IDH1調(diào)控干擾素通路相關(guān)基因IFIT3的表達(dá)
2021-07-19 06:52:34唐婉軍彭小忠
基礎(chǔ)醫(yī)學(xué)與臨床 2021年7期
唐婉軍,彭小忠,2*,韓 為*
(1.中國(guó)醫(yī)學(xué)科學(xué)院基礎(chǔ)醫(yī)學(xué)研究所 北京協(xié)和醫(yī)學(xué)院基礎(chǔ)學(xué)院 生物化學(xué)與分子生物學(xué)系 醫(yī)學(xué)分子生物學(xué) 國(guó)家重點(diǎn)實(shí)驗(yàn)室 醫(yī)學(xué)靈長(zhǎng)類(lèi)研究中心 神經(jīng)科學(xué)中心, 北京 100005; 2.中國(guó)醫(yī)學(xué)科學(xué)院 醫(yī)學(xué)生物學(xué)研究所,云南 昆明 650118)
膠質(zhì)瘤(glioma)是最常見(jiàn)的原發(fā)性中樞神經(jīng)系統(tǒng)腫瘤。在膠質(zhì)瘤中,異檸檬酸脫氫酶1(isocitrate dehydrogenase 1, IDH1)是最重要的分子標(biāo)志物之一。膠質(zhì)母細(xì)胞瘤(glioblastoma multiforme,GBM)屬于惡性程度最高的膠質(zhì)瘤,其中IDH1野生型較為常見(jiàn),而IDH1突變型僅占10%[1]。與IDH1突變型患者相比,IDH1野生型患者預(yù)后較差,放射化學(xué)藥物治療(簡(jiǎn)稱(chēng)放化療)效果不佳[1-2]。以往針對(duì)IDH1的研究多關(guān)注該催化酶發(fā)生突變后在代謝和基因調(diào)控中的作用[3],而野生型IDH1表達(dá)失調(diào)對(duì)基因表達(dá)的影響尚不清楚。IDH1在膠質(zhì)瘤中的關(guān)鍵地位決定了對(duì)其進(jìn)行廣泛而深入研究的必要性。
RNA結(jié)合蛋白(RNA binding proteins, RBPs)通過(guò)RNA結(jié)合結(jié)構(gòu)域與目的RNA相結(jié)合[4],在基因表達(dá)調(diào)控中發(fā)揮重要作用。RBPs廣泛參與多種細(xì)胞生物過(guò)程,如中間代謝、抗病毒反應(yīng)、信號(hào)傳導(dǎo)和蛋白質(zhì)代謝等,并且RBP-RNA相互作用網(wǎng)絡(luò)的失調(diào)在腫瘤和神經(jīng)退行性疾病中可作為一種驅(qū)動(dòng)因素[5]。本研究旨在探究IDH1表達(dá)失調(diào)能否影響RBPs的表達(dá),為進(jìn)一步理解IDH1在膠質(zhì)瘤中的關(guān)鍵作用提供更廣闊的思路。
1 材料與方法
1.1 材料
1.1.1 細(xì)胞系:人神經(jīng)膠質(zhì)瘤細(xì)胞系LN229(ATCC)。
1.1.2 主要試劑:Trizol (Ambion 公司);RNA純化試劑盒(Qiagen公司);反轉(zhuǎn)錄試劑盒(Vazyme公司);SYBR (TaKaRa公司);IDH1抗體(Abcam公司),IDH1R132H(Dianova公司),β-actin 抗體(Sigma-Aldrich 公司),慢病毒及感染試劑(上海吉?jiǎng)P基因有限公司)。
1.2 方法
1.2.1 慢病毒感染構(gòu)建IDH1敲低、過(guò)表達(dá)和R132H突變的細(xì)胞株: 慢病毒載體構(gòu)建和包裝均委托上海吉?jiǎng)P基因有限公司。慢病毒感染細(xì)胞12~16 h后更換新鮮培養(yǎng)基, 后檢測(cè)熒光蛋白表達(dá)情況。然后使用病毒載體攜帶其抗性基因的抗生素篩選細(xì)胞72 h左右,此時(shí)空白對(duì)照組細(xì)胞應(yīng)全部死亡。提取各組細(xì)胞蛋白和RNA檢測(cè)IDH1的表達(dá)水平以確認(rèn)慢病毒感染效果。
1.2.2 RT-qPCR檢測(cè)目的基因的表達(dá):收取細(xì)胞后利用RNA純化試劑盒提取細(xì)胞總RNA。使用反轉(zhuǎn)錄試劑盒將RNA反轉(zhuǎn)錄為cDNA。使用SYBR預(yù)混試劑及定量PCR儀進(jìn)行實(shí)時(shí)熒光定量。每個(gè)樣品設(shè)置3~4個(gè)復(fù)孔,以GAPDH作為內(nèi)參計(jì)算樣品的相對(duì)表達(dá)量。針對(duì)9個(gè)候選基因分別設(shè)計(jì)引物,并確認(rèn)引物特異性。進(jìn)行RT-qPCR檢測(cè)候選基因在IDH1敲低、過(guò)表達(dá)和R132H突變的細(xì)胞株中的表達(dá)水平。
1.2.3 RNA-seq:將IDH1敲低組和對(duì)照組LN229細(xì)胞在6 cm細(xì)胞培養(yǎng)皿中培養(yǎng),每組設(shè)置3個(gè)重復(fù)。待細(xì)胞匯合至70%左右,每個(gè)6 cm皿加入1 mL Trizol 收取細(xì)胞,送北京博奧晶典生物技術(shù)有限公司進(jìn)行建庫(kù)及高通量測(cè)序。
1.2.4 數(shù)據(jù)分析:利用limma進(jìn)行差異表達(dá)基因分析,差異基因篩選標(biāo)準(zhǔn)為:|log2FC|≥1且P<0.05。利用KOBAS軟件進(jìn)行差異表達(dá)基因富集通路分析。比對(duì)RBPs數(shù)據(jù)庫(kù)[6]選出候選RBPs后,使用String數(shù)據(jù)庫(kù)(http://string-db.org/)對(duì)候選RBPs間的相互作用進(jìn)行預(yù)測(cè)分析。
1.3 統(tǒng)計(jì)學(xué)分析
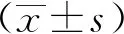
2 結(jié)果
2.1 IDH1敲低組和對(duì)照組LN229細(xì)胞株的構(gòu)建及RNA-seq樣品相關(guān)性分析
RT-qPCR和Western blot結(jié)果顯示:與對(duì)照組(sh-control)相比,IDH1敲低組(shIDH1-1和shIDH1-2)的IDH1RNA(圖1A)和蛋白(圖1B)水平顯著降低,表明成功構(gòu)建IDH1敲低的細(xì)胞株。同時(shí),用于轉(zhuǎn)錄組測(cè)序的IDH1敲低組和對(duì)照組細(xì)胞樣品,3個(gè)生物學(xué)重復(fù)樣本間相關(guān)性系數(shù)r≥0.96(圖1C)。

A, B.IDH1 expression was validated by Western blot and n=3),*P<0.001 compared with the control (sh-control) group; C.Heat map demonstrated the correlation between samples(n=3)
2.2 RNA-seq分析結(jié)果
聚類(lèi)分析顯示樣本被分為對(duì)照組和IDH1敲低組兩個(gè)群體,每個(gè)群體內(nèi)3個(gè)重復(fù)樣本間具有相似的表達(dá)特征,并且基因表達(dá)成簇分布(圖2A)。與對(duì)照組細(xì)胞相比,IDH1敲低組共檢測(cè)到259個(gè)差異表達(dá)基因(differentially expressed genes,DEGs)(圖2B)。對(duì)這些差異表達(dá)基因進(jìn)行通路富集分析后發(fā)現(xiàn),差異表達(dá)基因顯著富集在干擾素等相關(guān)通路(圖2C)。

A.heat map for expression profile in the IDH1 knockdown and the control group; B.identification of DEGs after IDH1 knockdown, red dots represent upregulated genes and blue dots represent downregulated genes,|log2fold-change|≥1 and P<0.05; C.pathway enrichment of DEGs after IDH1 knockdown
2.3 候選RBPs篩選及其蛋白間相互作用分析
通過(guò)比對(duì)RBP數(shù)據(jù)庫(kù),將差異表達(dá)基因與RBP數(shù)據(jù)庫(kù)取交集后得到14個(gè)差異表達(dá)RBPs(圖3A)。進(jìn)一步對(duì)其進(jìn)行蛋白間相互作用分析(圖3B)后,挑選出9個(gè)樞紐RBPs(表1)做進(jìn)一步驗(yàn)證。與對(duì)照組相比,這9個(gè)候選RBPs的RNA水平在IDH1敲低的LN229細(xì)胞株中顯著升高(P<0.001)(圖3C)。

表1 IDH1調(diào)控的候選RBPs列表

A.screening IDH1-regulated RBPs; B.protein-protein interaction analysis of candidate RBPs; C.validation of chosen RBPs by RT-qPCR in IDH1 knockdown cell line LN229; *P<0.001 compared with the control (sh-control) group
2.4 過(guò)表達(dá)IDH1R132H突變型及野生型的LN229細(xì)胞株構(gòu)建和候選RBPs表達(dá)驗(yàn)證
RT-qPCR和Western blot結(jié)果顯示:與對(duì)照組(IDH1WT)相比,IDH1過(guò)表達(dá)組(IDH1OE)的IDH1RNA(圖4A)和蛋白(圖4B)表達(dá)顯著升高,表明成功構(gòu)建IDH1OE的細(xì)胞株。用IDH1R132H特異性抗體檢測(cè)結(jié)果顯示,成功構(gòu)建IDH1R132H細(xì)胞株(圖4B)。與IDH1WT相比,在IDH1R132H細(xì)胞株中,DDX58、IFIH1、IFIT2和OAS2的RNA表達(dá)顯著升高(P<0.05)(圖4C)。而在IDH1OE細(xì)胞株中,IFIT3的RNA表達(dá)水平顯著降低(P<0.01)(圖4C)。

A, B.IDH1 expression was demonstrated by Western blot and RT-qPCR, *P<0.05, **P<0.01, ***P<0.001 compared with the control (IDH1WT) group; C.identification of the expression levels of candidate RBPs in IDH1R132H and IDH1OE cells
3 討論
以往研究表明,IDH1高表達(dá)及IDH1突變均可促進(jìn)膠質(zhì)瘤的發(fā)生與發(fā)展。IDH1高表達(dá)通過(guò)促進(jìn)脂質(zhì)合成等代謝途徑發(fā)揮促癌作用,并且與病人的不良預(yù)后顯著相關(guān)[7]。而IDH1突變通過(guò)生成腫瘤代謝物羥基戊二酸(α-hydroxyglutaric acid, 2-HG)影響膠質(zhì)瘤進(jìn)展[8]。本研究發(fā)現(xiàn),在膠質(zhì)瘤細(xì)胞株LN229中敲低IDH1可導(dǎo)致多種基因的表達(dá)顯著改變,進(jìn)行通路富集分析顯示這些基因顯著富集在干擾素信號(hào)通路和免疫系統(tǒng)相關(guān)通路等。這一結(jié)果提示,IDH1可能會(huì)調(diào)控免疫應(yīng)答相關(guān)基因的表達(dá)。
進(jìn)一步分析發(fā)現(xiàn),IDH1敲低后的差異表達(dá)基因包括14個(gè)編碼RBP的基因,其中9個(gè)RBPs間具有密切的相互作用,且均參與干擾素相關(guān)通路。與對(duì)照組相比,在IDH1敲低的膠質(zhì)瘤細(xì)胞系LN229中,這9個(gè)RBPs表達(dá)水平均顯著上調(diào);IDH1過(guò)表達(dá)時(shí),IFIT3表達(dá)顯著下降;而IDH1突變時(shí),IFIT3表達(dá)未檢測(cè)到顯著變化。以上結(jié)果提示,在膠質(zhì)瘤細(xì)胞系LN229中,IFIT3可能受到IDH1的調(diào)控。
IFIT3屬于三角形四肽重復(fù)干擾素誘導(dǎo)蛋白(interferon-induced protein with tetratricopeptide repeats, IFIT)家族。該家族成員包括IFIT1、IFIT2、IFIT3和IFIT5,它們均具有多個(gè)TRP(tetratricopeptide repeat)模體,通過(guò)識(shí)別病毒RNA發(fā)揮抗病毒功能[9]。除此之外,IFIT蛋白家族還參與一系列生物學(xué)過(guò)程,例如細(xì)胞增殖、 遷移和雙鏈RNA信號(hào)通路等[10]。多項(xiàng)研究表明,IFITs家族成員在腫瘤惡性進(jìn)展中發(fā)揮重要作用:在膠質(zhì)母細(xì)胞瘤中,IFIT1可誘導(dǎo)細(xì)胞凋亡,并且IFIT1高表達(dá)的患者預(yù)后較好[11];IFIT2通過(guò)抑制上皮間質(zhì)轉(zhuǎn)化(epithelial-mesenchymal transition, EMT)降低腫瘤細(xì)胞遷移和侵襲能力,從而影響口腔鱗狀細(xì)胞癌進(jìn)展[12];在人單核淋巴瘤細(xì)胞系U937中,IFIT3通過(guò)下調(diào)c-Myc,增強(qiáng)細(xì)胞周期負(fù)性調(diào)控因子p27和p21的表達(dá),抑制腫瘤細(xì)胞增殖[13];IFIT5通過(guò)調(diào)控miRNA的代謝促進(jìn)腎細(xì)胞癌進(jìn)展[14]。目前為止,IFIT3在膠質(zhì)瘤發(fā)生發(fā)展中的功能未見(jiàn)報(bào)道,還需進(jìn)一步研究。
綜上所述,本研究在人膠質(zhì)瘤細(xì)胞系LN229中敲低IDH1后,通過(guò)RNA-seq檢測(cè)IDH1調(diào)控的基因,并且對(duì)干擾素通路相關(guān)的RBPs編碼基因進(jìn)行驗(yàn)證,最終挑選出IDH1調(diào)控的候選基因IFIT3。這為進(jìn)一步研究IDH1在膠質(zhì)瘤發(fā)生發(fā)展中的關(guān)鍵作用提供了新的線索,然而IDH1對(duì)IFIT3的具體調(diào)控機(jī)制還需要深入探索。
