裂擬迷孔菌生物學特性及馴化栽培分析*
2019-08-13 09:05:42郭艷芳
中國食用菌 2019年7期
關鍵詞:生長
汪 陽,李 碩,張 桐,郭艷芳,李 丹,張 波,李 玉
(吉林農業大學食藥用菌教育部工程研究中心,吉林 長春 130118)
木質纖維類生物質是自然界中最大的有機碳源庫,白腐真菌可以分解此類物質,該類真菌在自然界碳循環中起著極其重要作用[1]。白腐真菌在分解木質素過程中會產生錳過氧化物酶(manganese peroxidase,MnP)、木質素過氧化物酶 (lignin peroxidase)、漆酶(laccase) 3種木質素酶氧化與分解木質素,使白腐真菌及其酶在將木材轉化為食品、飼料或纖維制品方面具有潛在應用性。其中錳過氧化物酶不但可以使存在于木質素中的酚類結構單元被氧化,也可以使非酚類結構單元被氧化,是高效的木質素降解酶[2-3]。因為MnP具有降解木材中木質素的能力而被應用于造紙業,且MnP也可對有毒酚類物質染料等降解脫色,被應用于工業廢水的處理中,所以近年對MnP的研究得到了廣泛關注[4]。
裂擬迷孔菌(Daedaleopsis confragosa),俗名粗糙擬迷孔菌,隸屬于真菌界(Fungi)擔子菌門(Basidiomycota) 蘑菇亞門 (Agaricomycotina) 蘑菇綱 (Agaricomycetes) 多孔菌目 (Polyporales) 多孔菌科 (Ployporaceae) 擬迷孔菌屬 (Daedaleopsis)[5]。該種廣泛分布在北溫帶林地中,常生長在柳樹群落(濕地或沼澤)的老樹上,也可生長在其他闊葉樹木上。該種英文名字(blushing bracket)源于其受傷后的變紅反應,還有一個可替代的英文名字(thinwalled maze polypore)[6]。
裂擬迷孔菌子實體木栓質,菌蓋扇形至半圓形,菌蓋表面淺黃色至褐色,幼時被細弱絨毛,后期變光滑,具同心環帶,邊緣較銳。孔口表面乳黃色,近圓形、長方形、迷宮狀;擔孢子(6.1~7.8) μm×(1.2~1.9) μm,圓柱形,無色,光滑,非淀粉質,不嗜藍[5,7]。
裂擬迷孔菌多應用在東方造紙術中,子實體干燥后可做成紙漿制成有紋理和顏色的紙張。其造成木材白色腐朽,具有木質素酶活性。但國內對其菌絲活性和馴化栽培研究未見報道,本試驗對該菌菌絲和馴化栽培以及錳過氧化物酶活性進行了研究分析,以期為該菌的進一步開發應用提供參考。
1 材料方法
1.1 試驗材料
菌種:野生子實體采自遼寧省白狼山國家級自然保護區,采集人高洋,標本號41558,現藏于吉林農業大學菌種保藏中心(CCMJ),菌種編號2790。通過PDA培養基進行菌種分離培養,置于4℃恒溫保存。
1.2 供試培養基
PDA:馬鈴薯200 g、葡萄糖20 g、瓊脂18 g,蒸餾水1 000 mL,pH不做處理[8];碳源培養基配方:碳源20 g、磷酸二氫鉀3 g、酵母膏2 g、蛋白胨2 g、硫酸鎂1.5 g、瓊脂18 g,蒸餾水1 000 mL,pH不作處理;pH培養基配方:馬鈴薯200 g、葡萄糖20 g、硫酸鎂1.5 g、磷酸二氫鉀3 g、瓊脂18 g,蒸餾水1 000 mL;溫度培養基配方:普通PDA培養基[9-10]。
1.3 試驗方法
對碳源培養基作4個處理:葡萄糖、蔗糖、可溶性淀粉、糊精,對照組不添加碳源;氮源培養基作4個處理:蛋白胨、酵母浸粉、磷酸氫二銨、硝酸銨,對照組不添加氮源;pH作5個處理:5、6、7、8、9(滅菌后用1 mol·L-1鹽酸和氫氧化鈉調節);溫度作6個處理:15℃、20℃、25℃、30℃、35℃、40℃,每個因素、每個處理各3次重復。選用7 mm規格打孔器打孔接種至培養皿,黑暗培養,十字交叉法測量菌絲生長速度,間隔24 h劃線[11-12]。
正交試驗培養基:碳源20 g、氮源2 g、磷酸二氫鉀2 g、硫酸鎂1 g、瓊脂粉20 g,蒸餾水定容至1 000 mL,調節相應的溫度和pH。
MnP活力測定:無菌條件下,從已活化的菌株中選取直徑7 mm接種塊5個,接種至50 mL液體發酵培養基中,搖床30℃、150 r·min-1條件下震蕩培養。自接種后24 h開始,每間隔24 h取1.0 mL菌液于1.5 mL無菌離心管中,-4℃低溫離心后取上清液,-20℃保存。11 d后,統一使用試劑盒(蘇州科銘生物技術有限公司)進行MnP活力測定,試驗重復3次。
馴化栽培:從二級種制備、栽培出菇措施方面進行分析。
2 結果與分析
2.1 鑒定
使用新型植物基因組DNA提取試劑盒(康為世紀) 提取DNA,選用通用引物ITS1和ITS4對裂擬迷孔菌進行PCR擴增,將獲得的自測序列(WY2790)在NCBI數據庫進行比對,與X-78序列具有98%的相似性,定其為裂擬迷孔菌Daedaleopsis confragosa。選取NCBI數據庫序列以多孔菌屬Polyporus作為外群,使用Mega 7軟件以最大似然法構建系統發育樹,Daedaleopsis聚類到一支,另一支為外群Polyporus,如圖1所示。
從圖1可以看出,自測序列(WY2790) 與NCBI中同種序列聚集在一起,可以輔助證明,該種正是裂擬迷孔菌Daedaleopsis confragosa。
2.2 不同碳源對菌絲生長速度影響比較
4種碳源對菌絲生長的影響見表1。
由表1得出,裂擬迷孔菌菌絲在可溶性淀粉、糊精2種碳源培養基上生長較好,在蔗糖、葡萄糖2種碳源培養基上生長較慢,雖然菌絲以可溶性淀粉和糊精為碳源時,生長速度差異并不明顯,但糊精培養基中菌落邊緣不整齊,故總體來看,可溶性淀粉是最佳碳源。其次葡萄糖培養基中裂擬迷孔菌菌落邊緣整齊,蔗糖培養基菌落邊緣不整齊且菌絲生長速度最慢。從菌絲長勢看,裂擬迷孔菌菌絲在4種碳源培養基中菌絲都較為濃密。綜合菌絲生長速度、菌絲長勢,裂擬迷孔菌菌絲對供試4種碳源的喜好順序為可溶性淀粉>糊精>葡萄糖>蔗糖。
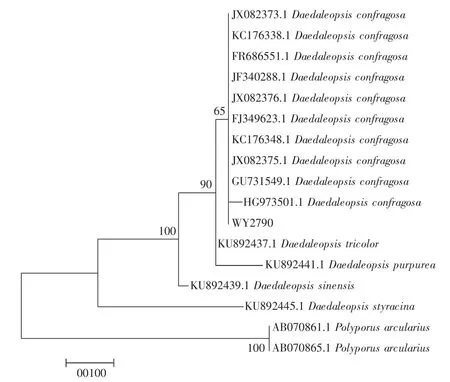
圖1 最大似然法構建的基于裂擬迷孔菌屬與多孔菌屬ITS序列的系統發育樹Fig.1 A phylogenetic tree based on the ITS sequence of the genus Daedaleopsis and the genus Polyporus constructed by the maximum likelihood method
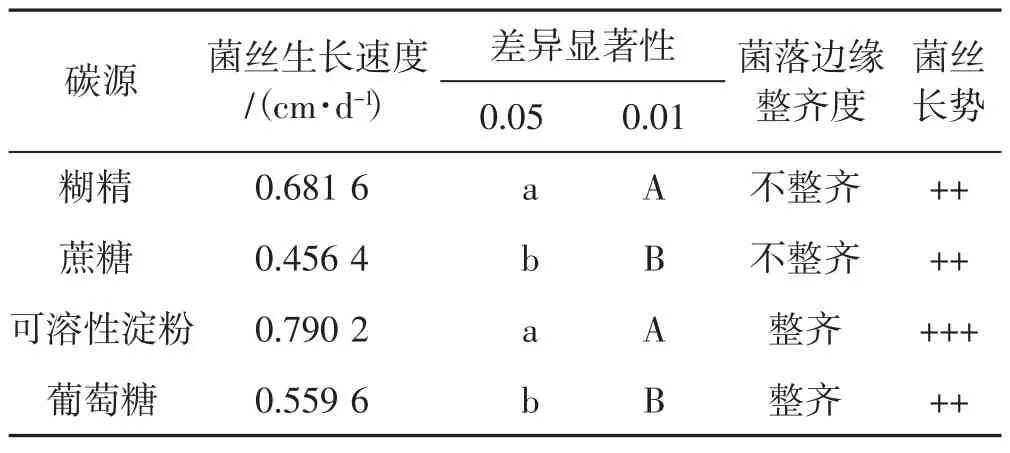
表1 4種碳源下菌絲生長情況Tab.1 Mycelial growth with four carbon sources
2.3 不同氮源對菌絲生長速度影響比較
4種氮源對菌絲生長速度影響見表2。
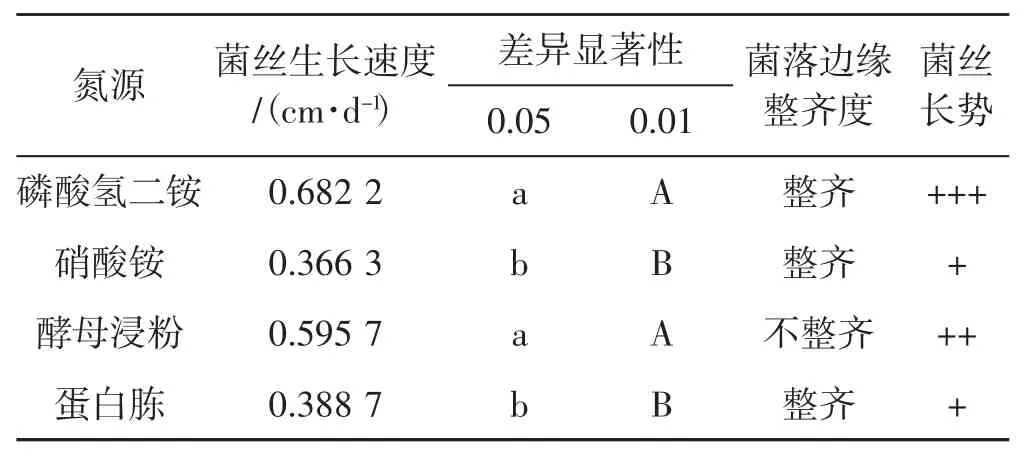
表2 4種氮源下菌絲生長情況Tab.2 Mycelial growth with four nitrogen sources
從表2可以看出,裂擬迷孔菌菌絲在磷酸氫二銨、酵母浸粉2種氮源源培養基上生長較好,在蛋白胨、硝酸銨2種氮源培養基上生長較慢。酵母浸粉培養基中菌落邊緣生長極不整齊,故磷酸氫二銨是最佳的供試氮源。而以蛋白胨、硝酸銨作為培養基時,菌絲生長速度明顯減弱。綜合菌絲生長速度和菌絲長勢可以得出,菌絲對4種供試氮源適應性差異順序為磷酸氫二銨>酵母浸粉>蛋白胨>硝酸銨。
2.4 不同pH值對菌絲生長速度影響比較
5個pH值對菌絲生長速度影響見表3。
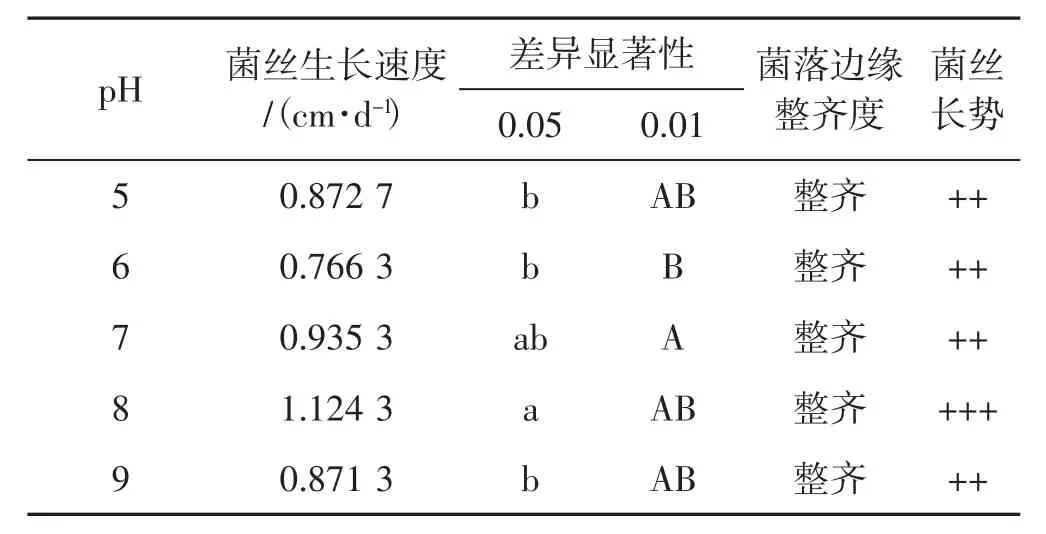
表3 5個pH值下菌絲生長情況Tab.3 Mycelial growth with five pH values
由表3得出,裂擬迷孔菌菌絲在pH為8時生長最好,其他pH時生長差異并不顯著。從菌絲生長速度得出,在5個不同pH情況下,菌絲長勢并無明顯差別,pH值為5和9時菌絲生長速度差異不明顯,pH值為6時菌絲生長速度最慢。綜合菌絲生長速度、長勢得出,裂擬迷孔菌菌絲不同pH生長優劣情況為pH8>pH7>pH5>pH9>pH6。
2.5 不同溫度梯度對菌絲生長速度影響比較
6個溫度梯度對菌絲生長速度影響見表4。
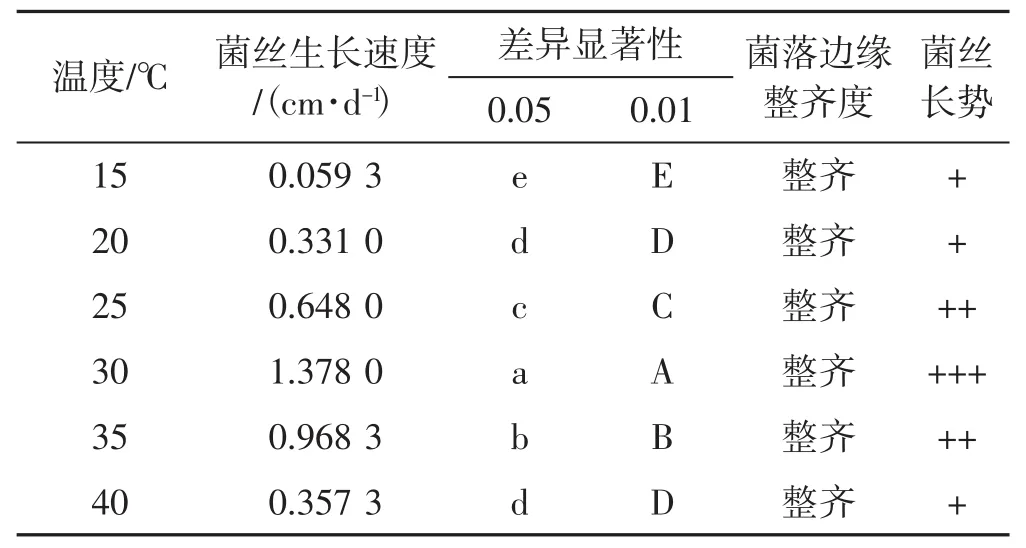
表4 6個溫度梯度下菌絲生長情況Tab.4 Mycelial growth with 6 temperature gradients
如表4所示,裂擬迷孔菌菌絲菌落直徑從起初接種1 d后至試驗結束,30℃下的培養皿菌落皆保持著生長最優勢。裂擬迷孔菌平板菌絲在15℃到30℃之間,菌絲生長情況與溫度升高成正比。裂擬迷孔菌菌絲對溫度的生長喜好為30℃>35℃>25℃>40℃>20℃>15℃。
2.6 正交試驗
單因素試驗并未考慮到各個單因素間交互作用的影響,不能將其結果作為最終結果。因此對氮源、碳源、pH和溫度4個因素進行正交試驗,從以上試驗中挑出每種因素3個最優水平,組建4因素3水平的正交分析表(表5),以期選出最適宜的搭配組合,為裂擬迷孔菌的廣泛應用奠定基礎。菌絲生長正交試驗結果直觀分析和方差分析見表6~表7。

表5 L9(34)正交試驗因素水平Tab.5 Factor level of L9(34)orthogonal test
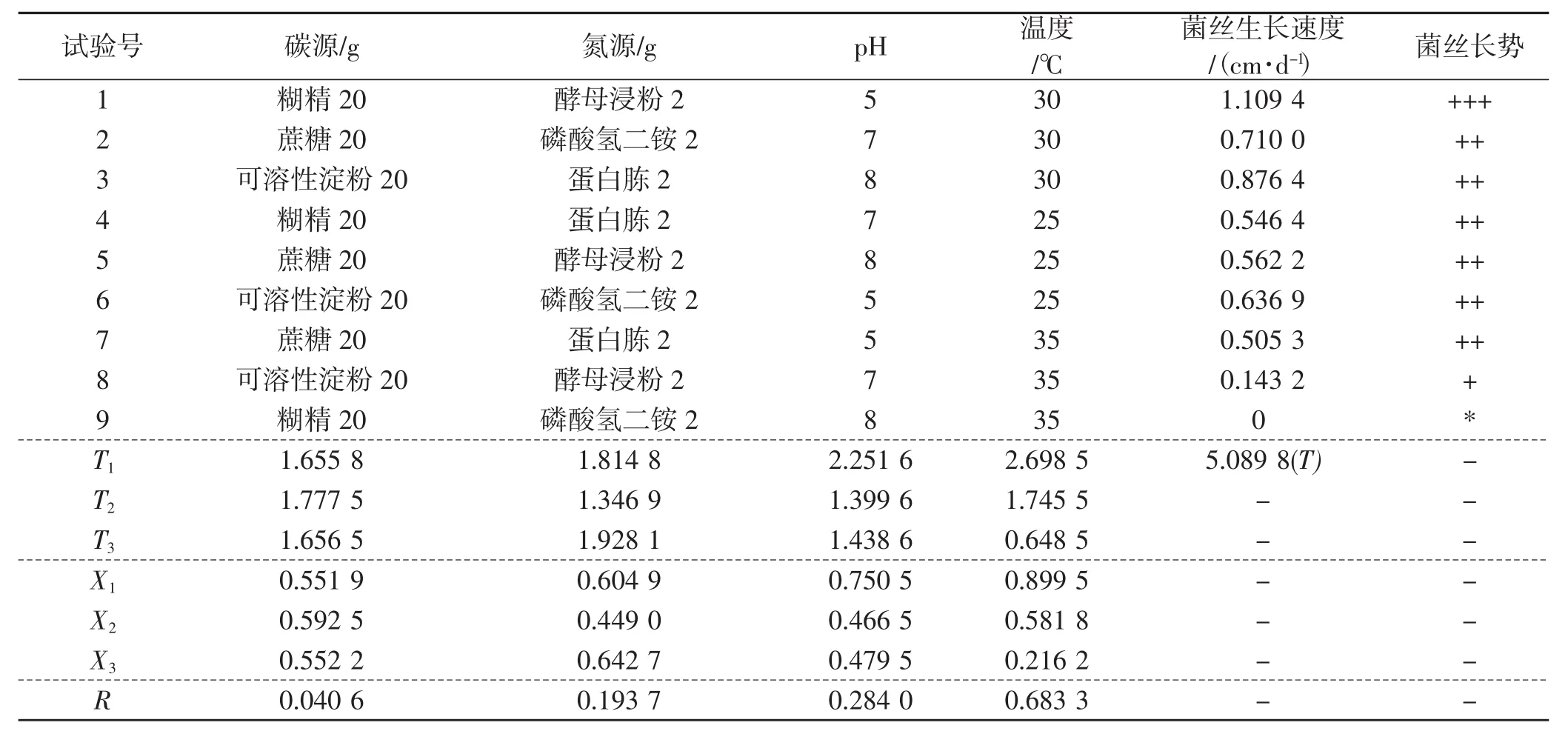
表6 菌絲生長正交試驗結果直觀分析Tab.6 Visual analysis of orthogonal test results of mycelial growth
從表6正交試驗結果直觀分析得出,溫度是影響裂擬迷孔菌最主要的因素,其極差達到0.683 3。然后是pH,極差為0.284。碳源、氮源的極差分別為0.040 6、0.193 7。從表6中可以看出,碳源的均值大小順序為X2>X3>X1,所以選擇蔗糖(X2),依次類推氮源選擇蛋白胨(X3),pH選擇pH=5(X1),溫度選擇30℃(X1)。
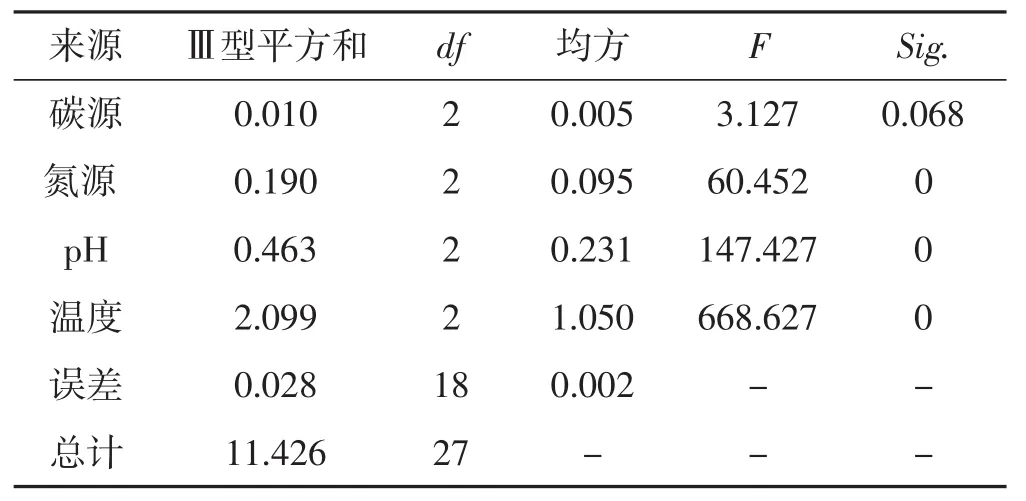
表7 菌絲生長正交試驗結果方差分析Tab.7 Analysis of variance of orthogonal test results of mycelial growth
從表7可以看出,4種生長因素對裂擬迷孔菌菌絲生長影響顯著性差異順序是溫度>pH>氮源>碳源,與直觀分析結果一致。
2.7 錳過氧化物酶活力測定
以正交試驗最優組合并加入硫酸鎂和磷酸二氫鉀作為液體發酵培養基,錳過氧化物酶活力測定結果見圖2。
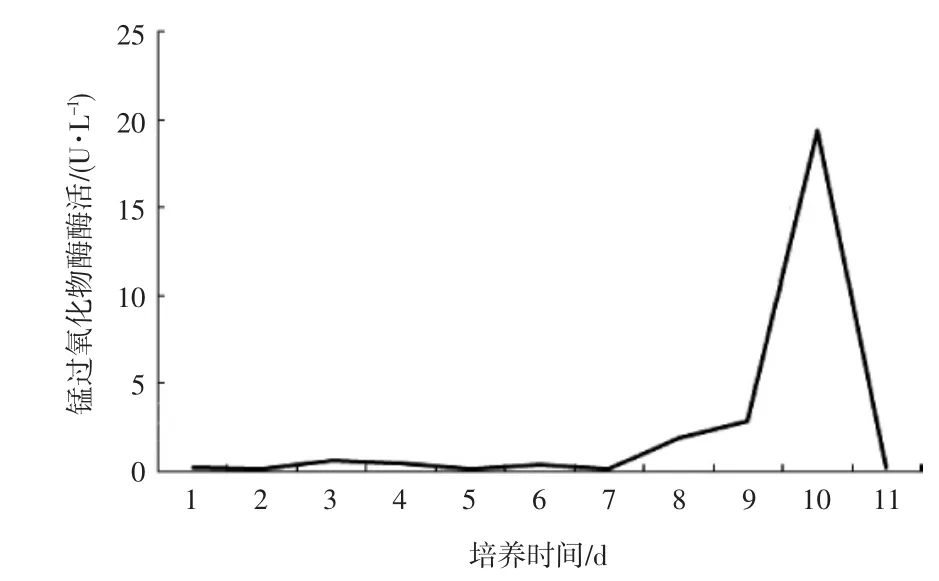
圖2 裂擬迷孔菌在液體發酵培養基中MnP活力Fig.2 Manganese peroxidase activity of Daedaleopsis confragosa in liquid fermentation medium
結果顯示MnP活力在第1天至第6天活力低且波動不劇烈,第7天活力開始上升,第10天出現峰值,達到19.4110U·L-1,第11天急劇下降為0.1652U·L-1。
2.8 馴化栽培
二級種選用麥粒菌種,500 mL罐頭瓶裝干料160 g,每瓶接整個培養皿接種塊,25℃培養[13]。
三級種配方:闊葉樹木屑78%、麩皮20%、石膏1%、蔗糖1%,加水攪拌充分混勻培養料,濕度約65%,裝入規格為17 cm×34 cm的菌袋中,每個菌袋裝濕料700 g,中間插入孔徑1.5 cm的打孔棒。121℃高壓滅菌2 h,無菌條件冷卻至25℃以下接種。
發菌和后熟:將已接入二級種的菌袋放入發菌室,保證室內溫度18℃~22℃,濕度60%~65%,黑暗培養。裂擬迷孔菌菌絲在28 d長滿菌袋。菌袋繼續放在發菌室后熟并嚴格控制發菌室溫度、濕度,9 d后完成后熟。
催蕾出菇:后熟完成后使出菇房溫度降低至14℃~16℃,并在菌袋上割口。低溫刺激有助于菇蕾的形成,在低溫刺激11 d后原基形成。在出菇期間,應保持較高的空氣濕度(95%左右),溫度控制在23℃~25℃,促進子實體形成。
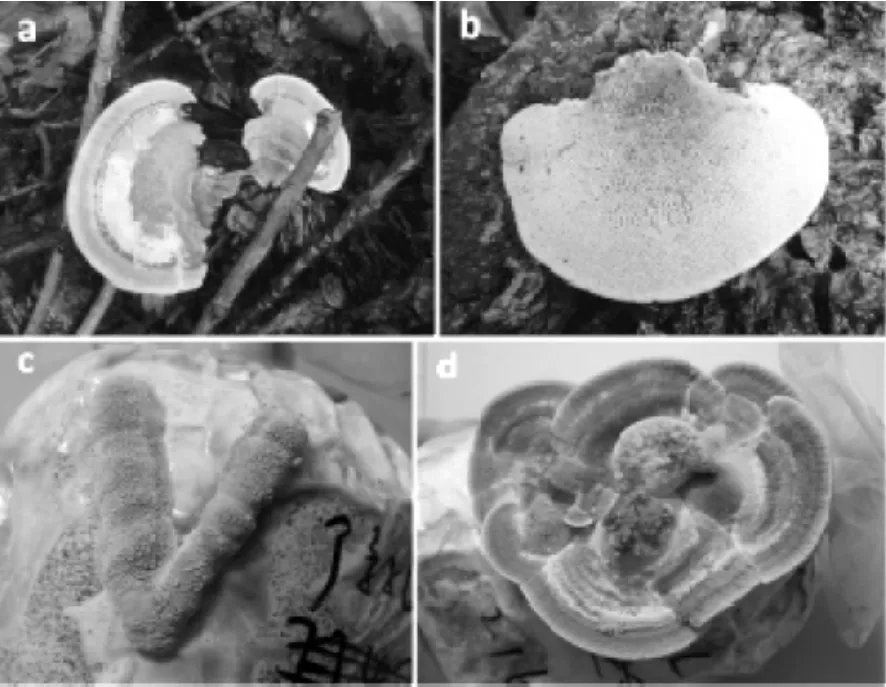
圖3 裂擬迷孔菌野生子實體及馴化栽培子實體照片Fig.3 Photos of Daedaleopsis confragosa wild fruiting body and domesticated cultivated fruiting body.
3 結論與討論
通過碳源、氮源、pH、溫度單因素試驗對裂擬迷孔菌菌絲生長的影響和正交試驗結果,最終確定裂擬迷孔菌在糊精作為碳源,酵母浸粉作為氮源,pH為5,培養溫度為30℃時長勢最好。在碳源單因素試驗中,菌絲對多糖碳源具有明顯的偏好性,碳源單因素試驗中可溶性淀粉培養基菌絲長勢最好,與Gary[3]報道的可溶性淀粉是最易被真菌分解的多糖結果相同。在正交試驗中,以糊精作為碳源時菌絲生長最優,其次可溶性淀粉,與單因素試驗結果相反。正交試驗考慮了各因素間的交互作用,前文已得出4種因素對裂擬迷孔菌菌絲生長影響顯著性差異順序是溫度>pH>氮源>碳源,碳源較其他3種因素對菌絲生長優劣的影響較小,結合魯鐵[9]、丁野[10]、程國輝[14]單因素試驗及正交試驗結果推測,多種因素共同作用時菌絲的生長會偏重對其影響更大的因素,而對影響較小的因素采取一定適應性。
在氮源單因素試驗中,菌絲長勢較好的依次是無機氮源磷酸氫二銨和有機氮源酵母浸粉,正交處理中菌絲長勢在酵母浸粉作為氮源時最優。蛋白胨和酵母浸粉都是有機氮源,而前者作為氮源時菌絲生長速度不及后者,這是因為酵母浸粉中氮源物質主要以易吸收的蛋白質降解產物形式存在,而降解產物特別是氨基酸可以通過轉氨作用直接被菌絲利用,為速效氮源,有利于被菌絲更快地利用;蛋白胨中氮源物質存在的形式多是大分子蛋白質,要被菌絲吸收利用則需要進一步降解成小分子的肽和氨基酸,是遲效氮源,利用速度緩慢。將此最優的正交處理與正交分析表試驗號2的正交處理相比,按前文所得的溫度、pH是影響菌絲生長最重要的2種因素,且試驗號2的正交處理選用氮源是單因素試驗中最優氮源,可2種正交處理菌絲生長速度差異卻接近0.4 cm·d-1,值得注意的是蔗糖為雙糖,糊精為多糖,推測在復合處理時,多糖碳源相比其他碳源更能促進菌絲生長,這與Paul Ander&Karl-erik Eriksson[15]在研究中發現多糖物質的存在,對于白腐真菌菌絲生長特性的選擇有較大影響一致。
本試驗中,菌絲在30℃情況下生長最快,在35℃時,平板生長顯著減慢,而接種塊在40℃且菌絲長度達到約2 cm時,菌絲生長即停止。與劉秀明等[16]基于白靈側耳菌絲保護酶活性與不同溫度間的聯系研究,推論溫度過高會抑制菌絲酶活性,從而抑制菌絲的生長速度一致。
試驗得出裂擬迷孔菌菌絲在pH值5~9范圍內均長勢較好,并且菌絲在pH為8的情況下生長最快,這與Sérgio Luiz Moreira Neto等研究得出大多數多孔菌適宜在pH顯酸性環境中生長不一致[17]。雖然裂擬迷孔菌可以侵染活木,是森林木材病原菌,可其只在較少情況下才會侵染活木[18-19],而不同的酸堿環境可能是真菌對植物致病性的重要決定因素[20],Nir Yakoby等[21]、Dov Prusk y等[22]對 Sclerotinia sclerotiorum的研究中發現在酸性條件下,2種可能在感染植物過程中起重要作用的酶:acp1(編碼非天冬氨酸蛋白酶)和pg1(編碼內聚半乳糖醛酸酶)的表達活性升高,推測裂擬迷孔菌菌絲偏好堿性,可能是其較少生長于活立木上的原因。
裂擬迷孔菌為白腐真菌,目前可有效對木質素進行降解的是白腐真菌[23],試驗得出的裂擬迷孔菌錳過氧化物酶活性峰值是19.411 U·L-1。作為一種誘導酶類,錳過氧化物酶的合成水平也特別易受誘導劑的影響。在何平等[4]對乳白耙齒菌的誘導劑篩選研究中,適宜的誘導劑能大幅度地提高乳白耙齒菌的MnP活性,使之從12.700 U·L-1升高到203.226 U·L-1。由于試驗已得出裂擬迷孔菌可馴化栽培,后續試驗可對裂擬迷孔菌進行誘導劑篩選,選出最優誘導劑,使其在染料等有毒酚類物質降解脫色與工業廢水治理中發揮較好作用。同時裂擬迷孔菌可以有效地清除·OH[24]。由此可見,裂擬迷孔菌具有良好的經濟應用前景。本試驗初步探索了裂擬迷孔菌的生長條件,成功栽培出白腐真菌裂擬迷孔菌,為規模化的木質素降解的進一步開發、生產提供了基礎數據,也對該屬其他種的生物學特性馴化栽培研究提供了參考。后續試驗可對裂擬迷孔菌的栽培配方、藥理活性方面進行篩選研究,也可進一步深入探討分解木質素酶系統等。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
