香菇單核菌株菌絲生長特性分析*
2019-08-13 09:05:46程爽爽杜雙田
中國食用菌 2019年7期
程爽爽,張 姣,杜雙田
(西北農林科技大學生命科學學院,陜西 楊凌 712100)
自古以來,香菇[Lentinus edodes(Berk.)Sing.]就有著“山珍之王”、“植物性食品的頂峰”等美譽,不僅營養豐富,還具有降低膽固醇,預防心血管疾病、糖尿病、佝僂病。
隨著人民生活水平的提高及保健意識的增強,香菇的市場需求量逐年增大。截止目前,我國已有“慶元香菇”、“隨州香菇”、“西峽香菇”、“平泉香菇”4個地理標志香菇產品,基本形成了以“慶元香菇”產地為核心區域的浙江、以“隨州香菇”產地為核心區域的湖北、以“西峽香菇”產地為核心區域的河南、以“平泉香菇”產地為核心區域的河北的4大產區。
香菇菌種在香菇產量及品質構成中占有十分重要的位置,尤其在以自然生態環境和傳統農藝方式為基礎發展起來的我國香菇產業中,優良菌種在產業發展所占的技術份額起到舉足輕重的作用。我國之所以能在幾年時間迅速發展為產量居世界第一的食用菌生產大國,除了因地制宜地采用適合中國國情的栽培體系和栽培方式以外,重視引進及選育適合我國栽培特點的優良菌種無疑也是一個重要因素。
我國香菇產量占食用菌總產量的70%。但在香菇的育種方面,與先進國家相比還有一定的差距。我國香菇的育種經歷了起步階段、引種階段、單孢雜交育種階段及原生質體技術選育菌種4個階段[1],主要方法有人工選擇育種、雜交育種、誘變育種、原生質體融合育種、基因工程育種、分子標記輔助育種等。其中,雜交選育是目前選育香菇新品種最有效的方法[2]。香菇的雜交育種是利用香菇的擔孢子有不同“性”的特性,選擇不同優良親本的單核菌株進行配對雜交以實現基因重組,綜合其兩親本的優良性狀,克服親本原有的不良性狀,從而育成符合人們需要的、具有雜種優勢的新品種。在雜交育種中,選擇優良單核菌株是獲得優良雜交株的關鍵,但關于單核菌株生長特性的研究甚少。
本研究通過分析香菇不同品種單核菌株的菌絲生長速率、菌落類型頻率分布、菌落長勢評分等指標以及低溫保藏對單核菌株生長的影響,比較不同香菇品種單核菌株生長性狀的差異性,以期為香菇雜交選育的材料選擇提供參考。
1 材料與方法
1.1 材料
1.1.1 品種
供試品種見表1。

表1 供試品種編號及其來源Tab.1 Test varieties and their sources
1.1.2 綜合PDA培養基
馬鈴薯200 g、葡萄糖10 g、蔗糖10 g、蛋白胨2 g、酵母浸粉 2 g、KH2PO41.0 g、MgSO4·7H2O 0.5 g、瓊脂粉12 g,水1 000 mL。
1.2 方法
1.2.1 單核菌株的獲取
參照參考文獻 [3],將單孢分離獲得的單菌落菌株接至綜合PDA平板,培養10 d,經插片鏡檢,菌絲上無鎖狀聯合即為單核菌株,即為研究材料。
1.2.2 單核菌株菌絲生長速率測定
將上述單核菌株轉接至綜合PDA平板,在(25±1)℃培養12 d,沿菌落邊緣用直徑5 mm的無菌打孔器取其菌餅再轉接在綜合PDA平板中央,記錄菌絲萌發時間,采用十字劃線法[4]測定菌絲生長速率(S,mm·d-1)公式為:

式中:D為菌落平均直徑(mm),T為培養天數(d)。
1.2.3 菌落長勢評分
根據單核菌株菌落的潔白程度、氣生菌絲的疏密及旺盛程度等將試驗菌落分為5組(優、良、中、較差、差),從差到優依次給1分~5分,不萌發菌株的長勢評分為0。
1.2.4 菌落類型
在綜合PDA平板培養中,單核菌絲較纖細,分枝少,菌絲生長速率緩慢。菌落形態可歸納為4種類型[5]:疏松型(A)、致密型(B,與雙核菌絲菌落相似)、貼生不均勻型(C)、貼生均勻型(D),如圖1所示。分別統計不同品種的單核菌株不同菌落類型的分布情況。

圖1 單核菌株菌落的類型Fig.1 Colony types of monokaryons strains
1.2.5 低溫保藏對單核菌株菌絲生長情況的影響
選取4℃低溫保藏3個月的單核菌株,在綜合PDA培養基上活化2次,依據方法1.2.2轉接綜合PDA平板,統計菌絲萌發時間,測定菌絲生長速率。
1.3 數據處理
采用Excel 2003和SPSS 20進行分析。
2 結果與分析
2.1 單孢分離及單核菌株鑒定
分離鑒定結果見表2。

表2 不同品種單孢分離及單核菌株數量統計Tab.2 Number of single colonies and number of monokaryons in different strains
由表2可見,研究中挑取的單菌落較多,但得到的單核菌株有限,表明孢子在無菌水中的分散性較差,部分孢子沒有完全分開,在今后的研究中可在制備無菌水時添加一定的表面活性劑,使孢子充分分散,以取得更好的分離效果。
2.2 單核菌株菌絲生長速率比較
不同品種單核菌株的菌絲生長速率測定結果見表3和圖2。
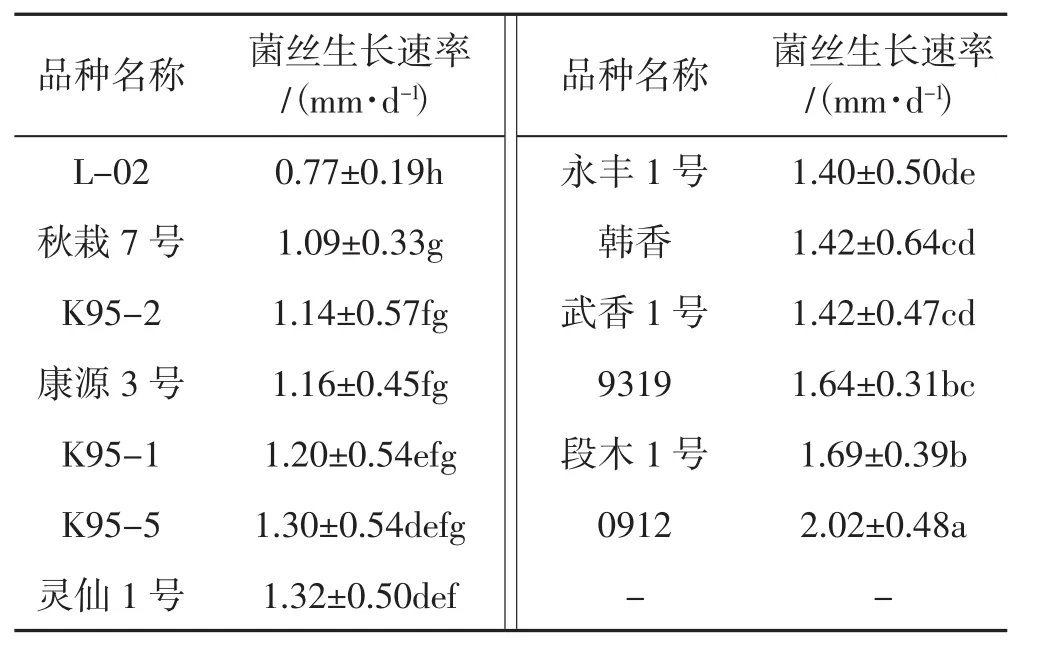
表3 不同品種單核菌株的菌株生長速率Tab.3 Mycelium growth rates of monokaryonsin in different varieties

圖2 不同品種單核菌株菌絲生長速率頻率分布Fig.2 Distribution on mycelium growth rates of monokaryons in different varieties
由表3可見,不同品種的香菇,其單核菌株的菌絲生長速率差異顯著。其中,L-02的單核菌株菌絲生長速率最小,為0.77 mm·d-1;0912的單核菌株生長速率最大,為2.02 mm·d-1,是L-02的單核菌株的2.62倍;二者的單核菌株菌絲生長速率均與其它品種的差異顯著。標準差的大小反映了同一品種不同單核菌株菌絲生長速率差異情況,標準差越大,表明不同單核菌株生長速率差異越大。可以看出,韓香的單核菌株生長速率差異最大,L-02的單核菌株生長速率差異最小。
由圖2可知,供試品種單核菌株生長速率分布接近正態分布,但不同品種的分布圖形態差異較大,表明不同品種的單核菌株生長速率差異程度不同。圖2Ⅰ中,L-02所有單核菌株生長速率均不超過1.50 mm·d-1,主要集中于 0.50 mm·d-1~1.00 mm·d-1,該品種不同單核菌株的生長速率差異性小,且生長速率小,與表3結果相同。韓香、K95-1、K95-2、K95-5、永豐1號的單核菌株生長速率分布較分散,表明該品種單核菌株生長速率差異性較大。圖2Ⅳ所示的3個品種,即段木1號、9319和0912,生長速率在0~1.50 mm·d-1之間的單核菌株很少,3個品種的單核菌株生長快的居多,平均生長速率高于其他品種。
2.3 單核菌株菌落長勢評分比較
單核菌株菌落長勢評分比較見圖3。由圖3可見,供試品種的單核菌株菌落長勢評分頻率分布接近正態分布,且集中分布在3分區域,分布頻率均在40%以上。表明香菇的單核菌株菌落長勢接近均值的居多,長勢差的單核菌株數量相對較少。供試品種中大部分品種單核菌株菌落長勢相對較好。僅在L-02、康源3號、靈仙1號、段木1號4個品種中,有少部分單核菌株的菌落長勢差,但其數量均不超過總樣本的6%。靈仙1號和永豐1號單核菌株菌落長勢較差的較多,分別為總樣本數的16.98%和16.13%。0912單核菌株菌落長勢中等的最多,分布頻率為77.19%。段木1號和武香1號菌落長勢良好的單核菌株較多,分布頻率分別為31.53%、31.25%。永豐1號菌落長勢優秀的單核菌株最多,分布頻率為16.13%。

圖3 不同品種單核菌株菌落長勢評分頻率分布Fig.3 Distribution on colony growth scores of monokaryons in different varieties

圖4 不同品種單核菌株菌落類型分布Fig.4 Distribution on colony types of monokaryons in different varieties
2.4 不同單核菌株菌落類型分布分析
不同單核菌株菌落類型分布分析見圖4。
由圖4可知,不同香菇品種單核菌落類型分布差異較大。在4種類型中,疏松型單核菌株分布頻率差異最大,其中L-02中疏松型單核菌株占86.76%,段木1號中疏松型僅占9.48%,相差9.15倍;致密型單核菌株分布頻率差異次之,其中段木1號中致密型單核菌株占49.14%,L-02中致密型占5.88%;貼生均勻型單核菌株菌落類型差異更小,其中康元3號中貼生均勻型的單核菌株分布頻率最大為36.17%,L-02的最小為2.94%;貼生不均勻型單核菌株分布頻率差異最小,靈仙1號貼生均勻型的單核菌株分布頻率最大為32.08%,L-02的最小為4.41%;同一品種的單核菌株菌落4種類型的分布頻率也存在一定差異性,韓香差異最小(20.59%~32.35%),L-02差異性最大(2.94%~86.76%)。
2.5 低溫保藏對單核菌株菌絲生長的影響
2.5.1 低溫保藏對單核菌株萌發期的影響
低溫保藏對單核菌株萌發期的影響見圖5。
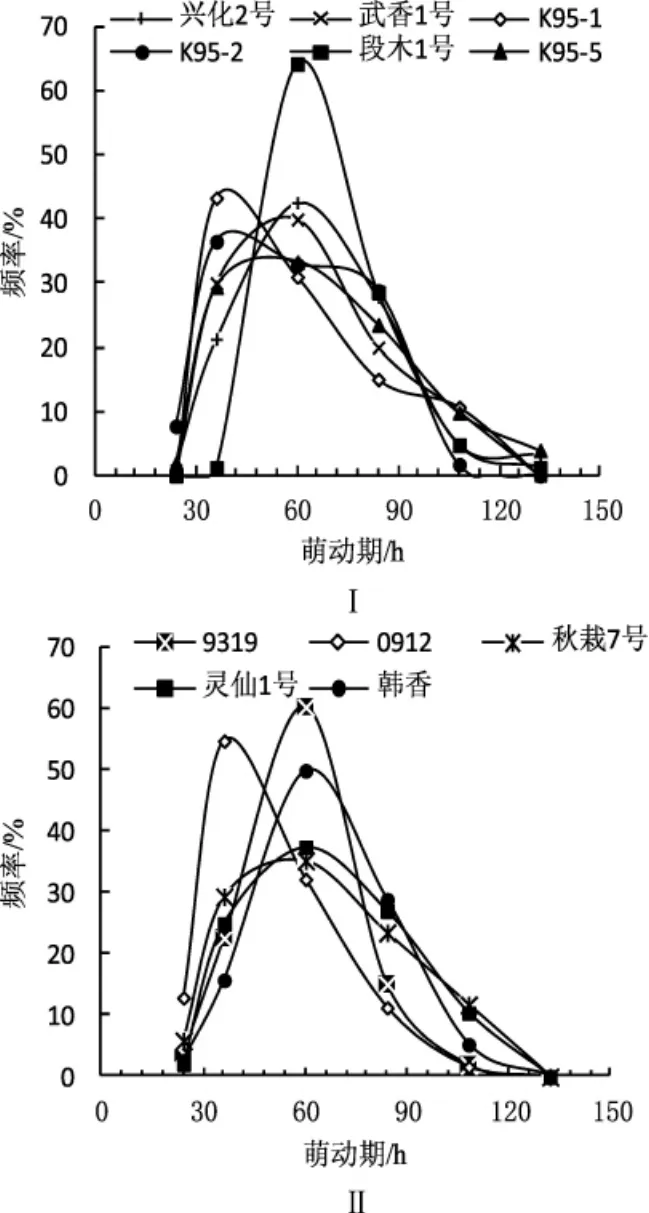
圖5 低溫保藏3個月單核菌株萌動期分布頻率Fig.5 Distribution on germination time of monokaryons
供試單核菌株未低溫保藏前其萌動期差異均在24 h內萌發。由圖5可知,保藏后的單核菌株萌動期從24 h到132 h不等,低溫保藏3個月以后,大部分菌株的萌動期延長。不同品種單核菌株萌動期分布頻率差異很大,但都接近正態分布。圖5Ⅰ中,段木1號、K95-1、武香1號單核菌株低溫保藏后24 h內均不萌發(從圖5Ⅰ中看出,3個品種的單核菌株在24 h~36 h范圍內均有萌發),其他品種24 h內有少量單核菌株萌發,其中0912最多,為總數的12.90%。0912、K95-1、K95-2的單核菌株在36 h左右萌發較多,段木1號、武香1號、L-02、9319、韓香、靈仙1號、秋栽7號的單核菌株在60 h左右萌發的較多。圖5Ⅰ中,段木1號、興化2號、K95-5少數單核菌株的萌動期為132 h,是低溫保藏前萌動期的5.5倍。總體而言,低溫保藏3個月使得大部分單核菌株萌動期延長,表明低溫保藏可導致單核菌株的活力下降。
2.5.2 低溫保藏對單核菌株菌絲生長速率的影響
不同品種單核菌株低溫保藏后的菌絲生長速率情況見表4。
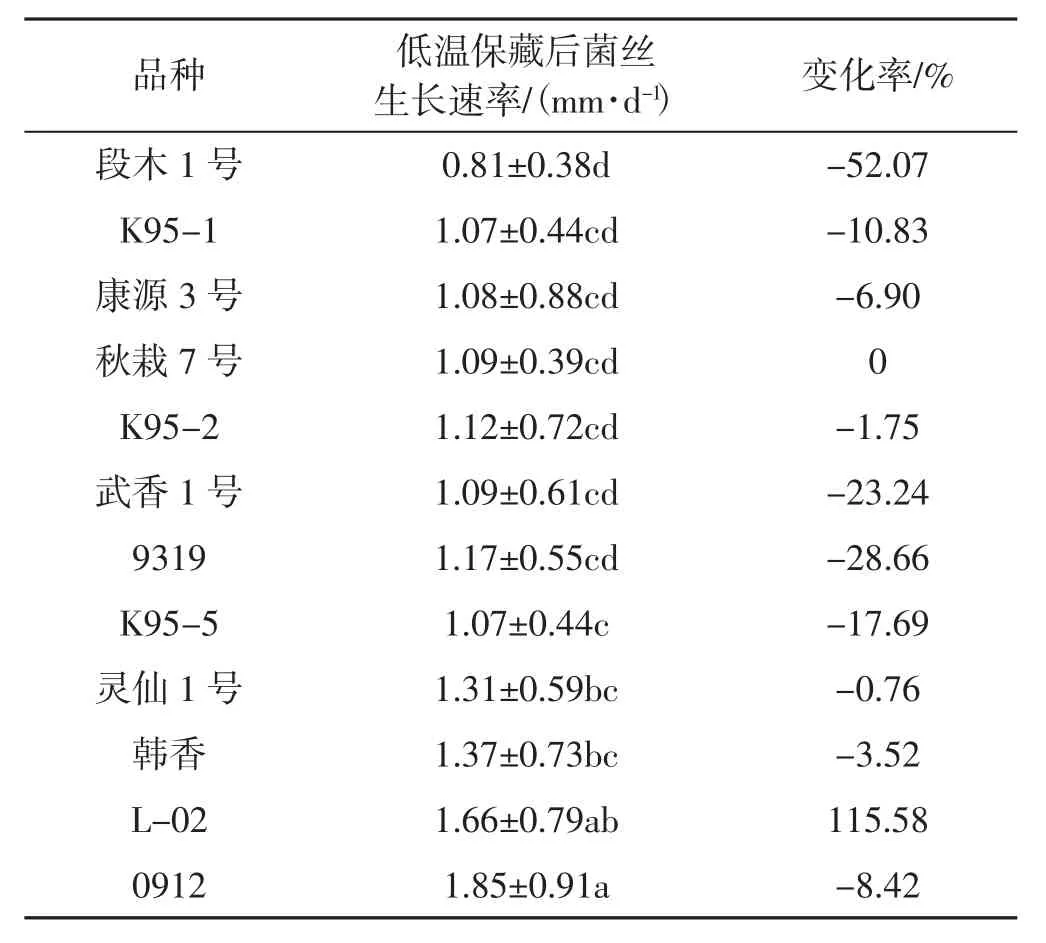
表4 不同品種單核菌株低溫保藏后的菌絲生長速率Tab.4 Mycelium growth rates of monokaryons in different varieties after cryopreservation
由表4可見,3個月的低溫保藏期對單核菌株的生長速率影響很大。L-02的單核菌株的菌絲生長速率顯著提高,是未保藏前的1.15倍。秋栽7號單核菌株的生長速率沒有受到影響。其余品種單核菌株的生長速率均有降低。其中,段木1號的下降率為52%,其次為武香1號及9319,再次為K95-5和K95-1等。由此可見,大部分香菇品種的單核菌株不宜長期低溫保藏,在單孢分離之后立即進行雜交,以免影響雜交效果。
3 討論
3.1 不同香菇品種單核菌株生長速率間差異顯著
從結果分析看出,供試13個品種單核菌株菌絲生長速率差異顯著。0912的單核菌株生長速率最高到達(2.02±0.48) mm·d-1,L-02的單核菌株菌絲生長速率最低為(0.77±0.19) mm·d-1。因為品種不同,其遺傳物質必然不同,菌絲生長速率有一定差異。因此在選擇單核菌株時不能一味地選擇菌絲生長快的菌株,林范學等[6]育種時發現,菌絲生長太快的菌株,基質營養消耗過快,香菇的產量不高,產量主要集中在第一潮、第二潮菇。Gong等[7]對157個香菇單孢單核體的生長速度等性狀進行研究,發現菌絲生長速度、生物量、萌發時間呈現連續變異,屬多基因控制,生長速度與生物量呈正相關,萌發時間與生長速率、生物量呈負相關。
3.2 同一品種單核菌株菌絲生長速率分布頻率不同
同一香菇得到的單核菌株菌絲生長速率間有較大差異,其生長速率的分布頻率呈正態分布,表明同一子實體所產生的孢子其遺傳因子有一定差異,擔孢子間有較大的選擇性。從頻率分布的標準差來看,其選擇差不同。潘迎捷等[8]研究發現,同一來源的香菇單核菌株之間存在酯酶同工酶等酶的多型性,表明同一來源單核菌株遺傳物質差異較大。因此采用單孢分離雜交選育是提高菌株生產性能的重要途徑。
3.3 單核菌株菌落長勢評分符合正態分布
菌落長勢是在菌種選擇中使用的一個綜合性概念,是菌落大小、白度、菌絲旺盛程度等指標的一個綜合指標。菌落長勢評分符合正態分布表明大部分菌株的生長情況集中在平均數附近,表現差的和優秀的僅占少數,表明以菌落長勢為指標選擇菌株時,必須有足夠的樣本數量。
3.4 不同品種單核菌株菌落類型分布頻率不同
同一朵香菇的單孢子有4種類型,在分離培養中,單孢子萌發的單核菌株在綜合PDA培養基上生長特征不同,分成疏松型、致密型、貼生不均勻型、貼生均勻型4種類型目的是選擇不同類型的單核菌株雜交。結果發現,不同的香菇品種中,單核菌株的4種類型所占比例并不一致。在供試品種中,品種9319、L-02和秋栽7號的疏松型單核菌株所占比例較高,分別達到69.49%、86.76%和72.22%,其他類型所占比例相應的就較少。韓香、康源3號、靈仙1號3個品種中4種類型的比例接近。因此,從單核菌株的菌落類型選擇雜交組合也有一定的局限性。按照經典遺傳學原理,香菇為四極性異核結合,孢子的4種類型不能完全從菌落的生長特性上反映出來。程水明和林范學[9]的研究結果表明,香菇中擔孢子交配型因子分離偏離理論預期,偏離程度栽培菌株大于野生菌株。張小雷等[10]認為,單核菌絲挑取的時間導致生長極慢的菌株丟失,進一步造成偏分離現象。所以試驗中挑取單菌落的時間可能會導致孢子萌發極慢的菌株丟失,使得菌落類型分布出現偏差,需要調整挑取單菌落的時間進行進一步的試驗驗證。
3.5 單核菌株不宜低溫長時間保藏
研究結果表明,3個月的低溫保藏,絕大部分菌株的萌發期延長,菌絲生長速率降低,表明菌株的活力下降。但也有特殊情況,品種L-02的萌發期雖然延長,但生長速率反而增加,這種情況雖不多見,但確實存在,有待于進一步研究。黃年來[5]指出,“分離到單核菌絲后,應立即進行雜交”,“香菇單核菌株隨保藏時間的延長也影響雜交成功率”。劉新銳等[11]發現,低溫保藏會降低香菇漆酶、羧甲基纖維素酶木糖醇酶和淀粉酶等酶的活力。衣川堅二郎等報道,從香菇得到的單核菌株雜交后都能正常的雙核化,將這些單核菌株保存每年轉接1次到2次,經幾年后,用其中的一些雜交發現22個親和組合中有3個為正常的雙核化,有7個為有限的雙核化,有12個為單側雙核化,大部分單核菌株隨保藏時間的延長失去了雜交能力。
香菇為四極性異宗結合的擔子菌,擔子產生4種交配型的孢子,在減數分裂產生孢子的過程中,來自2個核的16條染色體的基因發生重組,導致其攜帶的遺傳物質差異顯著。李莉云等[12]發現來自同一菌株的香菇單核菌株不僅有菌絲形態及交配型的差異性,而且存在酯酶同工酶差異。譚琦等[13]利用RAPD技術對香菇孢子單核體的遺傳差異變化進行分析,結果表明,同種交配型的孢子單核體遺傳相似性分別為66.3%和71.1%,遠低于原生質體單核體的。由此可見,香菇在減數分裂過程中染色體交換頻率較高,利用雜交選育香菇優良菌株的潛力很大[14]。深入研究單核菌株及其雜交雙核菌株的性狀,將對香菇的雜交選育工作具有重要意義。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
