ALDH2基因rs671單核苷酸多態性位點發生(A→G)RNA編輯*
2020-11-26 08:22:16夏萬松韋四喜周春歡
國際檢驗醫學雜志 2020年22期
夏萬松,夏 英,2,韋四喜,周春歡,杜 洪,袁 婷,金 泳,黃 海△
(1.貴州醫科大學醫學檢驗學院,貴州貴陽 550004;2.貴州中醫藥大學第一附屬醫院檢驗科,貴州貴陽 550001;3.貴州醫科大學附屬醫院臨床檢驗中心,貴州貴陽 550004;4.貴航貴陽醫院檢驗科,貴州貴陽 550009)
乙醛脫氫酶2(ALDH2)rs671單核苷酸多態性(SNP)被鑒定為活性純合ALDH2 rs671(GG)、非活性雜合ALDH2 rs671(GA)、非活性純合ALDH2 rs671(AA)[1]。ALDH2 rs671SNP存在于世界近十分之一人口中,是人類最常見的變異,約5.4億人攜帶ALDH2 rs671SNP[2]。攜帶ALDH2 rs671SNP的個體飲酒后會出現酒精代謝障礙的表現,如面部潮紅和心率增加[3-4]。重要的是ALDH2 rs671SNP除了會引起酒精反應外,還能影響某些藥物的代謝,危及攜帶者的健康。ALDH2 rs671SNP可增加胃癌、食管癌、上呼吸道鱗狀細胞癌的患病風險[5-9]。此外,ALDH2 rs671SNP與多種非癌癥疾病相關,是心肌梗死[10-11]、非酒精性脂肪肝病(NAFLD)的危險因素[12]。ALDH2 rs671SNP也可以明顯增加髖部骨折風險[13]。然而,目前的研究較為局限,僅在DNA水平研究個體的ALDH2 rs671SNP與疾病的相關性,關于攜帶ALDH2 rs671SNP的個體ALDH2基因在信使RNA(mRNA)水平的改變研究尚比較少見。因此,本研究探討了ALDH2 rs671SNP的個體ALDH2基因在mRNA水平的改變,為進一步揭示ALDH2 rs671SNP影響人類健康的機制提供基礎。
1 資料與方法
1.1一般資料 將貴州醫科大學附屬醫院體檢中心的48例體檢者作為研究對象,收集所有研究對象外周抗凝全血標本,其中男24例,女24例;年齡20~40歲。本研究經貴州醫科大學附屬醫院醫學倫理委員會批準。
1.2儀器與試劑 紅細胞裂解液購于北京索萊寶科技有限公司,DNA及總RNA提取試劑盒、TRIZOL reagent、DNA純化試劑盒購于生工生物工程(上海)股份有限公司,實驗所需引物合成及聚合酶鏈反應(PCR)產物測序也由生工生物工程(上海)股份有限公司完成,反轉錄試劑盒購于TaKaRa公司,Taq DNA聚合酶購于美國Vazyme Biotech公司。ABI Pro FlexTM梯度PCR擴增儀購于賽默飛世爾科技(中國)有限公司,瓊脂糖水平電泳儀購自北京六一儀器廠,GeneGnome凝膠成像分析儀購自伯樂生命醫學產品(上海)有限公司。
1.3方法
1.3.1位置特異性引物設計 于美國國家生物技術信息中心(NCBI)數據庫下載ALDH2基因的基本信息。設計上游引物671-F3734(位于內含子11)和下游引物671-R165(位于內含子12)可特異性擴增含ALDH2 rs671SNP位點的片段。設計上游引物F1455(位于外顯子11)和下游引物R1740(位于外顯子13)可特異性擴增含ALDH2 rs671SNP位點對應的互補脫氧核糖核酸(cDNA)的片段。所有引物序列如下,671-F3734:5′-GTCCTGGGAGTGTAACCCAT-3′;671-R165:5′-CCCAGCAGGTCCTGAACTTC-3′;F1455:5′-GGACAAGGCCAATTACCTGT-3′;R1740:5′-ATTCAGCACGCAAGGATCAT-3′。
1.3.2DNA及總RNA提取、總RNA反轉錄為cDNA 根據紅細胞裂解液產品說明書裂解人外周抗凝全血紅細胞,分別用于提取DNA和總RNA。根據DNA和總RNA提取試劑盒說明書提取人外周抗凝全血白細胞 DNA和總RNA。取5 μg總RNA按照反轉錄試劑盒說明書進行反轉錄獲得第一鏈RNA反轉錄cDNA。
1.3.3PCR擴增及測序 為了擴增位于外顯子12的ALDH2 rs671SNP位點,以上游引物671-F3734和下游引物671-R165進行PCR擴增可鑒定受試者的ALDH2 rs671SNP;以上游引物F1455和下游引物R1740進行PCR擴增可鑒定ALDH2 rs671SNP位點對應的cDNA序列。引物示意圖見圖1A。PCR體系(25 μL):2×Green Taq Mix 12.5 μL,上下游引物各1.0 μL,模板1.0 μL,ddH2O 9.5 μL。PCR反應程序:94 ℃預變性4 min;94 ℃變性30 s,58 ℃退火30 s,72 ℃延伸30 s,共40個循環后72 ℃延伸4 min。PCR產物經濃度為1.5%的瓊脂糖凝膠進行電泳分離,根據DNA純化試劑盒說明書回收目的片段后進行測序。以DNA為模板的PCR產物用上游引物671-F3734進行測序,以cDNA為模板的PCR產物用上游引物F1455進行測序。
2 結 果
2.1PCR擴增白細胞的DNA和cDNA 用上游引物671-F3734和下游引物671-R165對白細胞的DNA進行PCR擴增的部分結果見圖1B。用上游引物F1455和下游引物R1740對白細胞的cDNA進行PCR擴增的部分結果見圖1C。
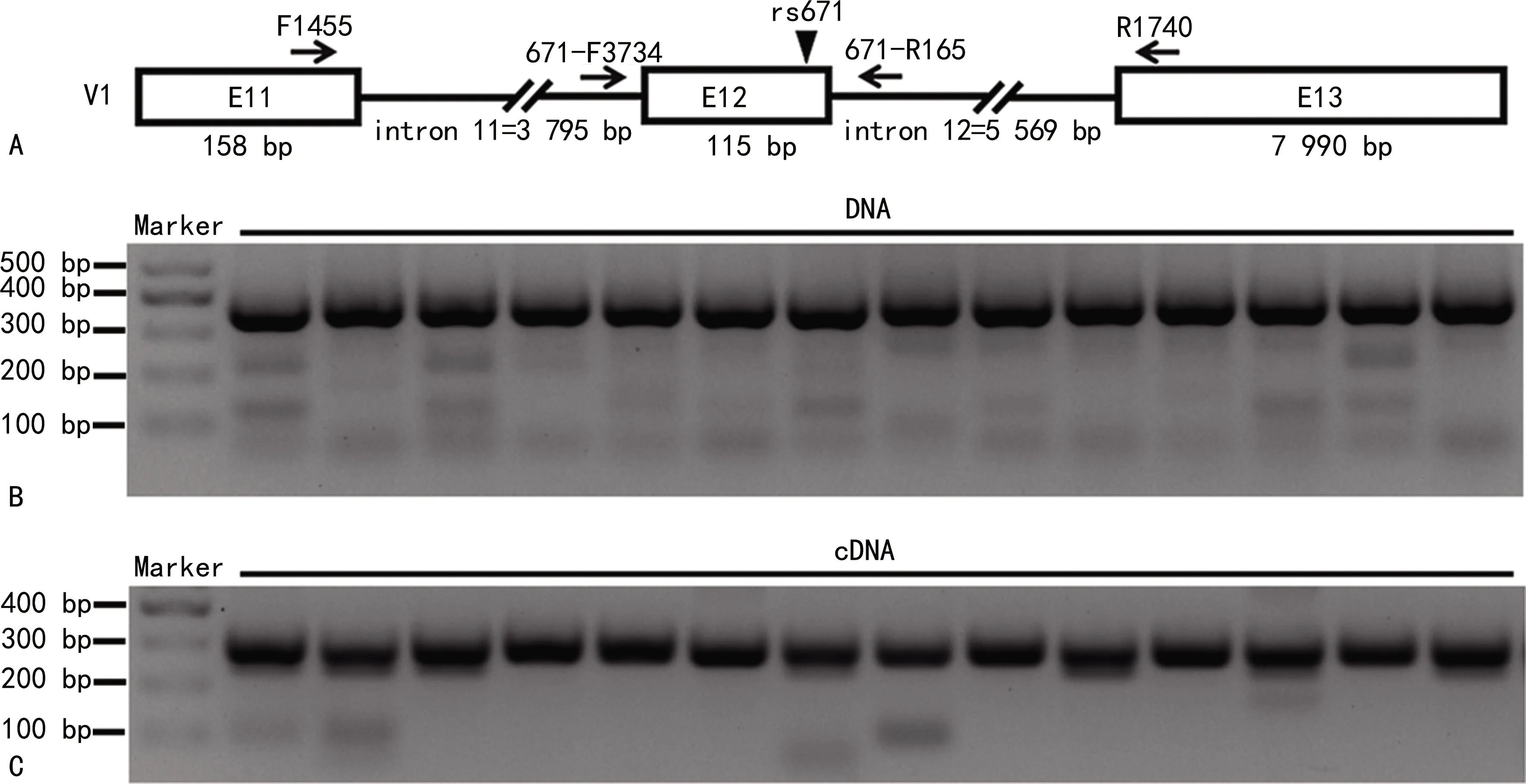
注:A為ALDH2 mRNA及引物示意圖,僅展示ALDH2變異體1(Variant,V1)的3′端的3個外顯子,?代表外顯子(E),?下方的數字表示外顯子堿基對(bp)的長度,▼指示ALDH2 rs671SNP位點,→的位置表示引物的位置;B為PCR擴增白細胞的DNA瓊脂糖凝膠電泳部分結果;C為PCR擴增白細胞的cDNA瓊脂糖凝膠電泳部分結果。圖1 來自白細胞的DNA和cDNA PCR擴增產物電泳圖
2.2PCR產物測序的結果 用上游引物671-F3734和下游引物671-R165對白細胞的DNA進行PCR擴增,PCR產物純化后用上游引物671-F3734進行Sanger測序的結果見圖2A,通過測序結果可鑒定受試者的ALDH2 rs671SNP。用上游引物F1455和下游引物R1740進行PCR擴增白細胞的cDNA,PCR產物純化后以上游引物F1455進行Sanger測序的結果見圖2B。在圖2A和圖2B中,當DNA在ALDH2 rs671SNP位點表現為雜合“GA”或純合“AA”時,cDNA在對應的位點表現為鳥苷(G),顯示出DNA與cDNA測序結果不符的情況。
2.3ALDH2 rs671(GG、GA、AA)3種基因型的基因頻率和cDNA的表現型 48例受試者ALDH2 rs671(GG、GA、AA)3種基因型的基因頻率見圖3A。在所研究的人群中,基因型為ALDH2 rs671(GG)的基因頻率最高,基因型為ALDH2 rs671(AA)的基因頻率最低(僅發現1例)。對48例受試者ALDH2 rs671SNP位點對應的cDNA的核苷酸序列進行計算,cDNA均表現為“G”,結果見圖3B。
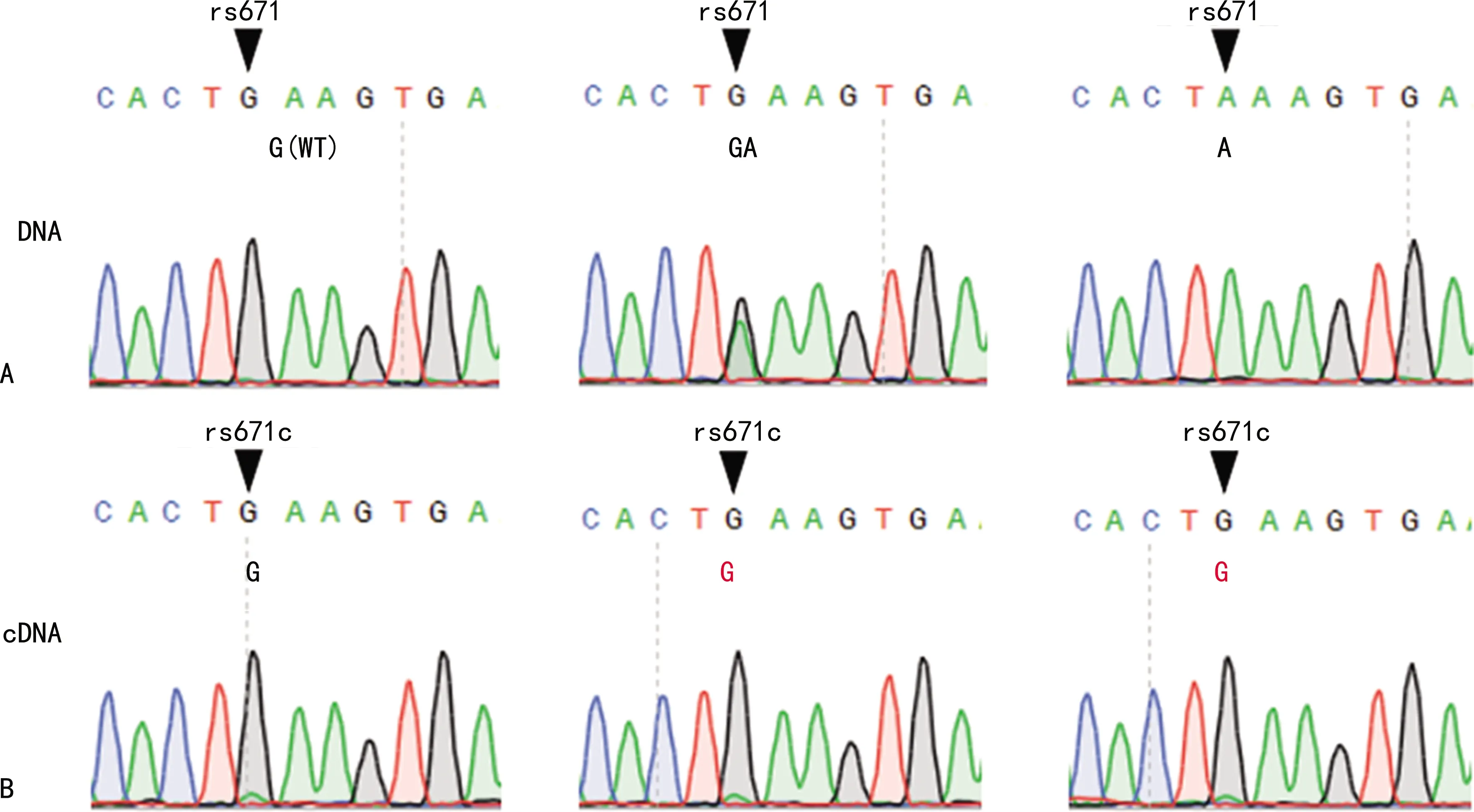
注:A為以白細胞的DNA進行PCR擴增后測序的結果,▼指示ALDH2 rs671SNP位點;B為以白細胞的cDNA進行PCR擴增后測序的結果,▼指示ALDH2 rs671SNP位點對應的cDNA的位點。圖2 配對DNA與cDNA的測序結果
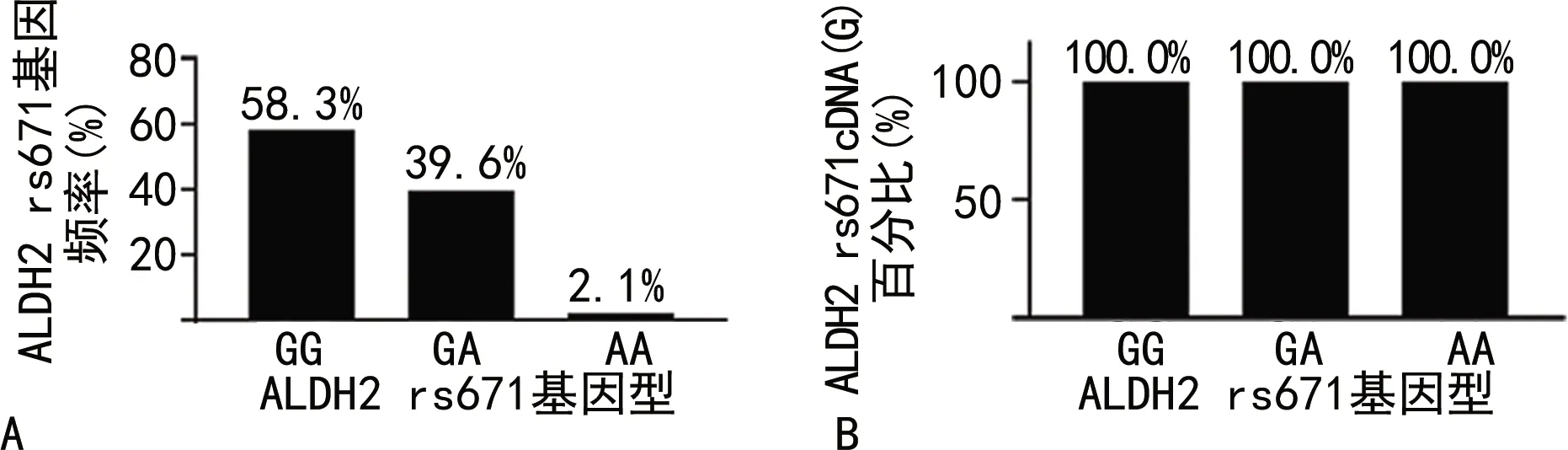
注:A為ALDH2 rs671(GG、GA、AA)3種基因型的基因頻率;B為ALDH2 rs671(GG、GA、AA)3種基因型的cDNA表現型。圖3 ALDH2 rs671(GG、GA、AA)的基因頻率和cDNA的表現型
3 討 論
ALDH2蛋白質是以四聚體的形式在體內發揮催化功能,基因型為雜合ALDH2 rs671(GA)的個體,主要是ALDH2蛋白質第504位谷氨酸→賴氨酸(504E→504K)的改變[14-15]。然而,這種改變會導致ALDH2蛋白質四聚體的穩定性下降,從而降低ALDH2蛋白質對乙醛的催化活性[16-17]。本研究的受試者中,基因型表現為雜合ALDH2 rs671(GA)的19例受試者,ALDH2 rs671SNP位點對應的cDNA都表現為“G”,并未檢測到腺苷(A)(圖2B),理論上ALDH2 mRNA翻譯出的蛋白質并不會出現ALDH2(504E→504K)的改變。矛盾的是,LAI等[18]報道基因型為雜合ALDH2 rs671(GA)的個體ALDH2蛋白質催化活性不到正常活性的1/2。因此,推測基因型表現為雜合ALDH2 rs671(GA)的受試者,ALDH2蛋白質催化活性明顯降低并不完全是因為ALDH2(504E→504K)的改變。一種原因可能是僅ALDH2的等位基因(G)被轉錄,等位基因(A)不被轉錄,ALDH2 mRNA的量相對減少,造成ALDH2蛋白質的量相對不足,導致機體對乙醛的代謝活性出現異常。盡管未檢測這些受試者ALDH2蛋白質對乙醛的催化活性。對于基因型表現為雜合ALDH2 rs671(GA)的受試者,也許機體認為等位基因(A)對于機體是“不利的”,因此,不轉錄等位基因(A),而僅轉錄等位基因(G)。
RNA編輯被定義為一種協同/轉錄后修飾機制,通過在RNA水平上插入、刪除或替換核苷酸,導致RNA序列與其模板DNA之間的差異來改變序列信息;這種修飾可以發生在編碼區,從而重新編碼氨基酸,具有典型的矯正功能;在哺乳動物中,由作用于RNA的腺苷脫氨酶(ADAR)催化的腺苷→肌苷(A→I)編輯是最常見的RNA編輯類型[19-20]。這種類型的RNA編輯在DNA水平表現為“A”,而對應的cDNA卻表現為“G”,DNA與cDNA測序時出現A→G不符的情況[21-22]。在進行蛋白質翻譯時,由于I通常被識別為G,因此A→I編輯也稱為A→G編輯[23]。本研究中的48例受試者中基因型表現為罕見的純合ALDH2 rs671(AA)的受試者,ALDH2 rs671SNP位點對應的cDNA也表現為“G”(圖2B),DNA與cDNA測序出現A→G不匹配。因此,推測基因型表現為罕見的純合ALDH2 rs671(AA)的個體ALDH2 mRNA發生了(A→G)RNA編輯。有研究表明,基因型表現為純合ALDH2 rs671(AA)時,ALDH2蛋白質的催化活性僅為正常活性的1%~5%[24]。然而,通過RNA編輯這種轉錄后的修飾機制,可以使原本DNA水平上異常的純合ALDH2 rs671(AA)在RNA水平矯正為正常的“G”,這是RNA編輯典型的矯正功能,在進化上也是一種非常好的現象。
4 結 論
48例受試者中,19例基因型表現為雜合ALDH2 rs671(GA)的個體,ALDH2 rs671SNP位點對應的cDNA都表現為“G”,基因型表現為罕見的純合ALDH2 rs671(AA)的個體,ALDH2 mRNA發生(A→G)RNA編輯,這些結果為ALDH2 rs671SNP如何影響人類健康的機制提供了新的見解。
