骨髓源肥大細胞通過甘露糖受體識別FMDV-VLPs的細胞因子應答
2022-11-29 13:20:08韓偉建張俊娟張義明王家鑫李麗敏
畜牧獸醫學報 2022年11期
韓偉建,張俊娟,張義明,王家鑫,李麗敏
(河北農業大學動物醫學院,保定 071000)
肥大細胞(mast cells,MCs)廣泛分布于皮膚與黏膜等部位的結締組織中,與樹突狀細胞和巨噬細胞共同構成機體抵御病原體的第一道防線,并構成固有免疫和適應性免疫應答之間的橋梁[1]。MCs可以快速識別入侵的病原體并協助啟動免疫應答。一旦活化, MCs可以迅速釋放大量的免疫介質,包括細胞因子、趨化因子、蛋白酶和抗菌肽[2],例如干擾素、TNF-α、IL-1α、IL-1β、IL-4、IL-6、IL-8、IL-10、IL-17、CCL2、CCL3、CCL5等[3]。這些介質可以激活免疫和非免疫細胞,并對其周圍的細胞功能進行調節[4]。
MCs分泌細胞因子需要其表面的識別受體識別相應的配體。MCs可通過多種模式識別受體(pattern recognition receptors, PRRs)識別病原體,包括TLR樣受體(toll-like receptors, TLRs)[5]、甘露糖受體(mannose receptors, MR)、NOD樣受體(nod-like receptors, NLR)、清道夫受體(scavenger receptor, SR)和CD48等受體[6]。MCs可以通過TLRs和RIG-I受體識別病原體,分泌細胞因子發揮調節作用及抗病毒作用[7-8]。MR是C型凝集素受體家族重要成員之一,不僅可以識別病原體表面的甘露糖、巖藻糖等,還可以識別病毒的結構蛋白[9],從而誘導機體產生抗微生物免疫應答。有研究表明,MCs可以通過MR識別百日咳桿菌并分泌TNF-α、IL-6和IFN-γ[10]。MCs分泌的TNF-α 在細菌感染過程中可以招募DC及T細胞至引流淋巴結,從而促進抗原特異性免疫應答[4,11],而MCs分泌的IL-6以及MCs與B細胞的相互作用是機體分泌IgA的主要機制[12]。那么,MCs通過MR識別FMDV-VLPs 分泌的細胞因子表達情況如何?
口蹄疫病毒樣顆粒(foot-and-mouth disease virus-like particles, FMDV-VLPs)目前已成為FMD的主要候選疫苗之一[13]。在FMDV衣殼的4個結構蛋白中,VP1可以刺激機體產生中和性抗體[14],而VP4是一種高度保守的結構蛋白,雖然位于病毒粒子的內部,但是在病毒的侵入過程中起了重要的作用[15],牛的多種單體型MHC分子都可以識別VP4蛋白的20-34位氨基酸序列[16]。因此,VP1和VP4被認為是研制通用型口蹄疫疫苗的理想后備抗原[17]。本實驗室將VP1-VP4連接在HBcAg分子α螺旋上,構建了真核表達重組質粒pCMV-HA-HBcAg-VP1-VP4,并成功制備了FMDV-VLPs。因此,作為黏膜免疫主要成員的MCs識別FMDV-VLPs 分泌的細胞因子表達情況如何需要進行研究。
前期研究表明,重組FMDV VP1-VP4蛋白可以抑制腹腔肥大細胞(peritoneal mast cells, PMCs)分泌一系列細胞因子[18]。近年來,骨髓源肥大細胞(bone marrow-derived mast cells, BMMCs)被作為肥大細胞體外模型用于許多研究中,但BMMCs能否通過MR識別FMDV-VLPs產生免疫應答尚不清晰。所以,本研究用IL-3和SCF體外誘導得到高純度的BMMCs,將其分為BMMCs組、MR抑制劑處理組,并負載FMDV-VLPs,利用蛋白芯片檢測BMMCs上清中細胞因子表達,以揭示甘露糖受體在肥大細胞識別FMDV-VLPs 過程中發揮的作用,從而為新型口蹄疫疫苗研制提供新思路。
1 材料與方法
1.1 材料與主要試劑
CHO-K1細胞購自普諾賽生命科技有限公司;pCMV-HA質粒由河北農業大學動物醫學院趙款老師提供;6~8周齡C57BL/6N小鼠購自維通利華實驗動物技術有限公司; RPMI 1640購自美國Gibco公司;小鼠IL-3和SCF購自近岸蛋白質科技有限公司;非必需氨基酸、L-谷氨酰胺和甲苯胺藍染液購自北京索萊寶科技有限公司;Mannan購自北京索萊寶科技有限公司;APC anti-mouse FcεRIα Antibody、APC Armenian Hamster IgG Isotype Ctrl Antibody、FITC anti-mouse CD117 (c-Kit) Antibody和FITC Rat IgG2b κ Isotype Ctrl Antibody購自美國BioLegend公司;Mouse Cytokine Array C6購自美國RayBiotech公司; T4 DNA Ligase購自美國Promega公司;PrimeSTAR?Max DNA Polymerase 2×、限制性內切酶EcoRⅠ和XhoⅠ購自日本TaKaRa公司;2×Es Taq MasterMix(Dye)購自北京康為世紀科技有限公司;Trans5α 購自北京全式金生物技術有限公司;膠體金標記羊抗兔IgG購自北京博奧森生物有限公司。
1.2 方法
1.2.1 重組質粒pCMV-HA-HBcAg-VP1-VP4的構建 以pcDNA3.1(+)-HBcAg-VP1-VP4為模板(該質粒為本實驗室構建并保存,具體的構建策略請參見文獻[19]),使用引物F:5′-CCGGAATTCACCATGGACATTGAC-3′,R:5′-CCGCTC-GAGTCAATGGTGATGGTG-3′,擴增目的片段HBcAg-VP1-VP4;用XhoⅠ和EcoRⅠ將擴增出的PCR產物進行酶切,使用T4 DNA Ligase將純化后的酶切產物連接至pCMV-HA質粒(16 ℃連接過夜)。將重組質粒pCMV-HA-HBcAg-VP1-VP4轉化到感受態細胞Trans5α,將PCR鑒定陽性的菌送至中科希林生物科技有限責任公司進行測序。
1.2.2 重組質粒pCMV-HA-HBcAg-VP1-VP4的表達與鑒定 按照重組質粒pCMV-HA-HBcAg-VP1-VP4與Lipofectamine 2000 Reagent(Invitrogen)的質量體積比為2∶1、1∶1、1∶2、1∶3配比,轉染CHO-K1細胞后72、60、48和24 h,并以1∶10稀釋的兔抗FMDV VP1-VP4多克隆抗體(以重組VP1-VP4蛋白制備的兔源多克隆抗體)為一抗37 ℃孵育1 h,PBS洗滌3次后,用1∶100稀釋的FITC標記的山羊抗兔IgG 37 ℃孵育40 min,進行間接免疫熒光鑒定。
按照間接免疫熒光鑒定結果,使用重組質粒與Lipo 2000的質量體積比1∶1進行轉染,收集轉染后24、36、48和60 h的上清進行Western blot鑒定。聚丙烯酰胺凝膠電泳(SDS-PAGE)后轉移至PVDF膜上;5% 脫脂牛奶4 ℃封閉過夜;使用1∶3 000稀釋的His標簽的單克隆抗體為一抗室溫孵育3 h;TBST洗滌5次后,以1∶40 000稀釋的山羊抗鼠IgG(辣根過氧化物酶標記)作為二抗,室溫孵育1.5 h,TBST洗滌5次后,使用超敏ECL化學發光試劑盒室溫避光顯色3 min,使用化學發光凝膠成像儀進行成像。
1.2.3 FMDV-VLPs的表達與鑒定 按照重組質粒pCMV-HA-HBcAg-VP1-VP4與Lipo 2000的質量體積比為1∶1進行轉染,48 h收集細胞上清,使用KTATMepure 蛋白純化系統進行 Ni 柱親合層析純化重組蛋白。純化后的FMDV-VLPs保存于-80 ℃備用。將蛋白變性后,進行SDS-PAGE電泳試驗鑒定純化效果。為了進一步驗證純化后的蛋白是否為目標蛋白,進行了Western blot鑒定:取純化后的蛋白懸液 20 μL進行SDS-PAGE;轉移至PVDF膜上,5% 脫脂牛奶4 ℃封閉過夜;使用1∶1 000稀釋的兔抗FMDV VP1-VP4多克隆抗體為一抗室溫孵育3 h;TBST洗滌5次后以1∶40 000稀釋的HRP標記的山羊抗兔IgG作為二抗,室溫孵育1.5 h,TBST洗滌5次后用DAB避光顯色5 min。
對FMDV VLP進行膠體金免疫電鏡鑒定,具體步驟如下:取0.5 mg·mL-1FMDV-VLPs 100 μL,加入100 μL 1∶1 000稀釋的兔抗FMDV VP1-VP4 IgG, 37 ℃作用30 min; 加入0.01 mol·L-1PBS緩沖液1 mL,4 ℃,13 000 r·min-1離心20 min, 棄上清, 反復兩次。最后一次離心棄去上清,用0.01 mol·L-1PBS緩沖液100 μL將沉淀重懸;懸液中加入50 μL 1∶50稀釋的膠體金抗體,37 ℃孵育30 min;取0.01 mol·L-1PBS緩沖液1 mL加入混合物中,離心20 min,棄上清,重復此步驟一次,棄上清,用100 μL去離子水將沉淀重懸;將懸液加入到15 mL超濾管中,加12 mL去離子水混勻,4 000 r·min-1離心至500 μL,重復兩次,最后一次離心至200 μL。將樣品進行磷鎢酸負染色,并在透射電子顯微鏡下觀察。
1.2.4 小鼠BMMCs的制備及鑒定 小鼠BMMCs的收取及培養方法參照文獻[18]進行:無菌條件下取骨髓細胞,用培養液(含10% FBS RPMI 1640培養基、0.1 mmol·L-1非必需氨基酸、2 mmol·L-1谷氨酰胺、10 ng·mL-1IL-3、20 ng·mL-1SCF)重懸細胞,調整細胞濃度為1×106個·mL-1,鋪到直徑10 cm細胞培養皿中,置于37 ℃、5% CO2培養箱中培養。每隔7 d更換培養液,每次收集上清中的細胞繼續進行培養。
收集BMMCs懸液,調整細胞濃度為1×106個·mL-1,取20 μL的細胞懸液進行涂片;用4%多聚甲醛固定細胞3 min,然后用PBS(pH7.2)沖洗3次;滴1 mL 0.1%甲苯胺藍染色30 s,用蒸餾水沖掉甲苯胺藍溶液;300 mL·L-1乙醇-PBS(pH 7.2)分色至細胞質呈現藍色,細胞核為紫紅色,立即用蒸餾水沖洗;中性樹脂封片,顯微鏡下觀察。
取在體外培養了8周的BMMCs進行流式細胞術鑒定。將BMMCs調整細胞濃度為1×106個·mL-1。每個1.5 mL離心管中加入100 μL細胞懸液,加入含3% BSA的PBS稀釋的抗體[終濃度為0.1 μg·mL-1的APC anti-mouse FcεRIα Antibody和FITC anti-mouse CD117 (c-Kit) Antibody,并用APC Armenian Hamster IgG Isotype Ctrl Antibody和FITC Rat IgG2b, κ Isotype Ctrl Antibody作同種型對照]。4 ℃避光孵育30 min。1 700 r·min-1離心5 min,棄掉上清,用含3% BSA、1%疊氮化鈉的冰冷PBS溶液1 mL重懸,1 700 r·min-1離心5 min重復3次。使用流式細胞儀檢測。
1.2.5 細胞因子芯片檢測 將鑒定后的BMMCs按1×106·mL-1鋪到24孔板中,每孔500 μL細胞懸液。將BMMCs分為3組,分別為MR抑制組(iMR-VLP組)、MR未抑制組(VLP組)和空白對照組(Control組),每組3個重復。MR抑制組的培養孔中加入終濃度為3 mg·mL-1的Mannan[19],2 h后,給MR體抑制組和MR未抑制組加入終濃度為20 μg·mL-1的FMDV-VLPs,空白對照組不做處理。將細胞培養板置于37 ℃,5% CO2培養箱中培養。24 h后收集細胞上清使用Mouse Cytokine Array C6試劑盒對細胞因子進行檢測。
1.2.6 數據分析 使用Graphpad Prism 5對掃描數據進行One-way ANOVA統計學分析。P<0.05為差異顯著,P<0.01為差異極顯著,并做差異分析柱狀圖和熱圖。
2 結 果
2.1 重組質粒pCMV-HA-HBcAg-VP1-VP4表達與鑒定
2.1.1 表達與鑒定 將測序成功的重組質粒pCMV-HA-HBcAg-VP1-VP4按照不同條件染轉染CHO-K1細胞,并進行間接免疫熒光和Western blot鑒定。結果顯示:在轉染后24 h,重組質粒與Lipo 2000質量體積比為1∶1的條件下重組質粒pCMV-HA-HBcAg-VP1-VP4的表達效率最高(圖1);在48 h左右并且重組質粒與Lipo 2000質量體積比為1∶1的條件下,細胞上清中目的蛋白表達效率最高(圖2)。
圖1中a、b、c和d分別為質粒與Lipo 2000質量體積比為2∶1、1∶1、1∶2和1∶3的條件下,在24 h時細胞的間接免疫熒光鑒定結果
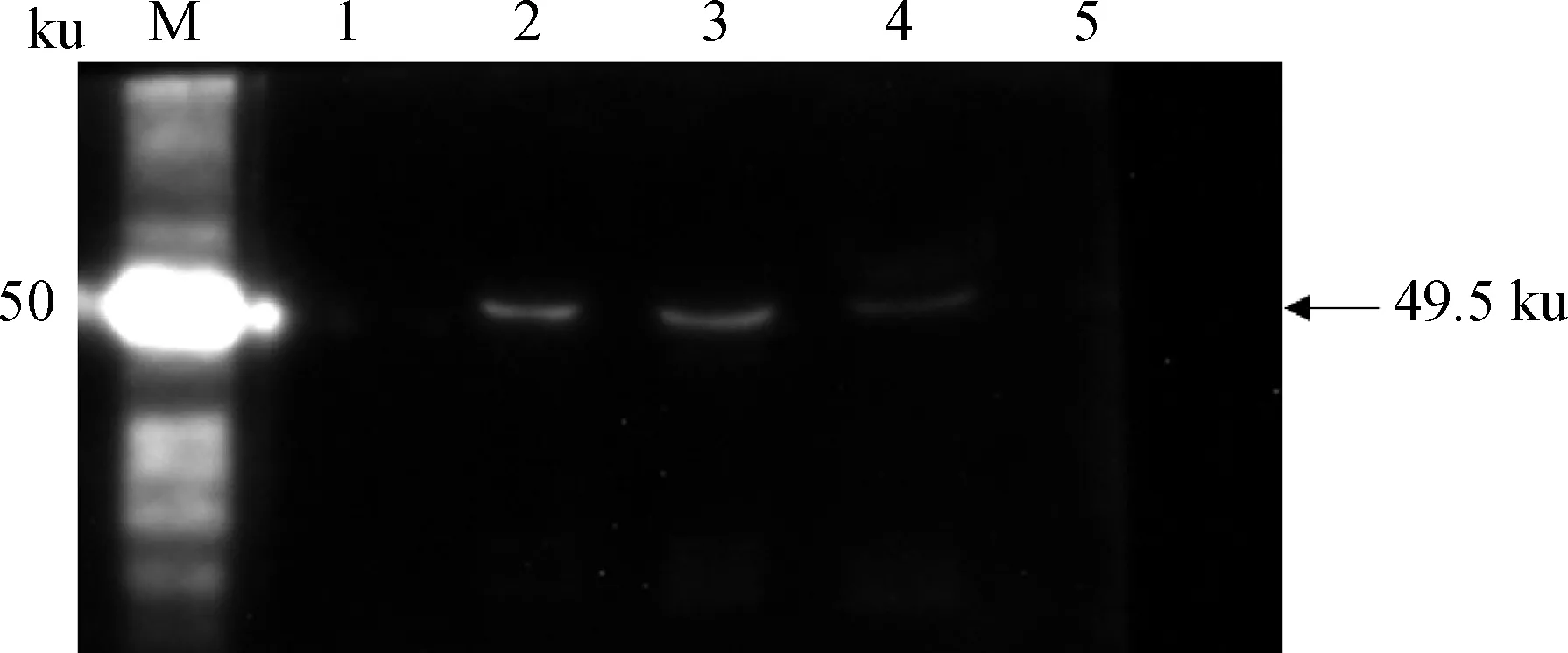
不同時相細胞上清的Western blot鑒定結果(重組質粒與Lipo 2000質量體積比為1∶1)。M.蛋白質相對分子質量標準;1、2、3、4分別為轉染后24、36、48、60 h的細胞上清;5.未轉染的細胞上清
2.1.2 純化與鑒定 將重組質粒pCMV-HA-HBcAg-VP1-VP4按照與Lipo 2000質量體積比為1∶1轉染CHO-K1細胞,在48 h收集細胞上清用鎳柱進行純化,然后進行SDS-PAGE鑒定和Western blot鑒定。SDS-PAGE的結果顯示,在49.5 ku左右處出現了單一條帶,與預期的大小相符,表明純化效果較好(圖3a);Western blot鑒定結果顯示在49.5 ku左右處出現了一條特異性條帶,提示純化后的蛋白是目標條帶(圖3b)。為了鑒定重組質粒表達后是否自我組裝成VLPs,進行了免疫透射電鏡觀察。結果顯示,透射電鏡下可以看到重組蛋白形成了顆粒,粒子直徑大約為30 nm,并且在粒子周圍,有直徑10 nm左右膠體金顆粒附著(圖3c),表明重組質粒成功組裝成FMDV-VLPs。

a.SDS-PAGE鑒定結果(M.蛋白質相對分子質量標準;1.純化前的細胞上清;2.純化后的重組蛋白); b.Western blot鑒定結果(M.蛋白質相對分子質量標準;3.純化后的重組蛋白);c.膠體金免疫電鏡鑒定結果,黑色箭頭為口蹄疫病毒樣顆粒(約30 nm),紅色箭頭為膠體金顆粒(約10 nm)
2.2 小鼠BMMCs的甲苯胺藍染色及流式細胞術鑒定
BMMCs通過甲苯胺藍染色后,可見細胞質呈藍色,細胞核呈紫紅色,無脫顆粒現象(圖4a);流式細胞術結果顯示,FcεRIα和CD117雙陽性的細胞占97.9%左右,表明制備的BMMCs純度良好,符合試驗要求(圖4b)。
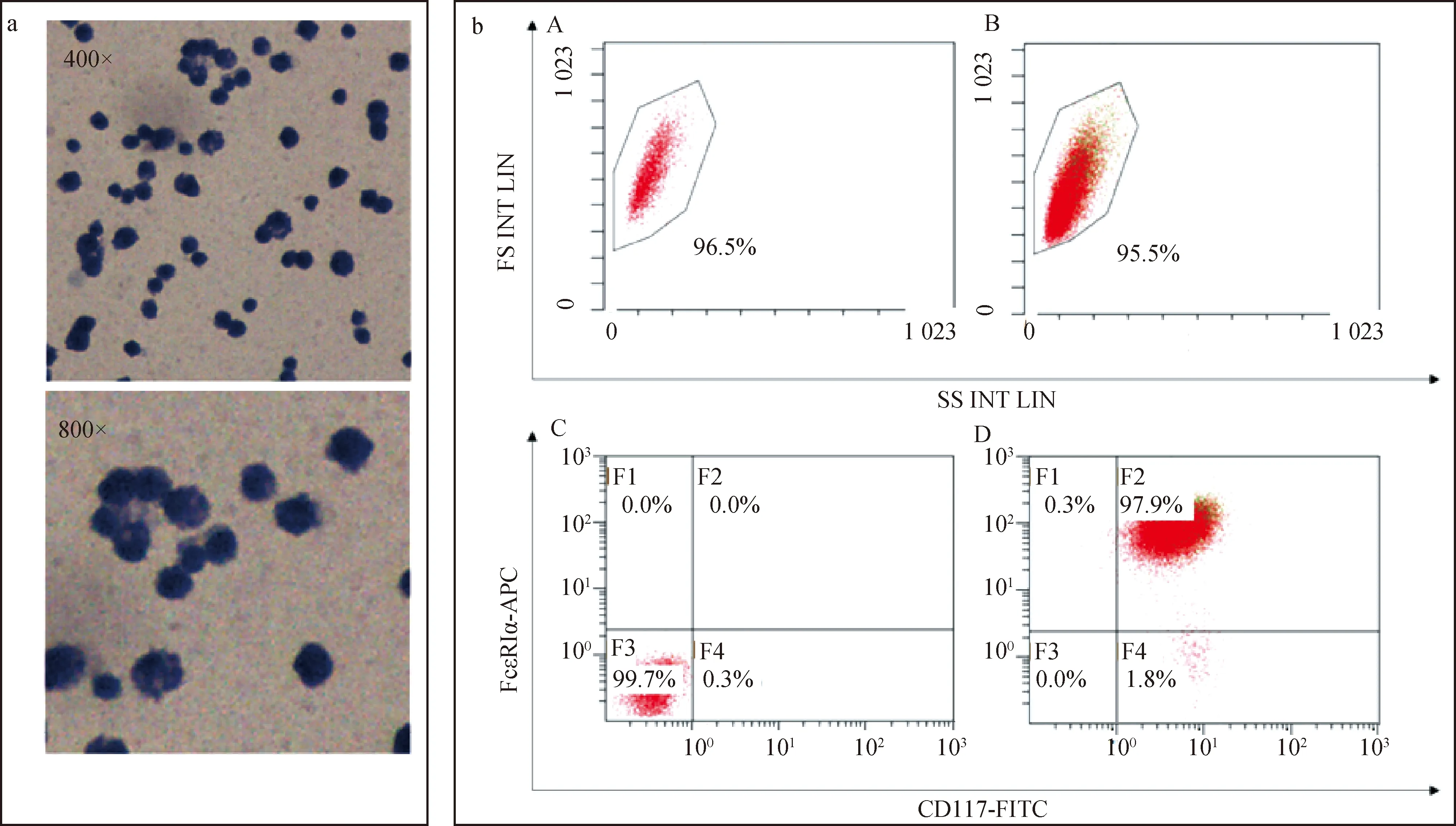
圖4a為BMMCs甲苯胺藍染色結果;圖4b為BMMCs流式細胞術結果[A、B.使用FSC(正向散射)和SSC(側散射)作為參數對BMMCs進行分析; C.使用APC Armenian Hamster IgG Isotype Ctrl 抗體和κ Isotype Ctrl 抗體作為同型對照;D.使用APC anti-mouse FcεRIα抗體和FITC anti-mouse CD117 (c-Kit) 抗體標記BMMCs,進行流式細胞術鑒定]
2.3 不同處理組BMMCs表達的細胞因子差異分析
為了分析細胞因子的差異變化,使用Image J軟件對芯片檢測結果進行灰度值(gray values)分析。結果顯示,相比于control組,VLP組BMMCs分泌的CCL21、IL-9、IFN-γ、IL-2、IL-4、IL-15、IL-17A、IL-21、IL-12p70、TNF-α、IL-1α、CCL17和L-selectin顯著上調(P<0.05);iMR-VLP組相比于VLP組,BMMCs分泌的IL-9、IL-10、CCL19和CCL21顯著上調(P<0.05),而IFN-γ、IL-2、IL-4、IL-15、IL-21、IL-12 p70、IL-1α、CCL17和 TNF-α顯著下調(P<0.05)(圖5)。
*.P<0.05;**.P<0.01;***.P<0.001
2.4 不同處理組BMMCs的蛋白質表達譜檢測結果
使用MeV軟件對“2.3”得出的不同處理組BMMCs分泌的細胞因子的表達量進行聚類分析,作聚類熱圖(圖6)。VLP組相比于Control組,IL-1、IL-2、TNF-α、IFN-γ和IL-12等Th1細胞因子以及IL-21、IL-4等Th2型細胞因子表達均發生了上調,而iMR-VLP組與VLP組相比,抑制MR后,IL-1、IL-2、TNF-α、IFN-γ和IL-12等Th1細胞因子以及IL-21、IL-4等Th2型細胞因子表達呈下調變化,CCL19、IL-9等卻發生上調。結果表明,BMMCs的 MR受體在識別FMDV-VLPs過程中發揮著重要的調節作用。

圖6 不同處理組BMMCs的蛋白質表達差異熱圖
3 討 論
MR作為肥大細胞表面重要的模式識別受體,在肥大細胞執行其免疫調節功能中起著非常重要的作用[10,20]。因此,作者構建了重組pCMV-HA-HBcAg-VP1-VP4質粒,并且成功制備了FMDV-VLPs,用FMDV-VLPs負載經過甘露糖受體抑制劑處理的BMMCs,檢測上清中細胞因子的表達,以此來闡釋甘露糖受體在骨髓源肥大細胞識別FMDV-VLPs過程中的作用。
在抗FMDV感染的過程中,IFN-γ和TNF-α發揮重要的作用。IFN-γ負責調控FMDV感染的免疫應答和抑制FMDV的復制,而TNF-α能夠促進固有免疫向適應性免疫的轉變,可以通過Th1通路激活T細胞,促使細胞免疫應答的發生[21];IL-10是主要的抑制炎癥的細胞因子,在FMDV感染的過程中,豬血清中的IL-10逐漸升高,在3、4 d達到頂峰,抑制T細胞和B細胞的增殖,因此,IL-10升高被認為與FMDV感染時的免疫抑制有關[22];IL-6是一種多效細胞因子,在激活和調節免疫反應中起重要的作用,研究表明,在FMDV感染的過程中,仔豬外周血IL-6會有明顯的升高,推測其可能與不同年齡FMDV致病性差異有關[21]。
先前的研究表明, 重組FMDV VP1-VP4蛋白負載腹腔肥大細胞(PMCs)后CCL19、L-selectin、CCL17和TNF-α等34個蛋白的表達水平顯著低于對照組(P<0.01);而抑制PMCs的MR后再負載VP1-VP4蛋白,TNF-α、IL-6、CCL19、IL-15、IL-9等功能分子的表達顯著升高[18]。而本研究結果顯示,負載VLPs后BMMCs分泌的TNF-α、IL-1、IL-4、IL-15、IFN-γ、IL-2、CCL17等細胞因子的表達顯著高于對照組(圖6),相比于負載FMDV-VLPs且MR未抑制組(VLP組),MR受體抑制組(iMR-VLP)的BMMCs分泌的TNF-α、IL-1、IL-4、IL-15、IFN-γ、IL-2等細胞因子均有明顯下降的趨勢,而IL-9、CCL19、CCL21等細胞因子的分泌則有明顯上升的趨勢(圖5、6)。其中IL-1a、IL-2、INF-γ、TNF-α、IL-17、IL-15等Th1細胞因子,可促進炎癥形成,并促進細胞免疫應答,有利于病原清除;IL-4和IL-10為Th2 細胞因子,可以抑制炎癥形成并有助于B細胞的增殖與分化;CCL17、CCL19和CCL21為趨化因子,主要功能是在炎癥和免疫應答過程中介導淋巴細胞的遷移。造成細胞因子分泌差異的可能原因:1)VLPs 在一定程度上可以緩解重組VP1-VP4蛋白帶來的免疫抑制。2)可能與肥大細胞本身有關。骨髓干細胞在IL-3和SCF共同誘導下可以分化成均質較成熟的BMMCs,和PMCs均為結締組織型肥大細胞[23-24]。而直接從腹腔細胞中利用Percoll分離液分離的腹腔肥大細胞雖然經甲苯胺藍染色檢測純度較高,但是利用CD117及FcεRIα 抗體鑒定顯示純度較低(數據沒有列出)。研究表明,與BMMCs相比,PMCs活化后不分泌或分泌少量的新合成的促炎因子,比如TNF-α和IL-6,但是IL-4、IL-10 和IFN-γ 兩者之間沒有差異[25]。并且,轉錄組學研究結果表明,IL-3誘導形成的BMMCs與成熟的PMCs的轉錄本在促炎因子方面,PMCs明顯低于BMMCs[26]。本研究中不同處理是否會影響甘露糖受體在BMMCs的表達以及該受體在BMMCs 和PMCs上表達是否有差異有待進行深入研究。
目前,體外培養MCs是研究肥大細胞生物學作用的重要方法。一般分為從腹腔分離PMCs和從骨髓中分離不成熟的MCs前體細胞,體外分化為成熟的BMMCs[26]。Akula等[23]發現,雖然相比于BMMCs,PMCs中一些細胞因子或者細胞因子受體幾乎不表達,包括IL-1β、IL-6、IL-10受體α鏈、IL-4受體α鏈、IL-2受體α鏈、TGF-β受體等,但是BMMCs與PMCs均表現出結締組織型肥大細胞的特性。并且,使用IL-3和SCF聯合誘導比IL-3單獨誘導形成的BMMCs 成熟度更高[23,27]。PMCs可以從腹腔直接分離得到,但數目很低(約1×105個·只-1小鼠),不僅難滿足試驗需求,而且純度變化大;腹腔細胞可以在體外繼續培養8周,可以得到成熟的純度較高的肥大細胞(PCMCs)[23]。骨髓干細胞在IL-3和SCF的誘導下可以分化成比較成熟BMMCs(5~8周),純度很高且細胞數目可以滿足試驗需求。鑒于不同組織中的MCs在不同組織中甚至同一組織中不同部位的表型不完全相同,體外培養的 BMMCs 與體內的MCs存在的一定差異[23,28],并且不同的刺激亦影響MCs的表型,體外研究應該結合體內試驗以得出更加科學的結論。
4 結 論
BMMCs可以通過MR識別FMDV-VLPs,促進IL-1α、IL-2、IL-4、IL-15、IL-17A、TNF-α、IFN-γ、CCL-17和L-Selectin表達,下調IL-9、IL-10、CCL19和CCL21的表達,表明FMDV-VLPs可能通過BMMCs的甘露糖受體增強Th1型細胞因子分泌、有效抑制能引起免疫抑制的IL-10的分泌,并降低介導T細胞再循環的CCL19和CCL21的形成。這為基于MCs的甘露糖受體應答效應的口蹄疫新型疫苗的研發提供了理論依據和新思路。
