基于WGCNA技術研究驢肉嫩度基因及代謝物調控網絡
2022-11-29 13:39:32李武峰邱麗霞關家偉
畜牧獸醫學報 2022年11期
李武峰,邱麗霞,關家偉,李 麗,杜 敏
(1.山西農業大學生命科學學院,太谷 030801; 2.美國華盛頓州立大學動物科學系,普耳曼 99164-6310)
驢(Equusasinus)是傳統的役用家畜,肉質鮮美、營養豐富,其包含亞麻酸在內的多不飽和脂肪酸,種類豐富且含量高,易于被人體吸收利用[1]。廣靈驢屬大型驢種,其體軀粗壯、產肉性能較好、食用口感與風味俱佳,是中國的優勢驢種之一[2-3]。肉的好壞和質量一般可通過嫩度、肌內脂肪(intramuscular fat, IMF)含量、系水力、肉色、風味等指標進行綜合評定,而嫩度則作為重要感官指標用來直接衡量肉品質[4]。嫩度是咀嚼肉的感覺,可以很大程度上影響和決定消費者購買選擇與欲望,因此探究驢肉嫩度的調控機制具有極其重要的價值和意義。
本試驗以廣靈驢為研究對象,以剪切力和IMF為表型信息,通過WGCNA技術篩選與驢肉嫩度相關的基因及代謝物并進行聯合分析,解析相應的驢肉嫩度調控機制,為廣靈驢肉質嫩度分子改良和分子育種提供理論依據。
1 材料與方法
1.1 試驗動物及數據獲取
選用廣靈驢作為試驗對象,選取30頭在相同環境與飲食條件下生長、24~36月齡的雌性廣靈驢(平均體重236.10 kg),獲取途徑來源于山西省忻州市繁峙縣田源毛驢養殖科技發展有限公司。對動物進行屠宰后于30 min內取第12~13肋骨間的背最長肌組織,立即儲存到液氮中供后續處理。本研究運用的前期轉錄組及代謝組樣本是依據剪切力大小與肌內脂肪含量從30頭廣靈驢中選取的最具差異的14個樣本,前期轉錄組及代謝組數據分析見[14-15]。為了讓片段數能真實的反映轉錄本表達水平,采用FPKM(fragments Per kilobase per million)方法作為衡量基因表達水平的指標,FPKM計算公式如下[16]:
其中,Xi為唯一比對到基因的片段數;li為基因的長度;N為唯一比對到參考基因組的總片段數。
1.2 試驗試劑及儀器
石油醚為Merck級別;索氏抽提器SXT-02(上海洪紀儀器設備有限公司);MAQC-12肌肉嫩度儀(南京銘奧儀器設備有限公司);電熱恒溫水浴鍋(上海一恒科學儀器有限公司)。
1.3 剪切力及肌內脂肪含量測定
剪切力測定:參照國家標準NY_T 1180-2006《肉嫩度的測定 剪切力測定法》測定驢肉剪切力。將驢肉置于80 ℃水浴鍋中加熱,利用溫度計監測肉樣中心溫度,達到70 ℃后放置室溫環境冷卻,后用肌肉嫩度儀測定肉樣的剪切力大小,設置3個樣本重復。
肌內脂肪含量測定:參照國家標準GB5009.6-2016《食品安全國家標準 食品中脂肪的測定》要求提取測定廣靈驢背最長肌的肌內脂肪含量。利用索氏抽提儀提取驢肉肌內脂肪,溶劑為石油醚,每個樣本3個重復。
1.4 基因共表達網絡的構建
利用R 4.1.0軟件中的WGCNA 1.7.0包將14個樣本的所有表達數據進行基因共表達網絡分析。利用pickSoftThreshold函數計算最佳軟閾值,選取無尺度網絡擬合指數R2>0.8時power值最小的數為最佳power值。利用WGCNA包中的blockwiseModules函數構建共表達矩陣,相似模塊合并閾值設置0.25(mergeCutHeight=0.25),TOM類型為unsigned,deepSplit = 1,每個模塊內的最小基因數設置為30(minModuleSize = 30),其他參數按照默認設置。
1.5 目標模塊的篩選
對模塊內的基因進行PCA分析,將主成分1作為模塊特征向量(module eigengenes, MEs)。為篩選出與IMF和剪切力密切相關的模塊,計算MEs與IMF和剪切力之間的相關系數r以及相應P值。對特異性模塊進行篩選,|r|≥0.5且P≤0.05范圍的模塊可進行后續分析。
2)將適應值排在第1/3Np+1(Np為種群規模)的個體記為mk。對每維個體,求適應值大于mk且符合式(18)范圍的所有個體集合,記為Ln。如果Ln中個體數大于k/2,則表示優異種群在該位置信息失效。
1.6 模塊基因的GO、KEGG富集分析
應用R軟件中的topGO 2.40.4包對特異性模塊基因進行基因本體(gene ontology, GO)功能富集分析,當P<0.05時認為該GO條目顯著富集。運用Cluster Profiler 3.16.1包進行京都基因與基因組百科全書(kyoto encyclopedia of genes and genomes, KEGG)功能富集分析,當P<0.05時認為該信號通路的富集具有顯著性。
1.7 代謝組WGCNA分析
代謝物共表達網絡的構建是選取無尺度網絡擬合指數R2>0.85時power值最小的數為最佳power值,其余步驟及參數與“1.4”部分相同,目標模塊的篩選同“1.5”部分。使用美吉云平臺(https://www. http://cloud.majorbio.com/)的自建數據庫對目標模塊內的代謝物進行人類代謝數據庫(human metabolome database, HMDB)化合物分類和KEGG功能富集分析。
1.8 轉錄組代謝組聯合分析
利用KEGG數據庫對目標基因進行共富集分析,通過Cytoscape 3.7.2軟件中的MetScape預測聯合通路中基因與代謝物互作網絡圖。然后用R語言cor運算基因和代謝物的Pearson相關系數,繪制相關性聚類熱圖并結合相關系數篩選出大于0.5的基因和代謝物,構建轉錄-代謝物網絡。
1.9 統計分析
剪切力及肌內脂肪測定數據采用Excel 2019軟件進行加權平均值計算,結果以每個樣品3個重復計算的平均值表示。以P<0.05標準篩選GO和KEGG顯著富集結果,對各通路中的基因及代謝物進行Pearson相關性分析,以相關性大于0.5為標準,使用R 4.1.0軟件對結果進行可視化呈現。
2 結 果
2.1 剪切力和肌內脂肪含量測定結果
14個廣靈驢背最長肌表型數據檢測結果如表1所示。從30頭動物中抽取了6頭具有高、低肌內脂肪含量顯著差異的24月齡廣靈驢和8頭具有高、低剪切力顯著差異的36月齡廣靈驢,從表中可以看到,肌內脂肪含量與剪切力大小呈負相關。
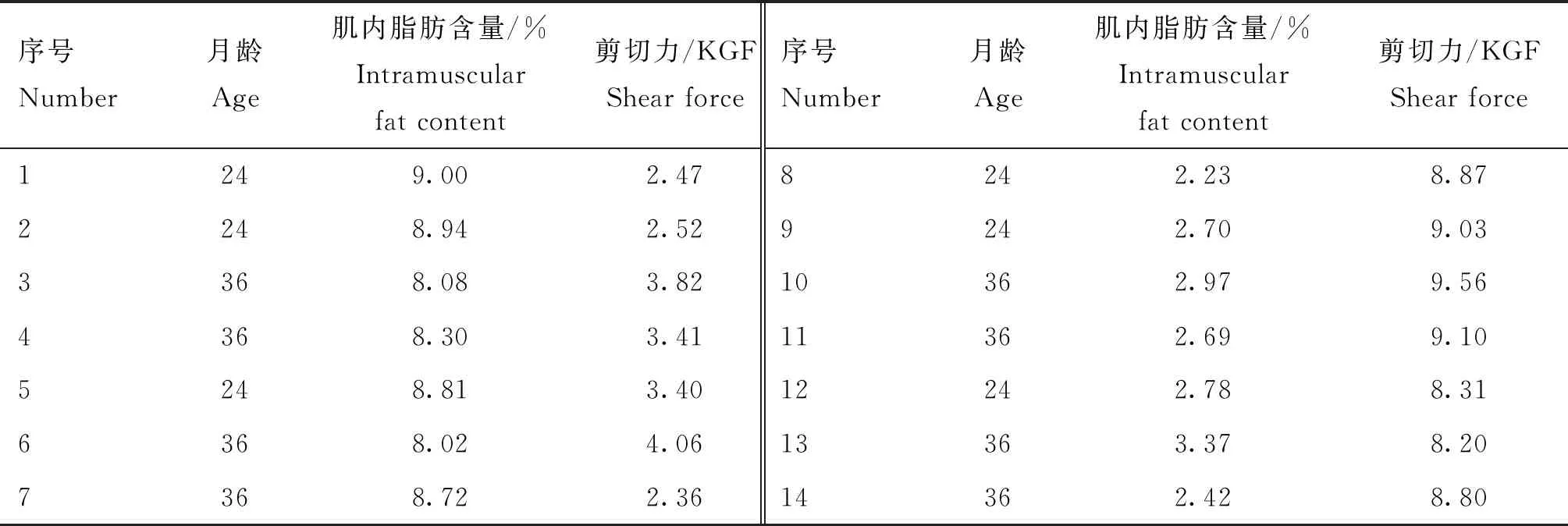
表1 廣靈驢肉用指標相關的描述性統計
2.2 基因共表達網絡構建及關鍵模塊鑒定
通過軟閾值選擇最佳β值,當power值等于5,無尺度網絡擬合指數R2>0.8(圖1A)。利用TOM值進行層次聚類,通過混合動態剪切來劃分模塊,構建出了28個模塊(圖2A),通過顏色差異區分不同模塊。每個模塊的詳細基因數目見圖1B,其中Turquoise模塊內的基因數目最多,共5 362個,White模塊內的基因數目最少,為34個,其他模塊數目介于43~2 779之間。
模塊與IMF和剪切力的相關性如圖2B所示,本研究以|r|≥0.5及P≤0.05為判定標準共篩選出3個關鍵模塊。其中Darkgreen模塊(r=0.73,P=0.003)含有NIT2、LST1、SOCS7、GAPT、PAPN2等95個基因;Greenyellow模塊(r=0.52,P=0.05)含有LSR、MYOG、AKT1、CD5、ELOVL2等372個基因以及Darkgrey模塊(r=0.56,P=0.04)含有CXCL9、CAD1、FADS6、ADH4、TSR1等73個基因,3個模塊均與IMF顯著正相關,Darkgreen模塊(r=-0.64,P=0.01)與剪切力顯著負相關。為了驗證和解析模塊的功能,本研究對這3個模塊進行后續分析。
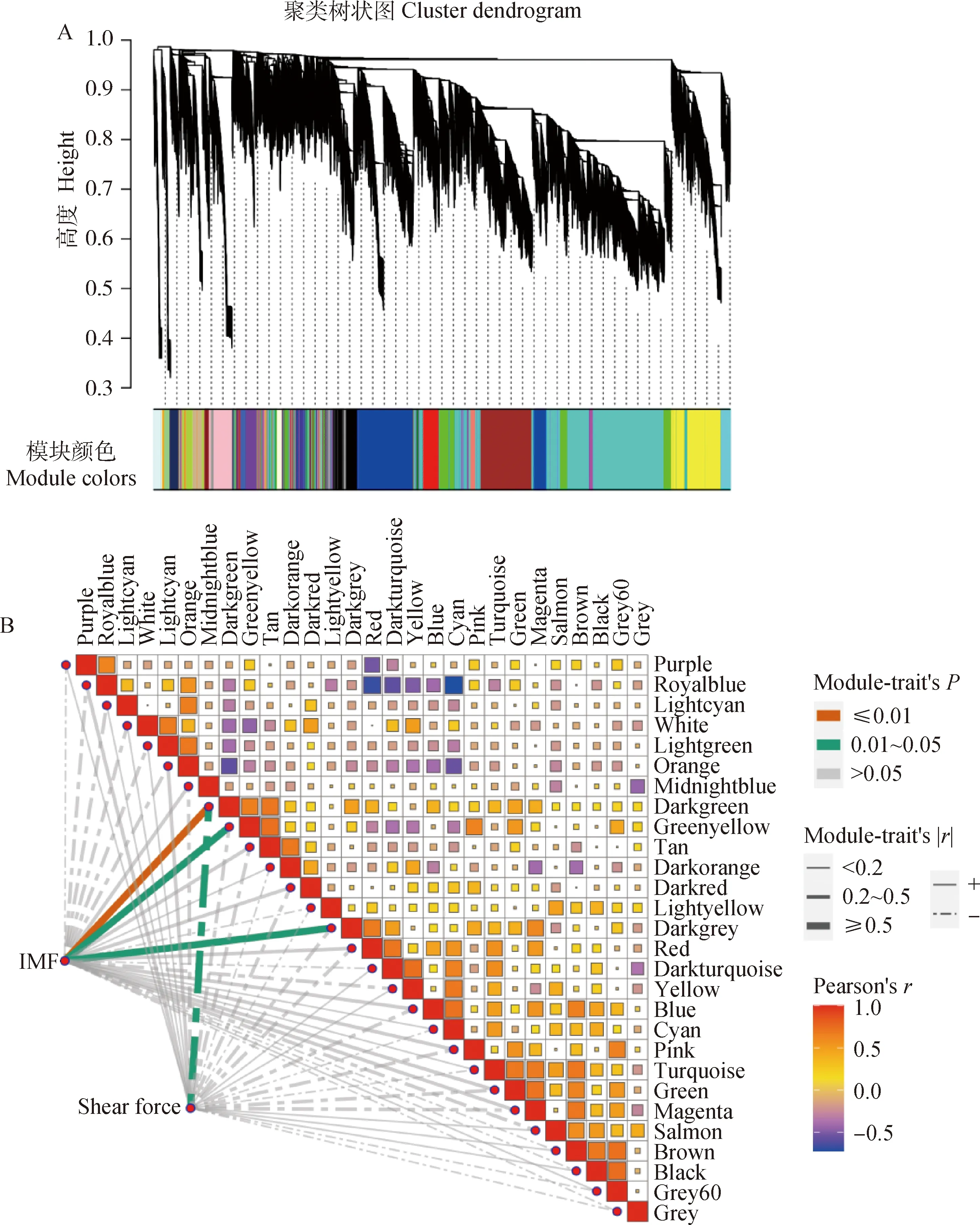
A.基因聚類模塊劃分;B.IMF、剪切力與基因模塊特征向量的交匯結果
2.3 模塊基因GO功能富集分析
本研究對Greenyellow、Darkgrey以及Darkgreen三個模塊進行GO功能富集分析(圖3)。Greenyellow模塊顯著富集到262個生物過程、37個細胞組分、53個分子功能,主要在甘油磷脂的生物合成、脂肪酸連接酶活性、脂肪酸β-氧化、脂質氧化、脂肪酸結合等功能上富集。Darkgrey模塊顯著富集到93個生物過程、20個細胞組分、30個分子功能,主要在羧酸生物合成過程、脂肪酸生物合成過程、G蛋白偶聯受體活性、蛋白磷酸酶抑制劑活性、ATP酶調節活性等功能上富集。Darkgreen模塊顯著富集到105個生物過程、28個細胞組分、21個分子功能,主要在肌管分化、細胞大分子分解代謝過程、肌肉器官發育、橫紋肌組織發育、肌肉細胞遷移、鈣離子結合、細胞骨架蛋白結合、肌動蛋白絲結合等功能上富集。
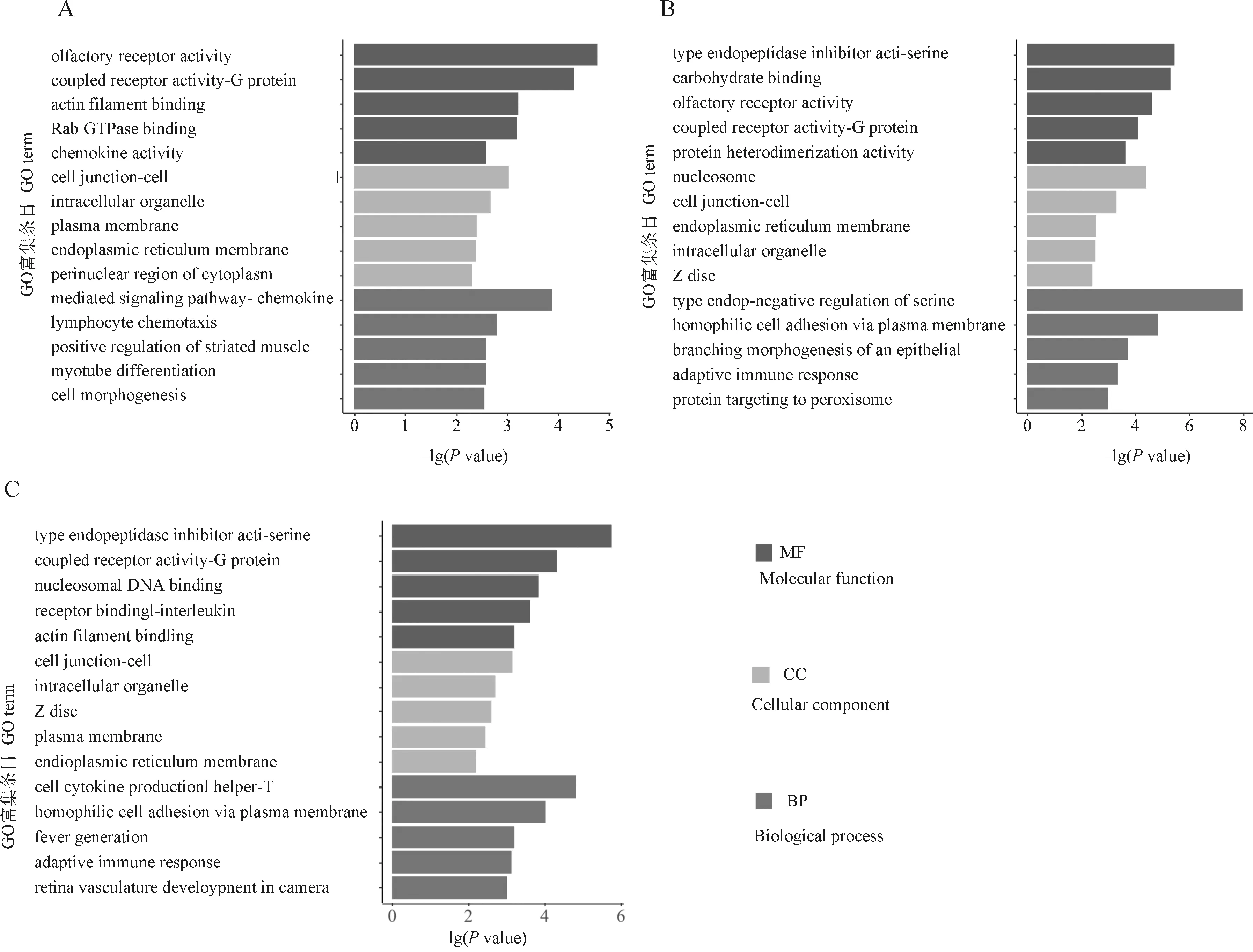
圖3 Greenyellow(A)、Darkgrey(B)、Darkgreen(C)差異模塊基因GO富集結果
2.4 模塊基因KEGG通路富集分析
對3個模塊進行KEGG功能富集分析,結果見圖4。Greenyellow模塊的富集結果顯示涉及267條通路,發現有40條通路被顯著富集(P<0.05),主要有HF-1信號通路、脂肪酸代謝、Wnt信號通路、IL-17信號通路以及花生四烯酸代謝等。Darkgrey模塊的富集結果顯示涉及89條通路,其中有9條通路被顯著富集(P<0.05),即藥物代謝-細胞色素P450、病毒蛋白與細胞因子和細胞因子受體的相互作用、細胞色素P450對異生素的代謝、藥物代謝-其他酶、致心律失常性右心室心肌病、視黃醇代謝、化學致癌、牛磺酸和亞牛磺酸代謝和軸突再生。Darkgreen模塊富集結果顯示涉及126條通路,其中有細胞因子-細胞因子受體相互作用、刺猬信號通路、神經營養因子信號通路、氨酰-tRNA生物合成、庫欣綜合征、胃癌、細胞凋亡、JAK-STAT信號通路、子宮內膜癌、脂肪細胞因子信號通路、甲狀腺激素合成、病毒蛋白與細胞因子和細胞因子受體的相互作用、EGFR酪氨酸激酶抑制劑耐藥、cAMP信號通路、致心律失常性右心室心肌病、甲狀旁腺激素的合成、分泌和作用共16條通路顯著富集(P<0.05)。
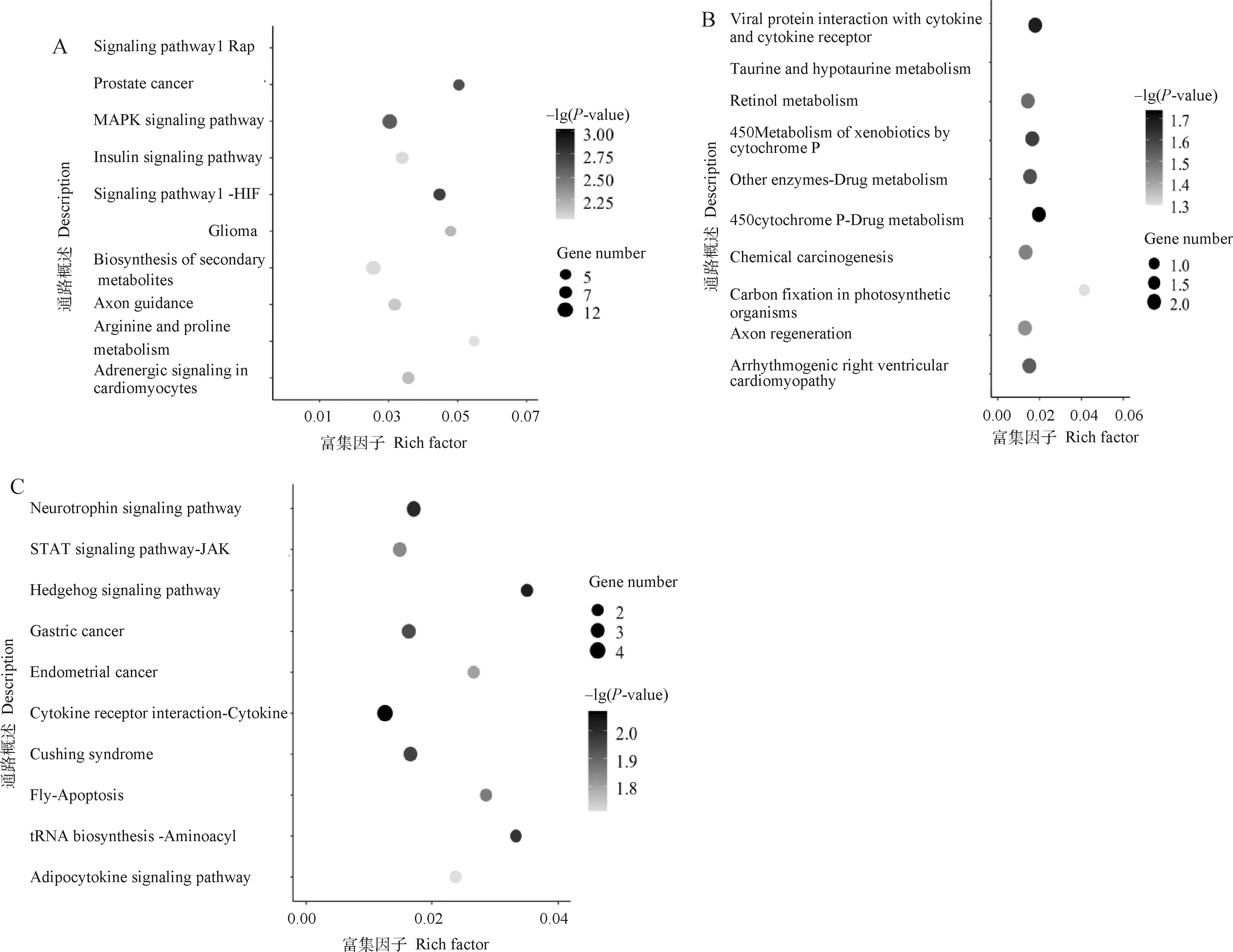
圖4 Greenyellow(A)、Darkgrey(B)以及Darkgreen(C)模塊基因Top10 KEGG富集散點圖
2.5 代謝物共表達網絡構建及關鍵模塊鑒定
利用pickSoftThreshold函數篩選合適的軟閾值,當power值為14時,R2>0.85(圖5A),層次聚類后共構建28個模塊(圖6A)。各模塊的代謝物數目如圖5B所示,其中Turquoise模塊內的代謝物數目最多,共2 238個,White模塊內的代謝物最少,為34個,其他模塊介于45~1 808之間。
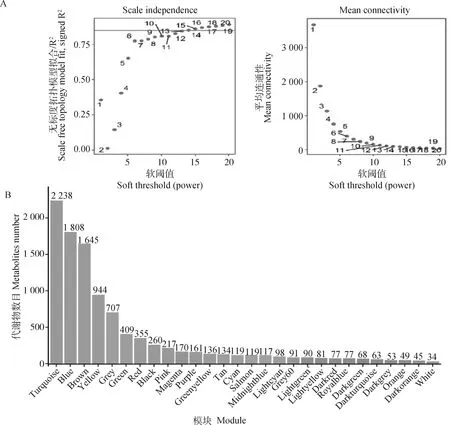
A.確定最佳軟閾值;B.各模塊中的代謝物數目分布
代謝物模塊與IMF和剪切力的相關性如圖6B所示,基于|r|≥0.5及P<0.05選定關鍵模塊,其中Brown模塊(r=0.76,P=0.004)、Yellow模塊(r=-0.63,P=0.03)以及Darkgreen模塊(r=-0.72,P=0.008)與IMF顯著相關。Brown模塊(r=-0.79,P=0.002)、Lightcyan模塊(r=-0.71,P=0.01)、Yellow模塊(r=0.69,P=0.01)、Darkgreen模塊(r=0.7,P=0.01)以及Darkred模塊(r=0.71,P=0.01)與剪切力顯著相關。然而,關鍵模塊中的代謝物大部分無法識別,Brown、Yellow、Darkgreen、Lightcyan以及Darkred模塊中代謝物分別有1 645、944、68、98、77個,其中可識別的代謝物分別有171、55、7、11、3個。Brown模塊中的代謝物包括丙氨酰-賴氨酸、L-脯氨酰胺、L-酪氨酸、腺嘌呤、丙氨酰-亮氨酸、腺苷酸基琥珀酸等,Yellow模塊中的代謝物包括N-乙酰-L-組氨酸、肌肽、尿烷酸、荊芥內酯、黃嘌呤酸8-O-硫酸鹽等。由于Darkgreen、Lightcyan以及Darkred模塊中可識別代謝物太少,因此本研究主要就Brown以及Yellow模塊進行后續分析。
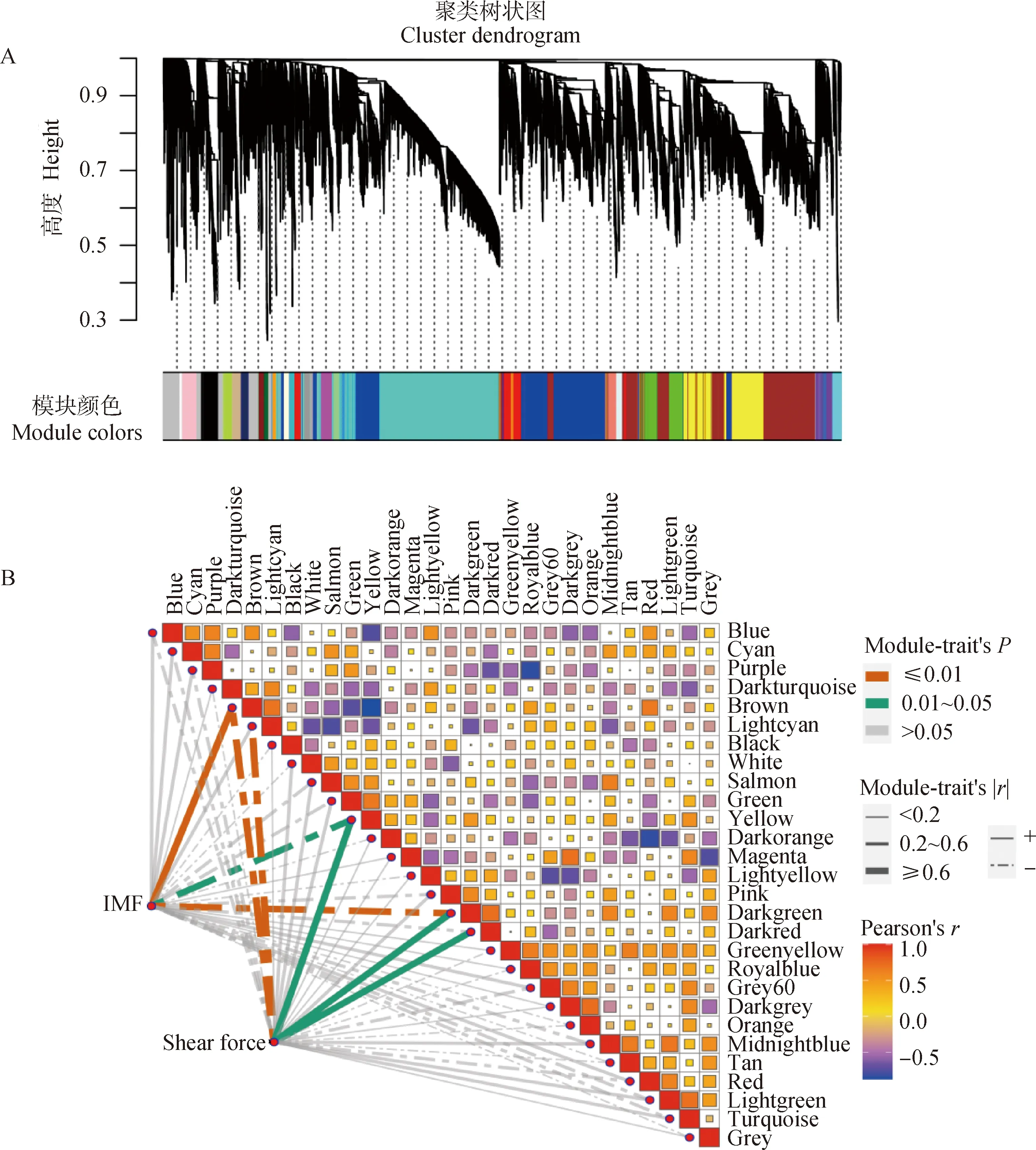
A.代謝物聚類及模塊劃分;B.IMF、剪切力與代謝物模塊特征向量的交匯結果
2.6 代謝物模塊HMDB化合物分類
將Brown和Yellow模塊注釋到HMDB數據庫,結果如圖7所示。Brown和Yellow模塊中脂質和類脂質分子占比最多(分別占37.98%和35.00%),其余的有機酸及其衍生物(分別占30.23%和25.00%)、有機雜環化合物(分別占12.40%和12.50%)以及有機氧化合物(分別占6.20%和15.00%)占比比較多。
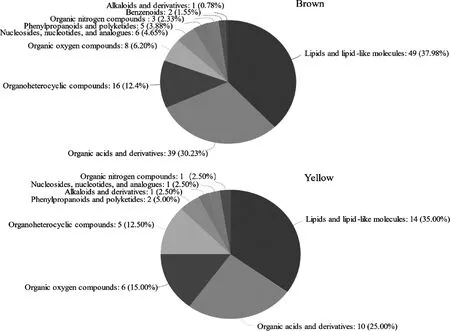
圖7 Brown、Yellow模塊代謝物在HMDB數據庫中的分類
2.7 代謝物模塊KEGG功能富集分析
對Brown和Yellow模塊內的代謝物進行KEGG功能富集分析,圖8展示了各模塊富集前10的代謝通路。Brown模塊共富集到96條代謝通路,其中37條通路顯著富集(P<0.05),主要有蛋白質消化吸收、谷胱甘肽代謝、FoxO信號通路、檸檬酸循環、精氨酸和脯氨酸代謝、甘油磷脂代謝等。Yellow模塊共富集到28條通路,其中僅有甘氨酸、絲氨酸和蘇氨酸代謝、嘌呤代謝、β-丙氨酸代謝、甘油磷脂代謝、泛酸和輔酶A生物合成、系統性紅斑狼瘡、組氨酸代謝、利什曼病和半胱氨酸和蛋氨酸代謝9條通路顯著富集(P<0.05)。
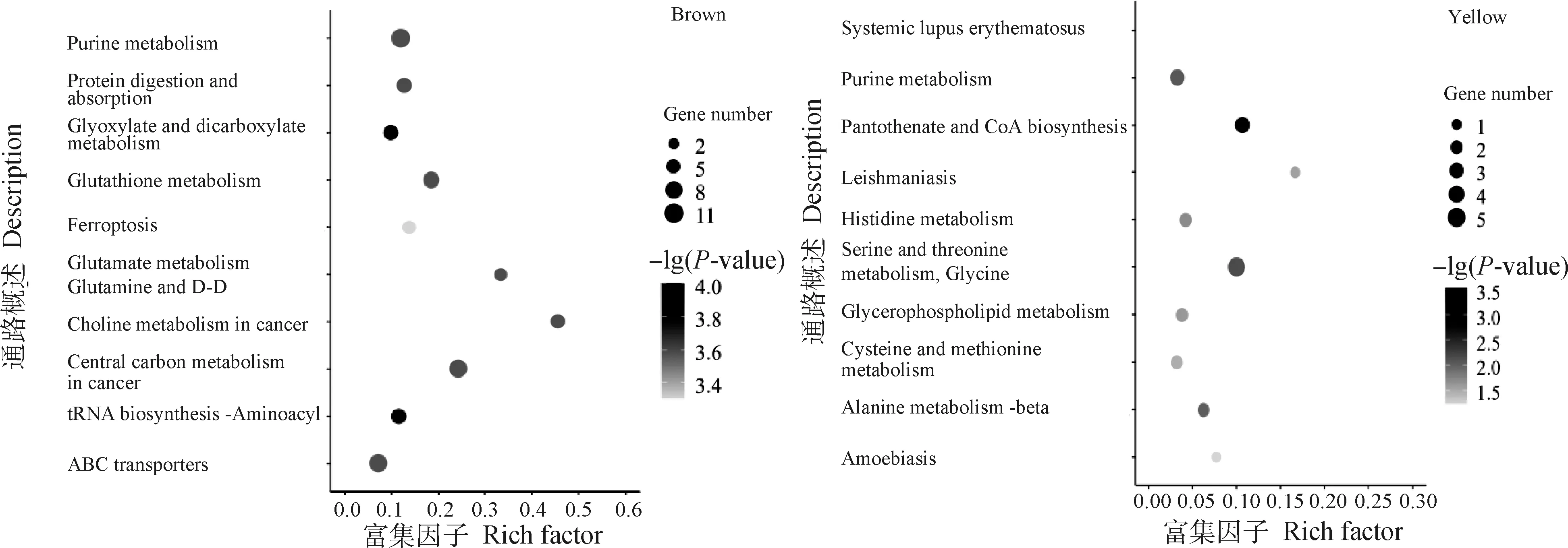
圖8 Brown、Yellow模塊代謝物Top10 KEGG富集散點圖
2.8 聯合KEGG通路分析
本研究對與IMF和剪切力相關模塊內的基因及代謝物進行KEGG通路共富集分析,基因模塊包括Darkgreen、Greenyellow及Daekgrey,共540個基因,代謝物模塊包括Brown、Lightcyan、Yellow、Darkgreen及Darkred,共247個代謝物。結果發現共有81個共富集通路,其中8條(丙氨酸、天冬氨酸和谷氨酸代謝、GABA能突觸、谷氨酸能突觸、精氨酸和脯氨酸代謝、苯丙胺成癮、近端小管碳酸氫鹽回收、β-丙氨酸代謝以及PPAR信號通路)在KEGG通路中顯著富集(P<0.1),其中丙氨酸、天冬氨酸和谷氨酸代謝、精氨酸和脯氨酸代謝、β-丙氨酸代謝以及PPAR信號通路可能與驢肉的嫩度相關(表2)。
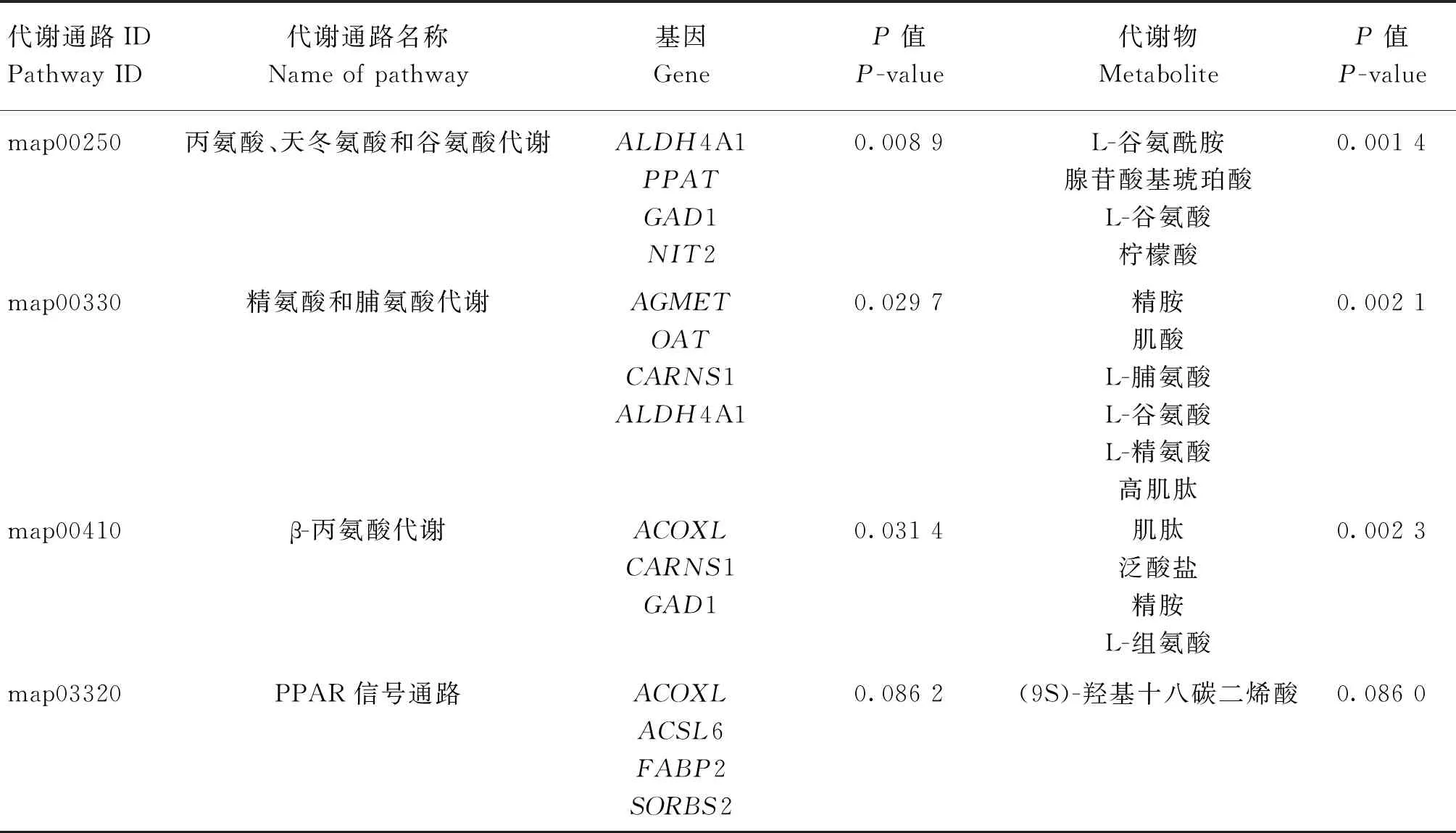
表2 模塊基因和代謝物聯合KEGG功能富集分析部分通路
2.9 模塊基因、代謝物相關性以及網絡圖分析
對4條KEGG共表達通路(丙氨酸、天冬氨酸和谷氨酸代謝、PPAR信號通路、β-丙氨酸代謝以及氨酸和脯氨酸代謝)中的基因與代謝物進行相關性分析,將Pearson相關系數絕對值大于0.5的基因與代謝物作為研究重點,發現4條共富集通路中均存在相關系數絕對值大于0.5的基因與代謝物,且各通路間具有連接性,圖9展示了共富集通路中基因與代謝物相關網絡。在丙氨酸、天冬氨酸和谷氨酸代謝通路中,腺苷酸基琥珀酸與PPAT、GAD1、NIT2基因正相關性強(圖9A)。在精氨酸和脯氨酸代謝通路中,AGMAT基因與代謝物L-脯氨酸、L-谷氨酸以及高肌肽負相關,與肌酸正相關;CARNS1基因與代謝物肌酸正相關,與高肌肽負相關(圖9B)。在β-丙氨酸代謝通路中,代謝物泛酸與基因CARNS1、ACOXL負相關,與GAD1正相關;肌肽與CARNS1、ACOXL正相關,與GAD1負相關(圖9C)。在PPAR信號通路中,代謝物(9S)-羥基十八碳二烯酸與SORBS2、ACOXL基因負相關(圖9D)。
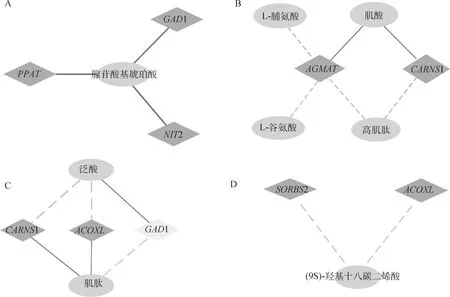
橢圓代表代謝物,菱形代基因,實線代表正相關,虛線代表負相關
3 討 論
嫩度評價指標直接受剪切力的數據結果影響,IMF也是影響嫩度的重要因素。本研究以剪切力和IMF為表型,通過WGCNA技術篩選與嫩度相關的候選基因,共鑒定到3個與IMF和剪切力相關的模塊以及各模塊中的相關基因。3個模塊均與IMF正相關,Darkgreen模塊與剪切力負相關,符合IMF與剪切力負相關特性。Greenyellow和Darkgrey模塊內的GO和KEGG均是與IMF相關的過程及通路,例如脂質氧化、脂肪酸生物合成過程、花生四烯酸代謝等,而Darkgreen模塊在GO功能富集分析上主要涉及肌肉發育及大分子降解、鈣離子結合等與肌肉及鈣調控相關的過程,KEGG功能富集分析則涉及到脂肪細胞因子信號通路、FoxO信號通路、MAPK信號通路等與IMF相關的信號轉導。
通過WGCNA篩選與肉質嫩度相關的代謝物信息和關鍵模塊,鑒定篩選到Brown、Yellow、Darkgreen、Lightcyan以及Darkred 5個關鍵模塊都與剪切力顯著相關,其中的Brown、Yellow、Darkgreen模塊與IMF顯著相關。但由于模塊中的大量代謝物無法進行識別,因此只對識別出相對較多代謝物的Brown和Yellow模塊進行后續分析。Brown模塊內的代謝物主要有丙氨酰-賴氨酸、L-酪氨酸和腺嘌呤等。肉類的嘌呤含量相對較高,肉類攝入量增加與痛風風險增加有關,Huang等[17]在研究中發現,豬肉中的次黃嘌呤和腺嘌呤(AH)的結合含量約占總嘌呤量的90%,AH含量與4種感官特征呈負相關,包括嫩度、多汁性、油性和總體喜好程度,降低肉中兩種尿酸嘌呤堿基的水平不僅有利于肉的營養價值,還能提高肉的感官接受度。Yellow模塊內的代謝物主要有N-乙酰-L-組氨酸、肌肽以及尿烷酸等。動物死后肉質嫩化的主要原因是由于肉中較大蛋白質結構的蛋白質水解,因此肉中的氨基酸量可能用于指示蛋白質的水解和肉質嫩化的結果。Straadt等[18]在5種不同雜交豬種的代謝組學技術與豬肉感官知覺關系的研究中發現,丙氨酸、肌肽、酪氨酸等7種氨基酸含量與肉嫩度呈正相關。
通過WGCNA篩選的候選基因及代謝物結果與KEGG功能富集分析,發現了81條通路共富集,其中天冬氨酸、丙氨酸和谷氨酸代謝、PPAR信號通路、β-丙氨酸代謝以及精氨酸和脯氨酸代謝顯著富集且與廣靈驢肉質嫩度相關,進行Pearson相關性分析后發現在4個通路中均有相關的基因及代謝物。Antonelo等[19]研究了與牛肉嫩度相關代謝物及代謝途徑,發現高嫩度牛肉代謝物乙酰肉堿、腺嘌呤、β-丙氨酸、纈氨酸水平顯著高于低嫩度牛肉且與剪切力負相關,表示這些代謝物可能影響牛肉嫩度。在候選通路中,PPAR信號通路與哺乳動物的肉質顯著相關,Fang等[20]在豬PPAR信號通路中挖掘了77個潛在功能性單核苷酸多態性(single nucleotide polymorphism, SNP),其中有13個與豬肉品質顯著相關的標記SNPs,它們對脂肪含量、肉色、背膘等性狀有顯著影響。PPAR是核受體超家族成員,充當配體誘導的轉錄因子,在葡萄糖和脂質代謝中起著至關重要的作用[21],而肌內脂肪沉積直接影響肉質嫩度,因此可以推測,PPAR信號通路可介導脂肪代謝相關途徑從而間接影響肉質剪切力。
在丙氨酸、天冬氨酸和谷氨酸代謝通路中,腺苷酸基琥珀酸(adenylosuccinate, S-AMP)在相關性分子中與基因GAD1、NIT2、PPAT的表達水平正相關。腺苷酸基琥珀酸是嘌呤代謝中合成AMP的前體物質,對β細胞功能障礙的逆轉有重要作用[22]。有研究發現,S-AMP與AMPKγ亞基通過形成復合體增強AMPK磷酸化水平,并提高甘油三酯脂肪酶和乙酰輔酶A羧化酶的表達量和磷酸化[23]。谷氨酸脫羧酶(glutamic acid decarboxylase, GAD),是一種可催化谷氨酸脫羧轉化成γ-氨基丁酸的脫羧酶,其廣泛表達于脊椎動物中樞神經的神經元中,同時GAD還具有刺激胰島素等激素分泌的作用,可增強動物食欲,提高動物機體的代謝率,促進動物生長[24]。GAD受GAD1和GAD2基因相互調控,現已發現在小鼠中GAD1是胰島細胞表達占絕對優勢的結構形式,這跟γ-氨基丁酸可與神經肽Y相互作用從而刺激食欲有關[25],然而目前還并未發現其對肉質影響的相關研究。同樣地,硝化酶樣蛋白2(nitrilase-like protein 2, NIT2)屬于硝化酶超家族成員之一,Krasnikov等[26]通過研究確定NIT2為ω-酰胺酶,可催化谷氨酰胺和天冬氨酸轉氨生成α-酮戊二酸和α-酮琥珀酸,谷氨酰胺轉氨可以挽救非特異性轉氨酶所產生必需氨基酸的α-酮酸。Wu等[27]利用基因組關聯分析研究發現,PPAT基因在肝中的表達水平與雞腹脂率、腹脂重顯著相關。另有研究表明,在牛肉屠宰老化過程中,谷氨酰胺和丙氨酸主要集中在更嫩的牛肉中[28]。在精氨酸和脯氨酸代謝通路中,AGMAT基因與代謝物L-脯氨酸、L-谷氨酸和高肌肽負相關,與肌酸正相關;CARNS1基因結果相反。羥脯氨酸作為膠原蛋白降解的最后一步,被用作判定膠原蛋白分解代謝的特定指標[29]。在雞屠宰后,肉質老化期間膠原溶解度、游離羥脯氨酸和蛋白水解率均增加,剪切力逐漸降低[30]。脯氨酸和谷氨酸與韓國黃牛肉質嫩度、多汁性和整體可接受性顯著正相關[31]。肌酸影響著肌肉的能量代謝,補充肌酸能提升肌肉性能[32]。在此通路中L-脯氨酸、L-谷氨酸、高肌肽和肌酸可能與AGMAT、CARNS1相互作用進而影響驢肉嫩度。
在β-丙氨酸代謝通路中,代謝物泛酸與基因CARNS1、ACOXL負相關,與GAD1正相關;肌肽與CARNS1、ACOXL正相關,與GAD1負相關。肌肽在肌肽合成酶(CARNS)作用下,將β-丙氨酸和L-組氨酸連接起來。β-丙氨酸是形成肌肽的限速因子,給育肥豬喂食β-丙氨酸能提高肌肉肌肽水平和運動能力[33]。肌肽參與調控機體內的抗氧化系統,有效喂食肌肽可提高豬的抗氧化能力和肉質[34]。持續熱應激破壞豬背最長肌促氧化/抗氧化平衡,肌肽含量和CARNS1 mRNA表達在30 ℃下3周后顯著降低[35]。肌酸和肌肽在安格斯牛中具有中等遺傳性,肌肽與牛肉整體嫩度呈負相關,風味研究發現魚腥味與肌酐和肌肽顯著負相關[36]。研究發現,輔酶A前體泛酸可以提高細胞的呼吸強度,推測泛酸可能通過提高線粒體輔酶A的含量進而增加了ATP及谷胱甘肽的含量[37]。ACOXL即酰基輔酶A氧化酶樣蛋白,具有酰基輔酶A脫氫酶活性,是過氧化物酶體脂肪酸氧化的主要限速酶。李強[38]通過全基因組關聯分析發現,ACOXLc.8C/T與大白豬第十肋骨處背膘厚顯著關聯。在PPAR信號通路中代謝物(9S)-羥基十八碳二烯酸與基因SORBS2和ACOXL負相關。SORBS2即SH3結構域包含蛋白2,是一種與Abl/Arg非受體酪氨酸激酶途徑相關的支架蛋白,已知和肌動蛋白及其他幾種不同類型的骨架蛋白相互作用。SORBS2可誘導原代人成纖維細胞衰老[39]。SORBS2可能通過影響細胞連接以及成纖維細胞衰老進而影響肉質嫩度。
4 結 論
本研究基于WGCNA技術篩選與嫩度相關的候選基因及代謝物,之后進行KEGG功能富集聯合分析。發現嫩度候選基因及代謝物共富集在丙氨酸、天冬氨酸和谷氨酸代謝、精氨酸和脯氨酸代謝、β-丙氨酸代謝以及PPAR信號通路上,說明這些通路可能對調控驢肉嫩度有重要作用。各通路上均存在Pearson相關系數大于0.5的基因及代謝物,GAD1、PPAT、NIT2、AGMAT、CARNS1、ACOXL、SORBS2以及腺苷酸基琥珀酸、L-脯氨酸、L-谷氨酸、肌酸、高肌肽、肌肽、泛酸、(9S)-羥基十八碳二烯酸可能是影響驢肉嫩度的候選基因及代謝物。本研究基于WGCNA技術以及KEGG共富集分析篩選了影響嫩度的候選基因及代謝物,為解析驢肉嫩度調控機制奠定了基礎,為驢肉分子改良提供了理論依據。
