降香黃檀葉綠體基因組密碼子偏好性分析
2021-06-29 20:52:38原曉龍李云琴張勁峰王毅
廣西植物 2021年4期
關(guān)鍵詞:選擇
原曉龍 李云琴 張勁峰 王毅
摘?要:?為了解降香黃檀葉綠體基因組密碼子使用模式,該文利用Codon W 1.4.2和在線軟件CUSP對降香黃檀葉綠體基因組中的52條基因編碼序列密碼子進(jìn)行中性繪圖、ENC-plot和PR2-plot分析。結(jié)果表明:降香黃檀葉綠體基因組密碼子的3個位置上GC含量依次為GC1(46.01%)>GC2(38.98%)>GC3(27.80%)。有效密碼子數(shù)(ENC)范圍為37.66~54.43,及ENC值>45的有37個;RSCU>1的密碼子有29個,其中16個以U結(jié)尾、12個以A結(jié)尾。這些說明其偏好以A和U結(jié)尾,且偏性較弱。中性繪圖分析顯示GC12與GC3的相關(guān)系數(shù)為0.250,相關(guān)性不顯著,回歸系數(shù)為0.394;ENC-plot分析顯示ENC比值位于-0.05~0.05區(qū)間外的基因有38個;PR2-plot分析說明在堿基的使用頻率方面,U>A、G>C,說明降香黃檀葉綠體基因組密碼子偏好性主要受選擇的影響;19個密碼子被確定為最優(yōu)密碼子。該研究為降香黃檀葉綠體基因工程、遺傳多樣性分析等研究提供了科學(xué)參考依據(jù)。
關(guān)鍵詞: 降香黃檀, 葉綠體基因組, 密碼子偏好性, 選擇
中圖分類號:?Q755
文獻(xiàn)標(biāo)識碼:?A
文章編號:?1000-3142(2021)04-0622-09
Abstract:?To comprehend the codon usage pattern of chloroplast genome in Dalbergia odorifera, the 52 coding DNA sequences were analyzed to obtain to the results of neutrality plot, ENC-plot and PR2-plot analysis using Codon W 1.4.2 and online software CUSP in the present study. The results were as follows:?The GC content in the three positions of codons from the chloroplast genome of D. odorifera was GC1 (46.01%)>GC2 (38.98%)>GC3 (27.80%) successively. The range of effective number codon was from 37.66 to 54.43, and there were 37 genes when ENC value was greater than 45, and there were 29 genes when RSCU value was greater than 1, including 16 genes ending with U and 12 genes ending with A. These suggest that the codons prefer ends with A and U, and have a weak bias. The neutrality plot showed that there was no significant correlation between GC3 and GC12, the correlation coefficient was 0.250, and the regression coefficient was 0.394; ENC-plot analysis revealed that there were 38 genes which ENC ratio located beyond the section from -0.05 to 0.05; PR2-plot analysis showed that U>A and G>C in the base usage frequency. These all illustrated that the codon usage bias in the chloroplast genome of D. odorifera was mainly affected by mutation; 19 codons were identified as the optimal codon. The present study could be useful in the chloroplast genetic engineering and genetic diversity analysis of D. odorifera.
Key words: Dalbergia odorifera, chloroplast genome, codon usage bias, selection
在生物體傳遞遺傳信息的過程中,作為聯(lián)結(jié)核酸和蛋白質(zhì)的密碼子扮演著重要的角色(Zhang et al., 2019),構(gòu)成基因組的4種核苷酸可形成64種密碼子,各密碼子與氨基酸相對應(yīng),除甲硫氨酸和色氨酸外,其余18種氨基酸均有2~6個密碼子,這些編碼同一氨基酸的不同密碼子被稱為同義密碼子(synonymous codon)(Duret, 2002)。在翻譯過程中,每個氨基酸相對應(yīng)同義密碼子的使用頻率存在差異,即有的同義密碼子使用頻率高于其他同義密碼子,這種現(xiàn)象被稱為密碼子偏好性(codon usage bias)(Romero et al., 2000)。密碼子偏好性廣泛存在于不同生物中,是物種在長期進(jìn)化過程中受環(huán)境選擇、堿基突變、基因漂變等因素共同作用,還受到基因組大小、tRNA豐度和基因表達(dá)水平等的影響(Romero et al., 2000; Duret, 2000; Xu et al., 2011)。密碼子偏好性通過對基因翻譯準(zhǔn)確性和效率的調(diào)節(jié)影響基因的表達(dá)水平(邢朝斌等, 2013),葉綠體具有基因組小、基因拷貝數(shù)多等特點(diǎn)(Wright, 1990; 牛元等, 2018);根據(jù)最優(yōu)密碼子設(shè)計葉綠體基因表達(dá)載體,可迅速提高葉綠體基因組中基因表達(dá)量,利用目前已知的密碼子使用模式推斷未知基因的表達(dá),或預(yù)測某些未知基因的功能(Wu et al, 2007);同時親緣關(guān)系較近的物種具相近的密碼子使用模式(楊國鋒等, 2015)。因此研究葉綠體基因組密碼子的使用模式對探討物種進(jìn)化、提高外源基因表達(dá)量等具有重要意義。
降香黃檀(Dalbergia odorifera)是蝶形花科(Papilionaceae)黃檀屬(Dalbergia)常綠半落葉喬木,其心材具極高的經(jīng)濟(jì)和藥用價值(梁遠(yuǎn)楠等, 2019; 王玥琳等, 2019; 張麗佳等, 2019),本研究已完成對降香黃檀葉綠體基因組的高通量測序,通過分析降香黃檀葉綠體基因組蛋白編碼區(qū)(coding DNA sequence, CDS)序列的堿基組成、中性繪圖、ENC-plot及PR2-plot等方法推斷影響降香黃檀葉綠體密碼子偏好性的主要因素,確定降香黃檀葉綠體基因組的最優(yōu)密碼子。本研究通過對降香黃檀葉綠體基因組密碼子使用模式和影響密碼子使用偏性的因素進(jìn)行分析,確定降香黃檀葉綠體基因組的密碼子偏好性及最優(yōu)密碼子,以期為降香黃檀葉綠體基因組的應(yīng)用和研究提供科學(xué)的參考依據(jù)。
1?材料與方法
1.1 材料
降香黃檀葉片采自云南省林業(yè)科學(xué)院熱帶林業(yè)研究所。將采集的新鮮葉片樣品保存在干冰環(huán)境中送至生工生物工程(上海)股份有限公司進(jìn)行葉綠體基因組的測序,并將結(jié)果提交至NCBI(登錄號:KC427274),通過GenBank Feature Extractor(http://www.bioinformatics.org/sms2/genbank_feat.html)、ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)、DNA man等軟件分析降香黃檀葉綠體基因組,根據(jù)各基因的注釋結(jié)果獲得83條CDS序列,剔除序列長度小于300 bp、重復(fù)基因和CDS內(nèi)部存在終止密碼子的序列后得到52條符合條件的CDS用于分析。
1.2 方法
1.2.1 密碼子組成分析?將52條符合條件的CDS 整合到一個.fasta文件中,應(yīng)用Codon W 1.4.2軟件分析獲得各CDS的ENC(effective number of codon, 有效密碼子數(shù))、同義密碼子相對使用度(RSCU),應(yīng)用在線軟件CUSP (http://emboss.toulouse.inra.fr/cgi-bin/emboss/cusp)分析獲得密碼子第1位、第2位、第3位堿基的GC含量(分別為GC1、GC2、GC3)及3位堿基的GC平均含量(GCall)等參數(shù),應(yīng)用SPSS和Excel等數(shù)理的統(tǒng)計分析軟件對結(jié)果進(jìn)行分析。
ENC是衡量同義密碼子使用偏度的重要指標(biāo),ENC的取值范圍為20~61,ENC值的大小能夠反映密碼子偏性的強(qiáng)弱。當(dāng)ENC值為20時,代表同義密碼子完全處于偏倚狀態(tài);當(dāng)ENC值為61時,代表同義密碼子完全沒有偏倚;ENC值從小到大表示偏倚性由強(qiáng)變?nèi)酰ǔ?梢訣NC值45作為區(qū)分偏倚性強(qiáng)弱的標(biāo)準(zhǔn)(秦政等, 2018)。RSCU是指某一密碼子實(shí)際使用頻率與無使用偏性時理論頻率的比值。無偏性時,RSCU為1;RSCU小于1則代表該密碼子的實(shí)際使用頻率低于其他同義密碼子的使用頻率,反之實(shí)際頻率高于其他同義密碼子的使用頻率(晁岳恩等, 2012)。
應(yīng)用SPSS軟件分析密碼子不同位置GC堿基組成GC1、GC2、GC3、GCall,密碼子數(shù)(N)與ENC等的相關(guān)關(guān)系,進(jìn)而判斷各因子對密碼子偏好性的影響。
1.2.2 中性繪圖分析?簡并密碼子第3位堿基通常所發(fā)生的為同義突變,而在密碼子第1位、第2位上發(fā)生的堿基突變通常會改變基因的功能或活性,即在不存在外界壓力時,密碼子3個位置的堿基含量組成應(yīng)該無差異;而在存在一定選擇壓力情況下時,密碼子3個位置上的堿基組成是存在差異的(Sueoka, 2001)。在以GC1和GC2的平均值GC12和GC3分為縱、橫坐標(biāo)的中性繪圖中,每一個散點(diǎn)代表一個基因。 如果中性圖中的所有基因均沿對角線分布,即GC12和GC3的變異基本一致,密碼子3個位置上的堿基組成無明顯差異,受選擇壓力較弱,而受突變影響較大;回歸系數(shù)(對角線斜率)是衡量中性程度的指標(biāo)之一,若回歸曲線斜率極小,GC12和GC3的變異的相關(guān)性同樣很低,說明影響密碼子偏好性的主要影響因素為選擇效應(yīng)(Sueoka, 2001)。同時通過分析密碼子不同位置堿基組成的相關(guān)性可分析密碼子偏好性受突變或選擇的影響,即當(dāng)GC12與GC3呈顯著相關(guān)時,說明密碼子3個位置的堿基組成無明顯差異,偏好性主要受突變影響; 當(dāng)GC12與GC3呈不顯著相關(guān)時,回歸系數(shù)趨近于0,說明密碼子的前兩位堿基與第3位堿基組成不同,基因組中的GC含量較為保守,密碼子的使用偏性主要受選擇影響(楊國鋒等, 2015)。
1.2.3 ENC-plot分析?ENC-plot分析可探討ENC和GC3S分布的關(guān)系,是一種通過對基因數(shù)據(jù)的密碼子偏好性情況的圖像可視化的有效方式,其中的標(biāo)準(zhǔn)曲線代表無選擇壓力存在時,密碼子偏好性完全由突變決定,即完全由核酸序列組成決定密碼子偏好性(Wright, 1990)。
ENC-plot繪圖分析含散點(diǎn)圖和標(biāo)準(zhǔn)曲線,散點(diǎn)圖則以ENC為縱坐標(biāo),GC3為橫坐標(biāo),標(biāo)準(zhǔn)曲線公式為ENC=2+GC3+29/[GC32+(1-GC3)2]。具體判斷標(biāo)準(zhǔn)為圖中散點(diǎn)與標(biāo)準(zhǔn)曲線的距離,散點(diǎn)與標(biāo)準(zhǔn)曲線的距離近則說明密碼子偏好性主要由堿基組成決定,受翻譯選擇的影響較為微弱;距離遠(yuǎn)則說明密碼子的ENC值偏低,與基因表達(dá)水平存在較強(qiáng)的顯著性相關(guān)關(guān)系,密碼子偏好性較強(qiáng)(尚明照等, 2011; 王鵬良等, 2018)。然而,ENC-plot繪圖分析不足以準(zhǔn)確區(qū)分中性突變和選擇壓力的影響程度,若當(dāng)選擇出現(xiàn)在密碼子第3位堿基上時,需要結(jié)合ENC比值頻數(shù)對差異進(jìn)行量化分析 (Sueoka, 2001)。
1.2.4 PR2-plot分析?分析各密碼子第3位上的A、T、C、G含量,以A3/(A3+T3)為縱坐標(biāo),以G3/(G3+C3)為橫坐標(biāo)進(jìn)行PR2偏倚分析(PR2-bias plot analysis)作圖,用平面圖顯示各基因的堿基組成,其中心點(diǎn)代表無偏性使用時的密碼子狀態(tài),即A=T且C=G,其余點(diǎn)與中心點(diǎn)的矢量距離則代表其偏倚程度和方向(楊國鋒等, 2015)。
1.2.5 最優(yōu)密碼子的確定?以降香黃檀葉綠體各基因的ENC作為偏好性參考標(biāo)準(zhǔn),從兩端各選擇10%的基因構(gòu)建高低偏性庫,將兩庫間ΔRSCU≥ 0.08的密碼子定為高表達(dá)優(yōu)越密碼子;將RSCU值大于1的密碼子確定為高頻密碼子(尚明照等, 2011)。將同時滿足高頻率密碼子和高表達(dá)優(yōu)越密碼子確定為最優(yōu)密碼子。
2?結(jié)果與分析
2.1 密碼子堿基組成
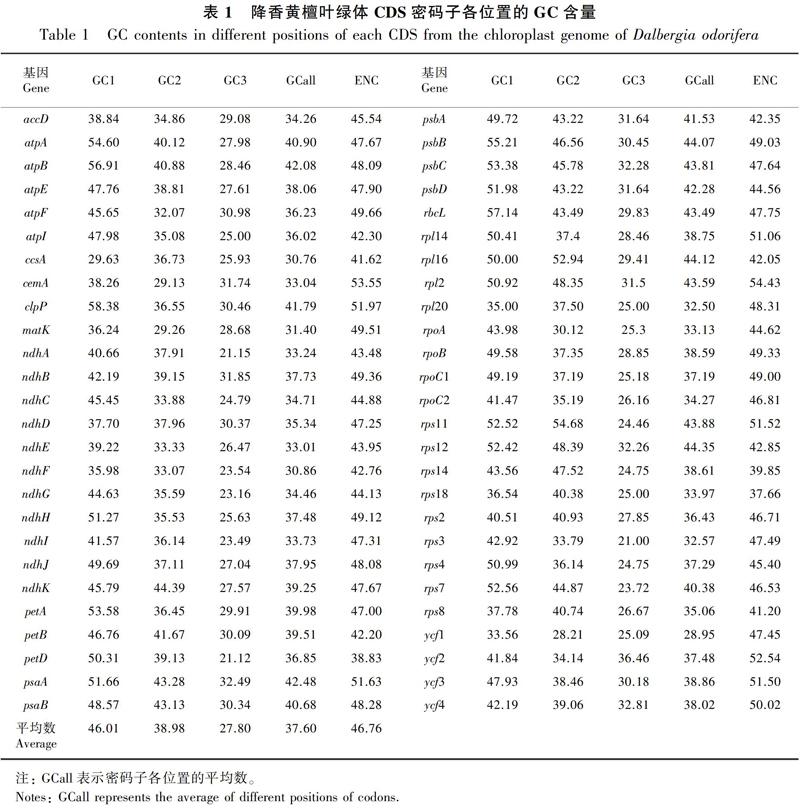
用在線軟件CUSP分析降香黃檀52條CDS的堿基組成,用Codon W 1.4.2分析其ENC值(表1),所有CDS密碼子的平均GC含量為37.60%,第1位GC含量為46.01%,第2位為38.98%,第3位為27.80%,GC含量在密碼子的不同位置分布頻率不同,由高到低依次為第1位>第2位>第3位,說明在降香黃檀中,葉綠體基因組密碼子末位堿基以A/U(T)為主,與植物葉綠體基因中A/U(T)含量較高的特征相符。表示偏倚強(qiáng)弱的ENC值的范圍為37.66~54.43,平均值為46.76;52個CDS的密碼子ENC值>45的有36個,說明其降香黃檀大部分基因編碼序列的密碼子偏性較弱。
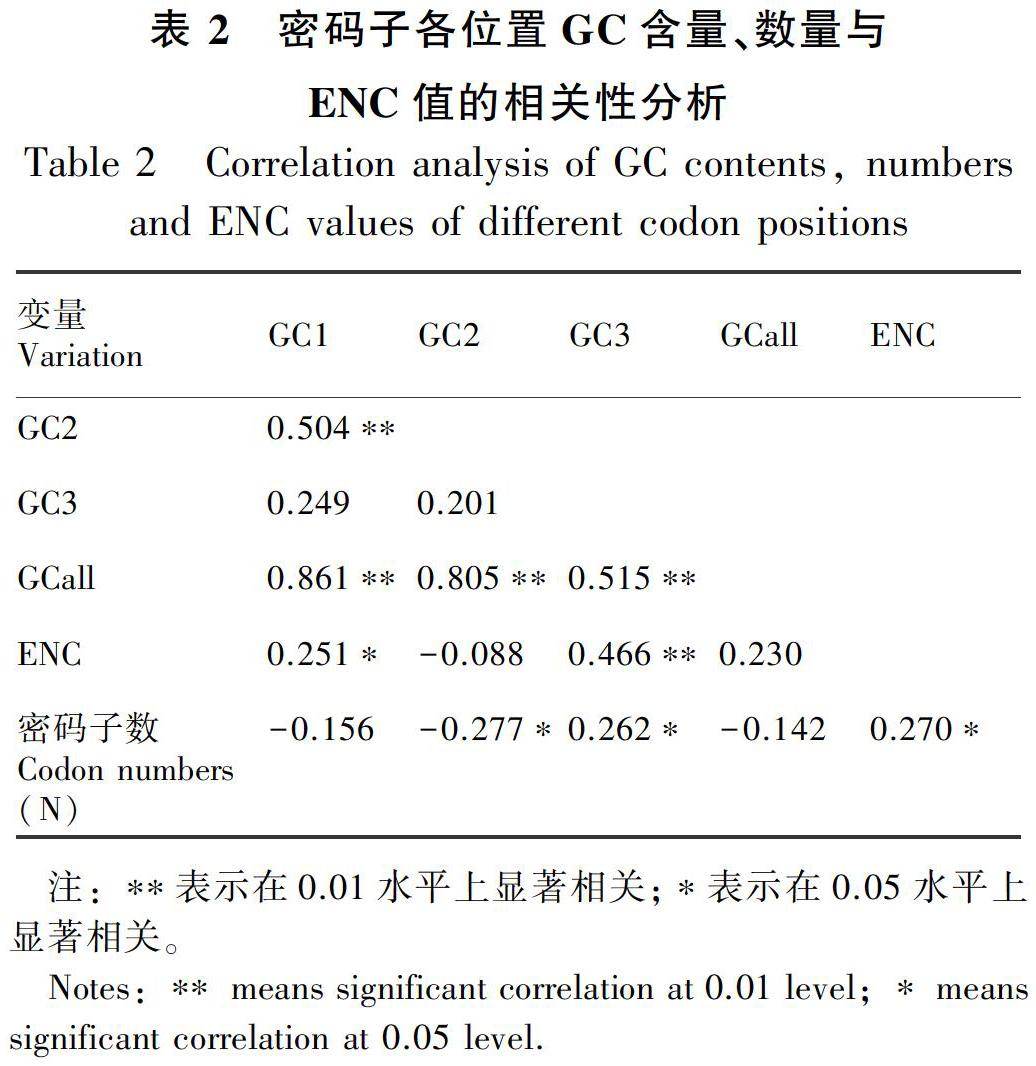
密碼子不同位置堿基的GC含量、密碼子數(shù)(N)與ENC值間的相關(guān)性分析(表2)顯示,GCall和GC1、GC2、GC3的相關(guān)性均達(dá)到極顯著水平,GC1和GC2達(dá)極顯著相關(guān), GC3與GC1、GC2均未達(dá)到顯著相關(guān),說明密碼子的第1位和第2位堿基組成相似,與第3位堿基組成存在差異。ENC與GC1呈顯著相關(guān),與GC2相關(guān)性不顯著,與GC3呈極顯著相關(guān),說明ENC與密碼子第3位堿基組成密切相關(guān)。ENC與密碼子數(shù)(N)呈顯著相關(guān),說明基因編碼序列長度對密碼子使用偏性具一定影響。
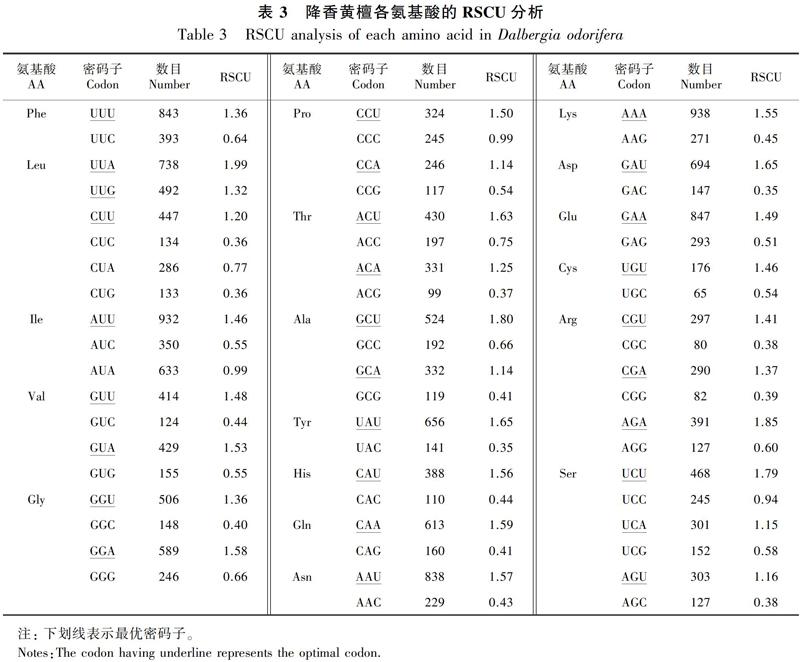
各編碼氨基酸的RSCU(表3)顯示,RSCU>1的密碼子以A和U結(jié)尾的頻率較高,其中16個以U結(jié)尾、12個以A結(jié)尾、1個以G結(jié)尾,說明降香黃檀葉綠體基因組偏愛以A和U結(jié)尾,而以C和G結(jié)尾的密碼子為非偏愛密碼子。
2.2 中性繪圖分析
降香黃檀葉綠體基因組各基因的中性繪圖分析(圖1),GC12的取值范圍稍大,在0.309~0.536之間,GC3的取值范圍很小,在0.210~0.365之間,且各基因均落在對角線上方;GC12與GC3的相關(guān)系數(shù)為0.249 6,相關(guān)性不顯著,且回歸系數(shù)(即趨勢線的斜率)為0.393 8,即在降香黃檀葉綠體基因組中性繪圖分析中,GC12和GC3兩個變量間的相關(guān)性很弱,說明密碼子第1位、第2位和第3位堿基組成存在差異,即降香黃檀葉綠體基因組中GC含量高度保守,且密碼子第3位的GC含量較低,其密碼子偏好性更多地受選擇的影響。
2.3 ENC-plot繪圖分析
ENC-plot繪圖分析是以ENC比值[(預(yù)期ENC值-實(shí)際ENC值)/預(yù)期ENC值]和ENC比值頻數(shù)來判斷影響基因的主要因素,與標(biāo)準(zhǔn)曲線的距離近的基因數(shù)量較多,則其偏好性主要受突變的影響;反之以選擇為主要影響因素。分析結(jié)果(圖2)顯示ENC比值頻數(shù)表(表4)中分布在-0.05~0.05區(qū)間的基因有14個,即有14個基因與預(yù)期ENC值較接近,而分布在-0.05~0.05區(qū)間之外的基因有38個,這38個基因與預(yù)期ENC值較遠(yuǎn),與標(biāo)準(zhǔn)曲線的距離較遠(yuǎn)。本研究中的降香黃檀葉綠體基因組中的大多數(shù)基因與標(biāo)準(zhǔn)曲線距離較遠(yuǎn),說明降香黃檀葉綠體基因組密碼子偏好性更多地受選擇的影響,而受突變的影響較弱。
2.4 PR2-plot繪圖分析
通過PR2-plot繪圖進(jìn)一步分析降香黃檀葉綠體基因組密碼子偏好性的影響(圖3),PR2平面圖中4個區(qū)域的散點(diǎn)分布不均勻,大部分基因位于平面圖的下半部,尤其是右下方的基因數(shù)量最多,說明在堿基的使用頻率方面,U>A、G>C。如果密碼子偏好性完全受到突變的影響,則4個堿基的使用頻率相當(dāng),因此,降香黃檀葉綠體基因組密碼子偏好性同時受到突變和選擇的影響。
2.5 最優(yōu)密碼子確定
將降香黃檀葉綠體基因組中的52條蛋白編碼基因作為一個整體在Codon W 1.4.2軟件上運(yùn)行,通過構(gòu)建高表達(dá)基因和低表達(dá)基因庫,分別計算這兩個基因庫中的RSCU值,結(jié)果顯示(表5),ΔRSCU≥0.08的密碼子,分別為UUU(TTT)、UUA(TTA)等23個密碼子,且均為高表達(dá)的優(yōu)越密碼子(表5中用*表示),其中:11個以A結(jié)尾,8個以U結(jié)尾,3個以C結(jié)尾,1個為G結(jié)尾。ΔRSCU≥0.3(表5中用**表示)的密碼子有11個;ΔRSCU≥0.5(表5中用***表示)的密碼子分別為UUU、UUA。以同時滿足高頻率密碼子和高表達(dá)優(yōu)越密碼子作為最優(yōu)密碼子, 分別為UUU、 UUA、 AUA、GUA、UCA、AGU、CCA、UAU、GCA、CAU、CAA、AAU、AAA、GAA、UGU、CGA、AGA、GGU和GGA等19個,其中12個以A結(jié)尾,7個以U結(jié)尾。
3?討論與結(jié)論
在生物體中,密碼子在核酸和蛋白質(zhì)的翻譯方面扮演著重要作用;植物中不同密碼子的使用頻率存在差異,這種密碼子使用偏好性是物種和基因長期進(jìn)化和對環(huán)境的適應(yīng)過程中形成的,是由于多種因子共同作用的結(jié)果,其中突變和自然選擇是該現(xiàn)象形成的重要影響因素(Romero et al., 2000; Xu et al., 2011)。葉綠體是植物進(jìn)行光合作用的細(xì)胞器,亦含有相對獨(dú)立的母系遺傳基因組信息,故葉綠體基因組在揭示物種進(jìn)化、不同物種間親緣關(guān)系、物種鑒定等方面具重要價值;同時葉綠體基因工程因其可高效表達(dá)、安全等特點(diǎn)已成為植物基因工程的研究熱點(diǎn)(Wright, 1990; Duret, 2000)。因此,植物葉綠體基因組密碼子使用偏性的研究能夠揭示物種基因組進(jìn)化關(guān)系及主要影響因素。
中性進(jìn)化理論認(rèn)為堿基突變和自然選擇對密碼子第3位堿基變化的影響是中性或近中性的(Sharp et al., 1993),即通常密碼子的第1位、第2位堿基的改變會造成編碼氨基酸的改變,而第3位堿基改變則對氨基酸編碼無影響,可以認(rèn)為密碼子使用偏性在某種程度上是對偏好密碼子使用與非偏好密碼子保留間的一種平衡,是進(jìn)化過程中的一種自我保護(hù)機(jī)制;同時因密碼子第3位堿基具有的兼并性及較小的選擇壓力、GC3含量與密碼子使用偏性的顯著相關(guān)性等因素,通常將GC3作為密碼子使用模式分析的重要依據(jù)(Gu et al., 2004; Ingvarsson, 2007)。本研究中降香黃檀葉綠體基因組密碼子的GC3含量遠(yuǎn)低于前兩位,與蒺藜苜蓿(Medicago truncatula)(楊國鋒等, 2015)、紫花苜蓿(M. sativa)(陶曉麗, 2017)的密碼子3個位置的GC含量進(jìn)行比較,即紫花苜蓿GC1(45.24%)>GC2(37.30%)>GC3(28.97%),蒺藜苜蓿GC1(45.5%)>GC2(36.8%)>GC3(26.9%),3個物種葉綠體基因組在密碼子3個位置上的GC含量趨勢一致,但具體數(shù)據(jù)存在一定的差異。中性繪圖分析顯示,密碼子第1位和第2位與第3位堿基組成存在顯著差異,通過其GC含量高度保守,其密碼子偏好性主要受選擇影響,與蒺藜苜蓿一致(楊國鋒等, 2015);同時結(jié)合ENC-plot和PR2-plot等分析方式發(fā)現(xiàn)降香黃檀葉綠體基因組密碼子的偏好性受多種因素綜合影響,主要影響因素為選擇,這個結(jié)論與其同科植物蒺藜苜蓿(楊國鋒等, 2015)一致。降香黃檀葉綠體基因組密碼子偏好以AT結(jié)尾,且其最優(yōu)密碼子為UUU、UUA,與大多數(shù)高等植物的最優(yōu)密碼子NNA、NNU的模式一致(尚明照等, 2011)。這種密碼子使用模式可能由于葉綠體基因組中含有豐富的A、T堿基,且其密碼子使用模式存在顯著差異,而在進(jìn)化和親緣關(guān)系較近的植物通常具相似的密碼子使用模式,葉綠體基因組密碼子的偏好性在進(jìn)化關(guān)系上較為保守。本研究中降香黃檀葉綠體基因組密碼子偏好性主要受突變的影響,同時與其他因素一起共同作用于其密碼子使用模式,確定了19個最優(yōu)密碼子,且均為NNA和NNU模式。降香黃檀的分布區(qū)較為零散,僅分布于海南全島,及廣東、廣西和福建的少數(shù)地區(qū) (張麗佳等, 2019), 但降香黃檀葉綠體基因組密碼子的偏好性與其他蝶形花科植物基本一致,未表現(xiàn)出特異性,說明蝶形花科植物至少在密碼子偏好性方面亦較為保守。本研究為以后降香黃檀通過外源基因密碼子改造的異源表達(dá)、葉綠體基因工程和遺傳多樣性分析提供了科學(xué)的參考依據(jù)。
參考文獻(xiàn):
CHAO YE, CHANG Y, WANG MF, et al., 2012. Codon usage bias and cluster analysis on chloroplastic genes from seven crop species[J]. Acta Agric Bor Sin, 27(4):60-64.[晁岳恩, 常陽, 王美芳, 等, 2012. 7種作物葉綠體基因的密碼子偏好性及聚類分析[J]. 華北農(nóng)學(xué)報, 27(4):60-64.]
DURET L, 2000. tRNA gene number and codon usage in the C. elegans genome are co-adapted for optimal translation of highly expressed genes[J]. Trends Genet, 16(7):287-289.
DURET L, 2002. Evolution of synonymous codon usage in metazoans[J]. Curr Opin Genet Dev, 12(6):640-649.
GU WJ, ZHOU T, MA JM, et al., 2004. The relationship between synonymous codon usage and protein structure in Escherichia coli and Homo sapiens[J]. Biosystems, 73(2):89-97.
INGVARSSON PK, 2007. Gene expression and protein length influence codon usage and rates of sequence evolution in Populus tremula[J]. Mol Biol Evol, 24(3):836-844.
LIANG YN, CHEN SL, ZHANG LJ, et al., 2019. Early growth evaluation of 10 Dalbergia odorifera families in Zhaoqing[J]. For Environ Sci, 35(2):106-110.[梁遠(yuǎn)楠, 陳水蓮, 張麗君, 等, 2019. 10個降香黃檀家系在肇慶地區(qū)的早期生長評價[J]. 林業(yè)與環(huán)境科學(xué), 35(2):106-110.]
NIU Y, XU Q, WANG YD, et al., 2018. An analysis on codon usage bias of chloroplast genome of Rosa odorata var. gigantea[J]. J NW For Univ, 33(3):123-130.[牛元, 徐瓊, 王崳德, 等, 2018. 大花香水月季葉綠體基因組密碼子使用偏性分析[J]. 西北林學(xué)院學(xué)報, 33(3):123-130.]
QIN Z, ZHENG YJ, GUI LJ, et al., 2018. Codon usage bias analysis of chloroplast genome of camphora tree(Cinnamomum camphora)[J]. Guihaia, 38(10):1346-1355.[秦政, 鄭永杰, 桂麗靜, 等, 2018. 樟樹葉綠體基因組密碼子偏好性分析[J]. 廣西植物, 38(10):1346-1355.]
ROMERO H, ZAVALA A, MUSTO H, 2000. Codon usage in Chlamydia trachomatis is the result of strand-specific mutational biases and a complex pattern of selective forces[J]. Nucl Acid Res, 28(10):2084-2090.
SHANG MZ, LIU F, HUA JP, et al., 2011. Analysis on codon usage of chloroplast genome of Gossypium hirsutum[J]. Sci Agric Sin, 44(2):245-253.[尚明照, 劉方, 華金平, 等, 2011. 陸地棉葉綠體基因組密碼子使用偏性的分析[J]. 中國農(nóng)業(yè)科學(xué), 44(2):245-253.]
SHARP PM, STENICO M, PEDEN JF, et al., 1993. Codon usage: Mutational bias, translation selection, or both?[J] Biochem Soc Trans, 21(4): 835-841.
SUEOKA N, 2001. Near homogeneity of PR2-bias fingerprints in the human genome and their implications in phylogenetic analyses[J]. J Mol Evol, 53: 469-476.
TAO XL, 2017. The study on the complete chloroplast genome of Medicago sativa and Vicia sativa[D]. Lanzhou: Lanzhou University: 22-31.[陶曉麗, 2017. 紫花苜蓿和箭筈豌豆葉綠體全基因組研究[D]. 蘭州: 蘭州大學(xué): 22-31]
WANG PL, YANG LP, WU HY, et al., 2018. Codon preference of chloroplast genome in Camellia oleifera[J]. Guihaia, 38(2):135-144.[王鵬良, 楊利平, 吳紅英, 等, 2018. 普通油茶葉綠體基因組密碼子偏好性分析[J]. 廣西植物, 38(2):135-144.]
WANG YL, XU DP, YANG ZJ, et al., 2019. Effects of pruning and ethylene on photosynthetic system characteristics in Dalbergia odorifera[J]. Mol Plant Breed, 17(7):2392-2398.[王玥琳, 徐大平, 楊曾獎, 等, 2019. 修枝和乙烯對降香黃檀光合系統(tǒng)特性影響[J]. 分子植物育種, 17(7):2392-2398.]
WU XM, WU SF, REN DM, et al., 2007. The analysis method and progress in the study of codon bias[J]. Hereditas, 29(4):420-426.
WRIGHT F, 1990. The ‘effective number of codons used in a gene[J]. Gene, 87(1):23-29.
XING CB, CAO L, ZHOU M, et al., 2013. Analysis on codon usage of chloroplast genome of Eleutherococcus senticosus[J]. Chin J Chin Mat Med, 38(5):661-665.[邢朝斌, 曹蕾, 周秘, 等, 2013. 刺五加葉綠體基因組密碼子的用法分析[J]. 中國中藥雜志, 38(5):661-665.]
XU C, CAI XN, CHEN QZ, 2011. Factors affecting synonymous codon usage bias in chloroplast genome of oncidium gower ramsey[J]. Evol Bioinform, 7(7):271-278.
YANG GF, SU KL, ZHAO YR, et al., 2015. Analysis of codon usage in the chloroplast genome of Medicago truncatula[J]. Acta Pratac Sin, 24(12):171-179.[楊國鋒, 蘇昆龍, 趙怡然, 等, 2015. 蒺藜苜蓿葉綠體密碼子偏好性分析[J]. 草業(yè)學(xué)報, 24(12):171-179.]
ZHANG LJ, PU YJ, LI DD, et al., 2019. Effects of potassium silicate at different concentrations on the plant growth and physiological characteristics of Dalbergia odorifera T. Chen seedlings[J]. J Hainan Univ (Nat Sci Ed), 37(1):6-13.[張麗佳, 蒲玉瑾, 李大東, 等, 2019. 不同濃度硅酸鉀對降香黃檀幼苗生長及其生理特征的影響[J]. 海南大學(xué)學(xué)報(自然科學(xué)版), 37(1):6-13.]
ZHANG JW, JIANG ZM, SU H, et al., 2019. The complete chloroplast genome sequence of the endangered species Syringe pinnatifolia(Oleaceae)[J]. Nor J Bot, 37(5). DOI: 10.1111/njb.02201.
(責(zé)任編輯?周翠鳴)
猜你喜歡
教學(xué)月刊·小學(xué)綜合(2016年11期)2016-12-05 14:19:42
商情(2016年40期)2016-11-28 10:26:18
啟迪與智慧·教育版(2016年9期)2016-11-26 09:06:57
職業(yè)(2016年11期)2016-11-23 20:30:37
儷人·教師版(2016年15期)2016-11-22 04:29:04
中小企業(yè)管理與科技·下旬刊(2016年10期)2016-11-18 20:02:15
南北橋(2016年10期)2016-11-10 16:43:01
農(nóng)機(jī)使用與維修(2016年10期)2016-11-10 10:01:58
中國科技博覽(2016年22期)2016-11-01 14:03:21
新聞前哨(2016年10期)2016-10-31 17:41:16
