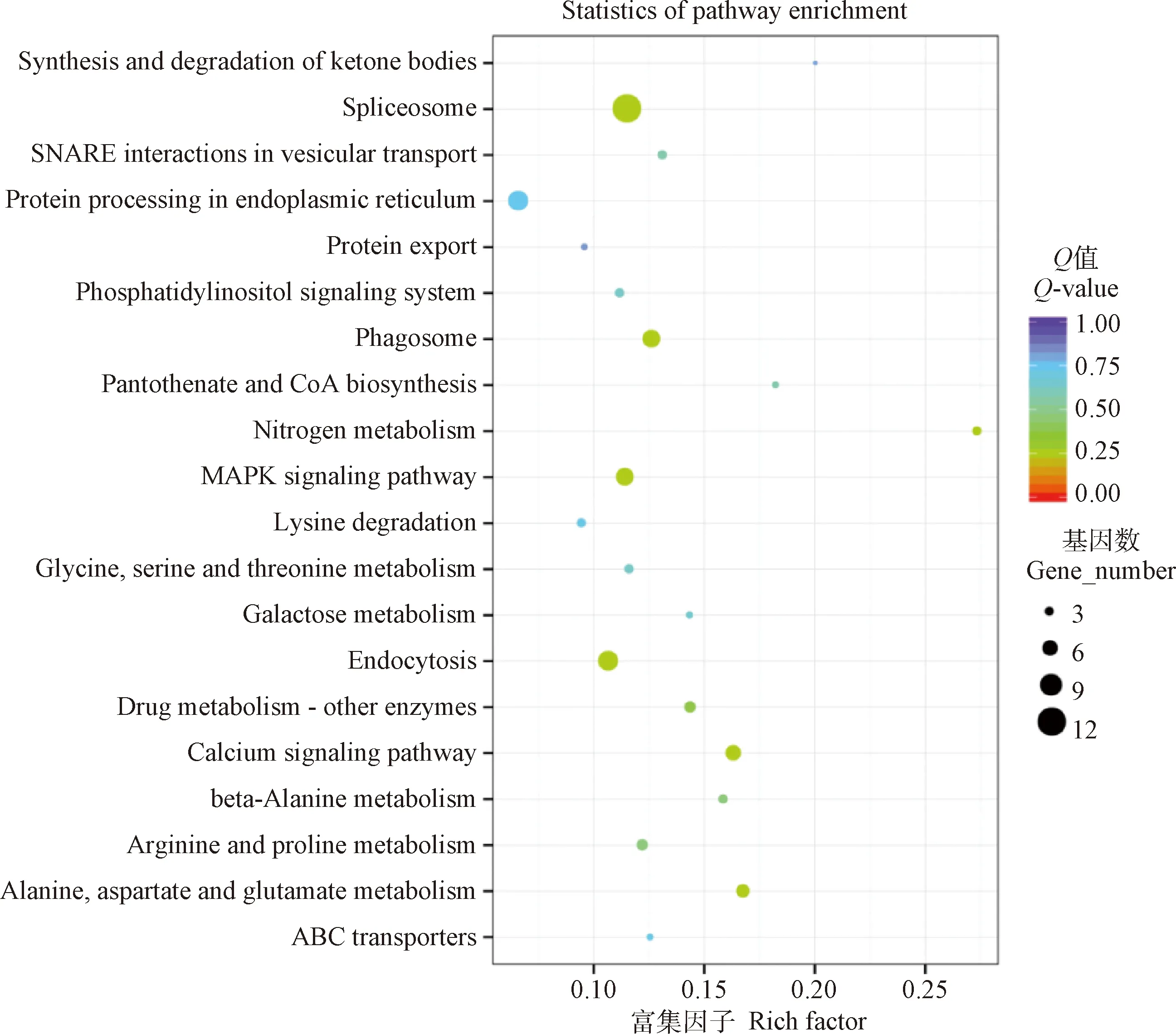
圖3 與RAW264.7共培養(yǎng)24 h后原頭蚴中差異表達(dá)基因的KEGG分析
2.4 原頭蚴差異表達(dá)基因中各種酶相關(guān)基因的表達(dá)譜
分析發(fā)現(xiàn),在435個(gè)差異表達(dá)的基因中,有79個(gè)各種酶相關(guān)的基因,其中,上調(diào)表達(dá)的26個(gè),主要包括乙酰氨基葡萄糖轉(zhuǎn)移酶、神經(jīng)鞘脂脫飽和酶、特異性二酰甘油脂肪酶、線粒體輸入性內(nèi)膜轉(zhuǎn)位酶、谷氨酸脫氫酶、谷氨酸半胱氨酸連接酶、胞苷脫氨酶、L-天冬酰胺酶、組蛋白賴氨酸甲基轉(zhuǎn)移酶、蛋白質(zhì)糖基轉(zhuǎn)移酶、信號肽酶、甘露糖-1-磷酸鳥苷轉(zhuǎn)移酶、甘油激酶、脫氧胞嘧啶合酶、葡萄糖胺6-磷酸N-乙酰基轉(zhuǎn)移酶、精氨酸轉(zhuǎn)氨酶、蛋白轉(zhuǎn)移酶、甲戊二酸單酰輔酶A合酶、磷脂促翻轉(zhuǎn)酶以及亞精胺合成酶。同時(shí)還包括3個(gè)抑制劑相關(guān)的基因,分別為Kunitz型絲氨酸蛋白酶抑制劑6、Kunitz型絲氨酸蛋白酶抑制劑Bt-KTI以及蛋白酶抑制劑I2(表3)。下調(diào)表達(dá)有53個(gè),主要包括天冬酰胺-tRNA連接酶、二氫嘧啶脫氫酶、醛糖還原酶、腺苷酸琥珀酸裂解酶、谷氨酸脫氫酶2、谷氨酸合酶(NADH)、鳥氨酸轉(zhuǎn)氨酶、L-蘇氨酸3-脫氫酶、L-乳酸脫氫酶A、尿苷磷酸化酶1等,圖4顯示了各種酶相關(guān)的基因差異表達(dá)top20的分析結(jié)果。

圖4 與RAW264.7共培養(yǎng)24 h后原頭蚴中差異表達(dá)酶相關(guān)基因top20的表達(dá)譜分析
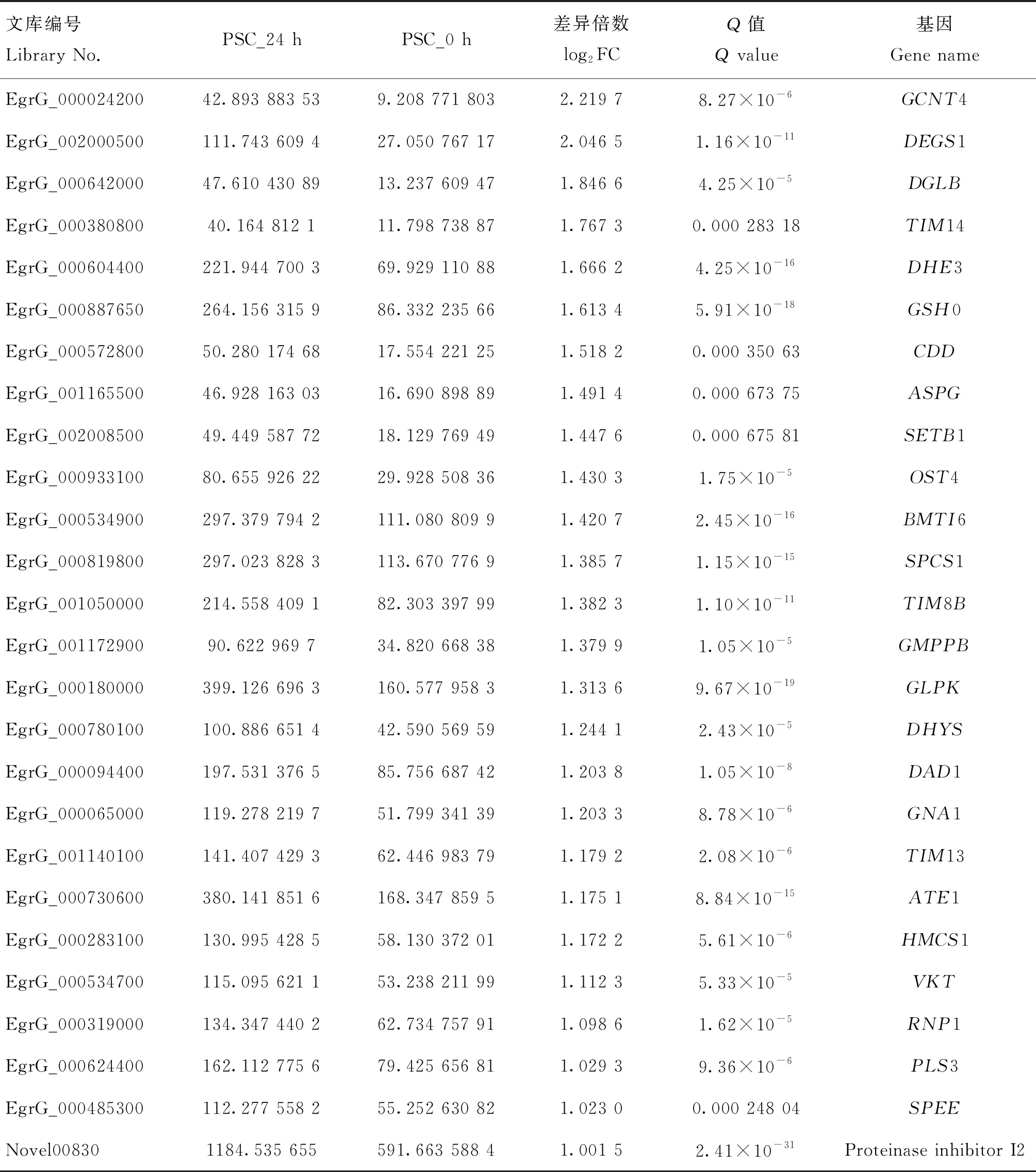
表3 與巨噬細(xì)胞RAW264.7共培養(yǎng)24 h后原頭蚴中上調(diào)表達(dá)的各種酶相關(guān)基因
2.5 巨噬細(xì)胞RAW264.7基因差異表達(dá)分析
當(dāng)巨噬細(xì)胞RAW264.7與細(xì)粒棘球絳蟲原頭蚴共培養(yǎng)24 h時(shí),和PBS對照組相比,原頭蚴處理組的RAW264.7細(xì)胞共有3 745個(gè)基因的表達(dá)出現(xiàn)顯著變化,其中,1 159個(gè)基因出現(xiàn)上調(diào)表達(dá),2 586個(gè)基因出現(xiàn)下調(diào)表達(dá)(圖5)。
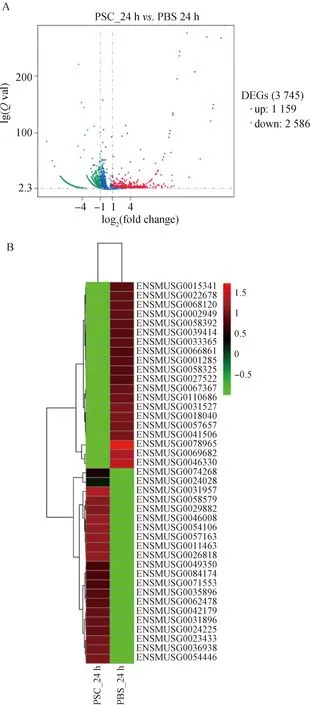
圖5 RAW264.7與原頭蚴共培養(yǎng)24 h后差異表達(dá)基因的火山圖(A)、top20熱圖(B)分析
其中,表達(dá)上調(diào)前10的基因包括ENSMUSG0011463(Cpb1)、ENSMUSG0058579(Cela2a)、ENSMUSG0029882(2210010C04Rik)、ENSMUSG0036938(Try5)、ENSMUSG0035896 (Rnase1)、ENSMUSG0062478(Ctrc)、ENSMUSG0084174(Sycn)、ENSMUSG0031896(Ctrl)、ENSMUSG0031957(Ctrb1)和ENSMUSG0074268(Amy2a5);下調(diào)表達(dá)前10的基因包括ENSMUSG0022678(Nde1)、ENSMUSG0067367(Lyar)、ENSMUSG00110686、ENSMUSG0057657、ENSMUSG0041506(Rrp9)、ENSMUSG0031527(Eri1)、ENSMUSG0018040(Rrp7a)、ENSMUSG0069682、ENSMUSG0046330(Rpl37a)和ENSMUSG0078965(表4)。
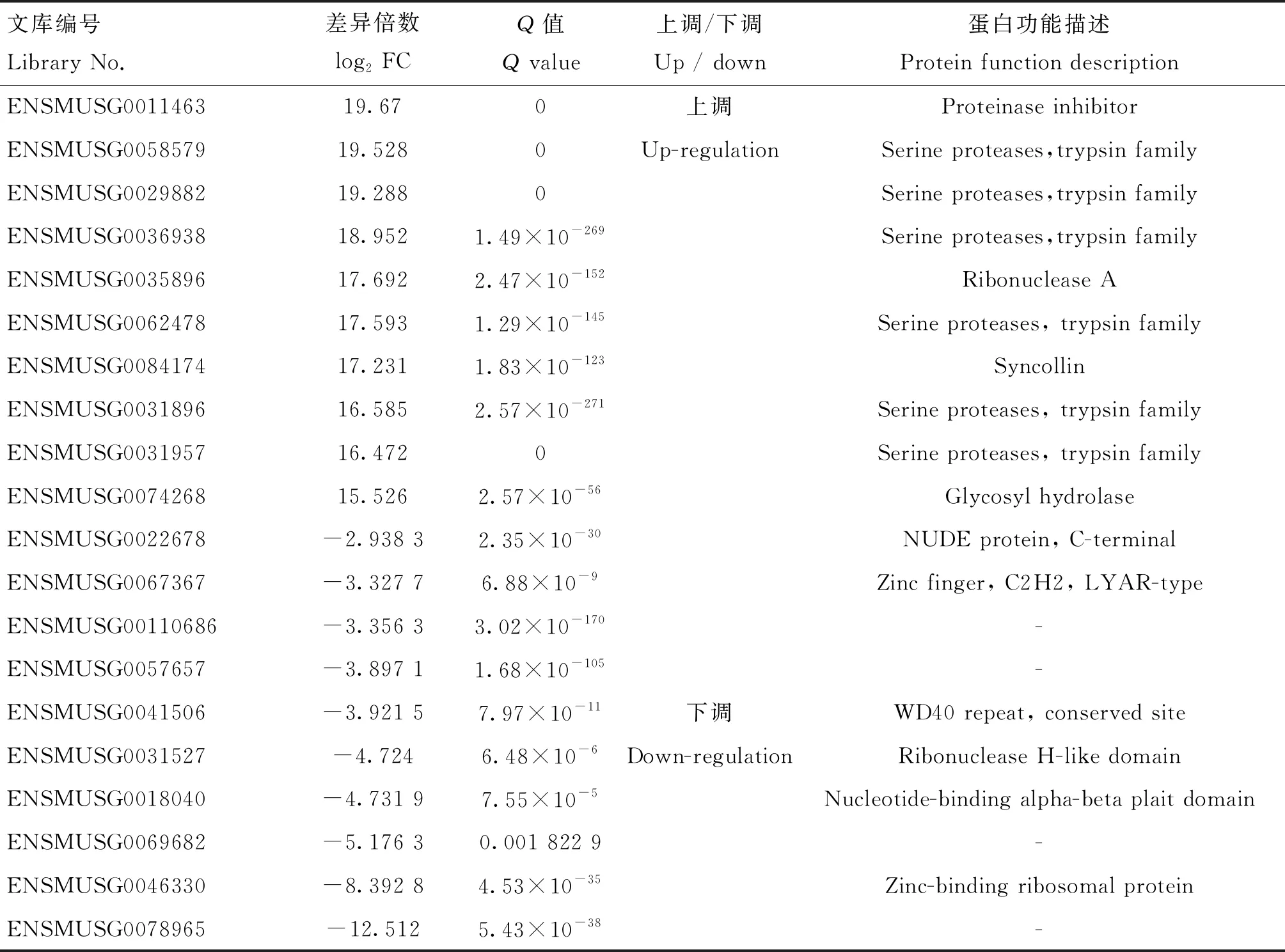
表4 共培養(yǎng)24 h后 RAW264.7 Top10差異表達(dá)基因
2.6 巨噬細(xì)胞RAW264.7差異基因表達(dá)的GO分析
當(dāng)巨噬細(xì)胞RAW264.7與細(xì)粒棘球絳蟲原頭蚴共培養(yǎng)24 h時(shí),與PBS對照組相比,GO分析結(jié)果顯示,差異表達(dá)基因主要富集在GO功能聚類的生物過程(BP)中的代謝過程、初級代謝過程、有機(jī)物代謝過程以及細(xì)胞代謝過程等。細(xì)胞組分(CC)中的細(xì)胞內(nèi)組分、細(xì)胞內(nèi)、細(xì)胞器以及膜結(jié)構(gòu)細(xì)胞器等(圖6)。

圖6 與原頭蚴共培養(yǎng)24 h后RAW264.7中差異表達(dá)基因的GO分析
2.7 巨噬細(xì)胞RAW264.7差異基因表達(dá)的KEGG分析
當(dāng)巨噬細(xì)胞RAW264.7與細(xì)粒棘球絳蟲原頭蚴共培養(yǎng)24 h時(shí),與PBS對照組相比,KEGG信號通路的分析結(jié)果表明,差異表達(dá)的基因主要參與代謝通路、核糖體通路、剪接體通路、RNA轉(zhuǎn)運(yùn)以及泛素介導(dǎo)的蛋白水解等通路(圖7)。
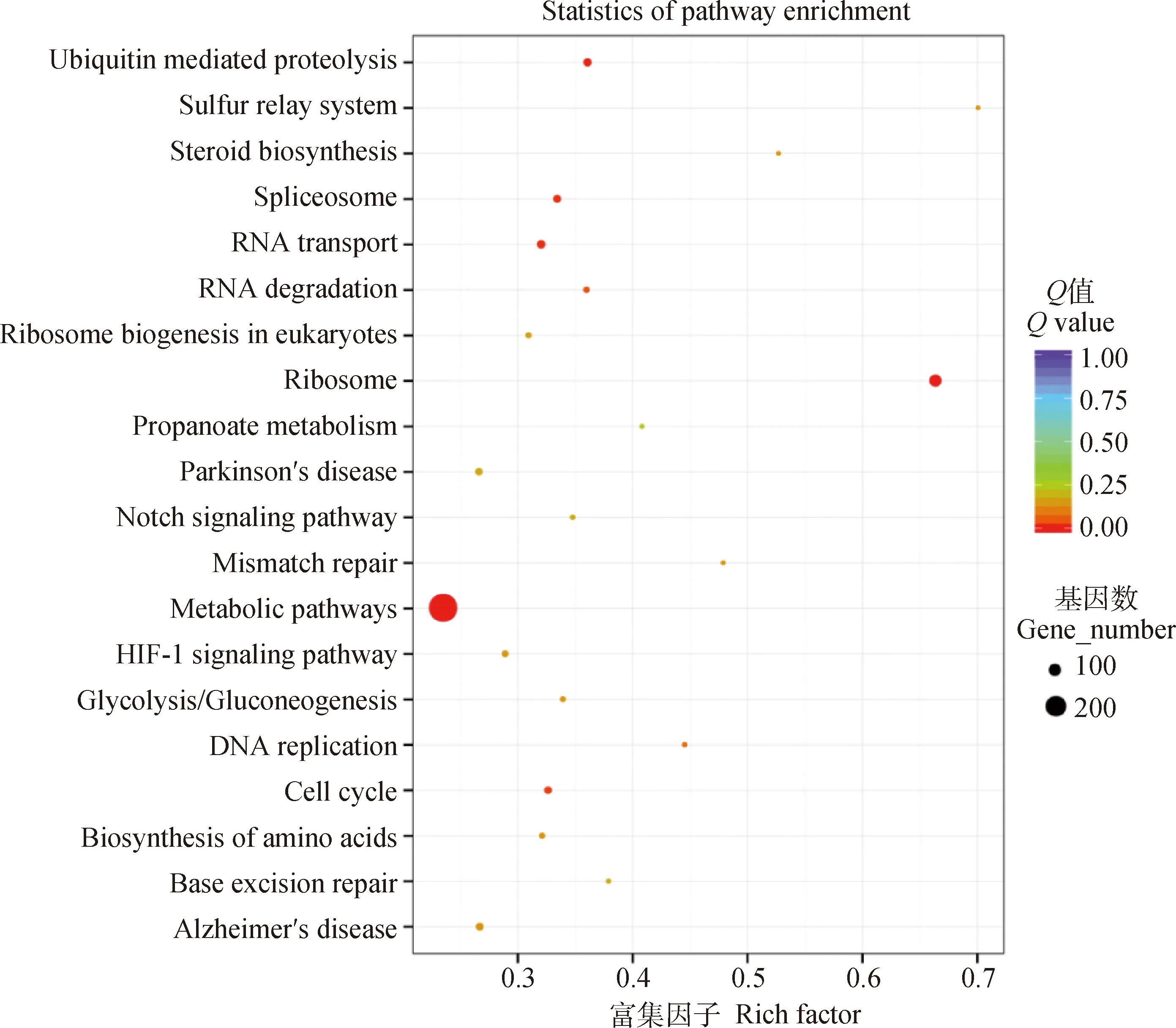
圖7 與原頭蚴共培養(yǎng)24 h后RAW264.7中差異表達(dá)基因的KEGG分析
2.8 差異表達(dá)基因的qRT-PCR驗(yàn)證
為了驗(yàn)證從轉(zhuǎn)錄組測序數(shù)據(jù)中獲得差異基因結(jié)果的可靠性,隨機(jī)選取10個(gè)差異表達(dá)的基因進(jìn)行qRT-PCR驗(yàn)證,其中,包括6個(gè)上調(diào)表達(dá)基因,以及4個(gè)下調(diào)表達(dá)基因,結(jié)果顯示,qRT-PCR驗(yàn)證結(jié)果與RNA-seq測序結(jié)果一致,進(jìn)一步說明轉(zhuǎn)錄組測序結(jié)果可信(圖8)。
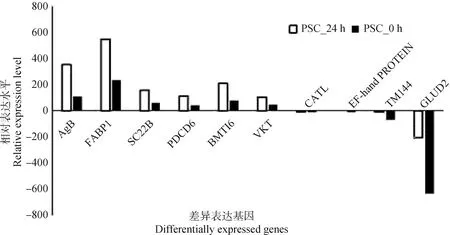
圖8 差異表達(dá)基因的qRT-PCR驗(yàn)證分析
3 討 論
原頭蚴是細(xì)粒棘球絳蟲的重要發(fā)育階段,具有雙向發(fā)育的特征,即在中間宿主體內(nèi)可以經(jīng)無性繁殖發(fā)育為新的包囊,在終末宿主體內(nèi)可以經(jīng)有性繁殖發(fā)育為成蟲[16]。原頭蚴也是中間宿主形成二次感染的重要原因,同時(shí)還是細(xì)粒棘球絳蟲感染終末宿主,在其腸道定植發(fā)育為成蟲的最初形態(tài)。可以看出,原頭蚴不僅是細(xì)粒棘球絳蟲生長發(fā)育的關(guān)鍵環(huán)節(jié),同時(shí)也是感染宿主的重要階段。因此研究細(xì)粒棘球絳蟲原頭蚴生長發(fā)育關(guān)鍵分子以及原頭蚴與終末宿主免疫互作關(guān)鍵分子均具有重要意義,研究結(jié)果將不僅揭示其獨(dú)特的發(fā)育現(xiàn)象,同時(shí)還可能挖掘出潛在疫苗候選靶標(biāo),進(jìn)而為疫苗的研發(fā)奠定基礎(chǔ)。基于以上原因,本研究通過轉(zhuǎn)錄組學(xué)技術(shù)解析了細(xì)粒棘球絳蟲原頭蚴與重要免疫細(xì)胞-巨噬細(xì)胞的免疫互作關(guān)鍵分子,以期初步闡明其免疫互作機(jī)制,同時(shí)發(fā)掘疫苗候選抗原。
本研究的結(jié)果表明,當(dāng)細(xì)粒棘球絳蟲原頭蚴與巨噬細(xì)胞RAW264.7共培養(yǎng)24 h后,和0 h相比,共培養(yǎng)24 h后的原頭蚴共有435個(gè)基因的表達(dá)出現(xiàn)顯著變化,其中227個(gè)基因出現(xiàn)上調(diào)表達(dá),208個(gè)基因出現(xiàn)下調(diào)表達(dá)。由此可見,當(dāng)原頭蚴在面對巨噬細(xì)胞RAW264.7免疫壓力時(shí),其自身基因表達(dá)會發(fā)生變化,包括與抗逆反應(yīng)相關(guān)的熱激蛋白HSP70、HSP10,與免疫調(diào)節(jié)相關(guān)的AgB、FABP1和Kunitz型絲氨酸蛋白酶抑制劑,還有與胞外囊泡運(yùn)輸相關(guān)的蛋白SC22B,這些基因的表達(dá)都出現(xiàn)顯著上調(diào),由此推測其可能在原頭蚴抗巨噬細(xì)胞RAW264.7免疫壓力過程中起到重要的調(diào)控作用,但其具體的調(diào)控機(jī)制還有待進(jìn)一步研究。反觀巨噬細(xì)胞RAW264.7的基因表達(dá)情況,當(dāng)巨噬細(xì)胞RAW264.7與細(xì)粒棘球絳蟲原頭蚴共培養(yǎng)24 h后,和PBS對照組相比,原頭蚴處理組的RAW264.7細(xì)胞共有3 745個(gè)基因的表達(dá)出現(xiàn)顯著變化,其中,1 159個(gè)基因出現(xiàn)上調(diào)表達(dá),2 586個(gè)基因出現(xiàn)下調(diào)表達(dá)。在這些差異表達(dá)的基因中,作者發(fā)現(xiàn)大量與天然免疫相關(guān)的基因的表達(dá)也發(fā)生了變化,諸如C型凝集素受體Reg1、Reg2、Reg3b、Reg3d、Reg3a和Reg3g的表達(dá)出現(xiàn)上調(diào)。病原模式識別受體下游相關(guān)效應(yīng)分子Fam132a、Cbln2、和Tnfrsf12a的表達(dá)也出現(xiàn)上調(diào),其中,F(xiàn)am132a和Cbln2屬于腫瘤壞死因子樣的結(jié)構(gòu)域,而Tnfrsf12a屬于腫瘤壞死因子受體12,都屬于炎型細(xì)胞因子調(diào)節(jié)因子。并且在表達(dá)上調(diào)前10的分子中有6個(gè)屬于絲氨酸蛋白酶家族,包括Cela2a、2210010C04Rik、Try5、Ctrc、Ctrl和Ctrb1[7]。提示這些基因可能在巨噬細(xì)胞RAW264.7抗原頭蚴感染過程中發(fā)揮重要功能。
在這些差異表達(dá)的基因中,AgB基因的表達(dá)出現(xiàn)上調(diào),AgB是棘球絳蟲中研究最為廣泛的抗原之一[17],其在細(xì)粒棘球絳蟲包囊液中普遍存在,是目前人類包蟲病診斷的主要抗原之一。已有的研究證實(shí),細(xì)粒棘球絳蟲分泌的AgB可以通過中性粒細(xì)胞分泌的彈性蛋白酶干擾中性粒細(xì)胞的活性,進(jìn)而有利于寄生蟲逃避宿主的免疫反應(yīng)。另外,AgB可以通過調(diào)節(jié)巨噬細(xì)胞的活性和抑制有效細(xì)胞因子的產(chǎn)生來調(diào)節(jié)宿主的免疫系統(tǒng)[18]。此外,本研究還發(fā)現(xiàn)FABP1基因的表達(dá)同樣出現(xiàn)上調(diào)。現(xiàn)有的研究結(jié)果表明,F(xiàn)ABP對于寄生的扁形動(dòng)物具有重要的意義,因?yàn)檫@些寄生蟲不能合成它們自身所需的大部分脂質(zhì),特別是長鏈脂肪酸和膽固醇[19-20]。因此,這種脂質(zhì)必須從其寄生宿主體內(nèi)獲得,然后通過載體蛋白質(zhì)運(yùn)送到寄生蟲體內(nèi)的特定部位。FABP蛋白可能參與了其細(xì)胞外脂質(zhì)的獲取,以及傳遞到宿主細(xì)胞的過程。同時(shí)FABP蛋白家族也是極具潛力的藥物傳遞以及疫苗候選靶標(biāo)[21-24]。研究者在棘球蚴囊液和原頭節(jié)分泌物中發(fā)現(xiàn)了FABP1分子[24-25]。細(xì)粒棘球絳蟲的FABP1被認(rèn)為是其亞家族的重要成員之一[26],其成員參與脂質(zhì)氧化過程。研究表明,細(xì)粒棘球絳蟲的FABP1通過一種機(jī)制與膜相互作用,這種機(jī)制涉及到與膜直接接觸以它們的物質(zhì)交換[27]。由此推斷,以上分子可能在細(xì)粒棘球絳蟲應(yīng)對巨噬細(xì)胞免疫反應(yīng)時(shí)起到重要免疫調(diào)控作用,以創(chuàng)造有利于蟲體寄生的免疫微環(huán)境。
蟲體表達(dá)的各種酶相關(guān)的蛋白以及抑制劑,不僅在蟲體自身的代謝中起重要的作用,同時(shí)可以降解對其有害的宿主蛋白以及抑制宿主來源的各種酶,保護(hù)蟲體自身不受危害[28]。為此,本研究系統(tǒng)分析了細(xì)粒棘球絳蟲原頭蚴在應(yīng)對巨噬細(xì)胞免疫壓力時(shí),其各種酶相關(guān)基因以及抑制劑相關(guān)分子的表達(dá)情況。結(jié)果發(fā)現(xiàn),有79個(gè)酶相關(guān)的基因其表達(dá)出現(xiàn)顯著差異,其中,包括3個(gè)上調(diào)表達(dá)的蛋白酶抑制劑,分別為Kunitz型絲氨酸蛋白酶抑制劑6、Kunitz型絲氨酸蛋白酶抑制劑Bt-KTI以及蛋白酶抑制劑I2。有學(xué)者在細(xì)粒棘球絳蟲中分離鑒定了2個(gè)Kunitz型蛋白酶抑制劑,其中,一種為Kunitz型絲氨酸蛋白酶抑制劑,是一種強(qiáng)有力的糜蛋白酶和中性粒細(xì)胞彈性蛋白酶抑制劑,在局部炎癥中結(jié)合鈣以及減少中性粒細(xì)胞浸潤。同時(shí)也可能通過抑制中性粒細(xì)胞彈性蛋白酶和組織蛋白酶G,參與宿主免疫逃避。而另一個(gè)是一種強(qiáng)有力的胰蛋白酶抑制劑。作為哺乳動(dòng)物腸道蛋白酶的強(qiáng)有力抑制劑,細(xì)粒棘球絳蟲Kunitz型蛋白可能在防止蛋白水解酶攻擊方面發(fā)揮重要的保護(hù)作用,從而確保細(xì)粒棘球絳蟲在其哺乳動(dòng)物宿主體內(nèi)的存活[29]。近期,有研究發(fā)現(xiàn),細(xì)粒棘球絳蟲的Kunitz型蛋白酶抑制劑可以抑制巨噬細(xì)胞炎型細(xì)胞因子的產(chǎn)生,并且抑制巨噬細(xì)胞的增殖[15, 30]。肝片吸蟲的排泄-分泌產(chǎn)物中存在兩種Kunitz型絲氨酸蛋白酶抑制劑,這些抑制劑可以保護(hù)寄生蟲在入侵時(shí)免受宿主蛋白酶,如糜蛋白酶等有害蛋白的水解作用。同時(shí)研究還發(fā)現(xiàn)此類絲氨酸蛋白酶抑制劑可以通過干擾激肽釋放酶-激肽系統(tǒng),進(jìn)而調(diào)節(jié)宿主的炎癥和通透性反應(yīng)[31]。由此作者推測,這3種蛋白酶抑制劑可能在細(xì)粒棘球絳蟲應(yīng)對巨噬細(xì)胞免疫反應(yīng)時(shí)起重要的作用,一方面,抑制宿主細(xì)胞釋放的蛋白酶活性,保護(hù)蟲體在寄生過程中免受宿主蛋白酶水解作用;另一方面,抑制宿主免疫細(xì)胞分泌炎性細(xì)胞因子,調(diào)節(jié)宿主免疫反應(yīng),達(dá)到寄生的目的。
4 結(jié) 論
本研究系統(tǒng)解析了細(xì)粒棘球絳蟲原頭蚴應(yīng)對巨噬細(xì)胞RAW264.7免疫壓力時(shí)其基因的差異表達(dá)譜規(guī)律及其巨噬細(xì)胞RAW264.7面對蟲體抗原刺激時(shí)其免疫相關(guān)基因的表達(dá)情況,篩選了與細(xì)粒棘球絳蟲抗宿主免疫反應(yīng)相關(guān)分子。研究結(jié)果顯示,當(dāng)細(xì)粒棘球絳蟲原頭蚴面對宿主巨噬細(xì)胞免疫壓力時(shí),原頭蚴中的AgB、FABP1、Kunitz型絲氨酸蛋白酶抑制劑等具有免疫調(diào)節(jié)作用的分子表達(dá)明顯上調(diào),進(jìn)而有利于蟲體在宿主的寄生以及免疫逃避。這些數(shù)據(jù)將為進(jìn)一步揭示細(xì)粒棘球絳蟲抗宿主免疫反應(yīng)以適應(yīng)寄生的機(jī)理提供理論依據(jù),同時(shí)為包蟲病防治提供新的疫苗和藥物靶點(diǎn)奠定基礎(chǔ)。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實(shí)用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50
