NaCl脅迫對紅砂萌發的影響及萌發期耐鹽性評價
2019-01-23 12:05:08劉曉威楊秀艷武海雯支曉蓉朱建峰張華新
生物技術通報 2019年1期
劉曉威 楊秀艷 武海雯 支曉蓉 朱建峰 張華新
(1. 國家林業和草原局鹽堿地研究中心,北京 100091;2. 中國林業科學研究院天津林業科學研究所,天津 300457)
目前,除了南極洲等極少數地區待查外,全世界大部分地區都有各種類型的鹽漬土分布,據不完全統計,全世界鹽漬土面積為9.543 8×108hm2,并呈現出逐年遞增的趨勢,全世界每年約有1.0×106-1.5×106hm2的土地成為鹽漬土,鹽漬土已成為限制農林業發展的主要因素之一[1-2]。鹽脅迫對植株的影響體現在植物生長發育的各個階段,在鹽脅迫條件下,植物種子細胞內水勢降低,造成種子生理性吸水困難。同時,類囊體膜結構發生變化,碳同化減少,影響光合作用[3-4]。此外,鹽脅迫下,活性氧代謝紊亂,影響蛋白質合成。與此同時,植物為了抵御鹽脅迫會合成各類有機物質造成耗能增加,最終導致植物生長緩慢[5]。我國鹽漬土約為9 913×104hm2,分布范圍廣、鹽分類型多樣、生態修復極為困難,且近半個世紀以來,由于不合理的灌溉而引起的次生鹽漬化問題已經成為我國農業發展的重要阻礙因子,如內蒙古河套灌區、寧夏銀川灌區等均有灌溉不當而引起次生鹽堿化發生[6]。
如何開發利用鹽堿土,破解鹽堿地區植被更新困難及農作物減產等難題,已迫在眉睫。科技人員研究發現,在鹽堿地區長期的植物演化過程中,演化出一種特殊的植物類型——鹽生植物,它能夠在離子濃度為200 mmol/L以上的生境中完成其生活史[6]。鹽堿地區生態修復的第一步就是如何使植物完成其生活史,而生活史的起始階段就是種子萌發期,它直接影響著后期的成活率、結實率以及物種的分布和豐度。萌發率的降低是非鹽生植物在鹽堿地實現種群延續的主要限制因素,故鹽堿地上的非鹽生植物多以人工移植為主、且類型單一,但是鹽生植物卻可以在鹽堿地上順利度過種子萌發、完成自然更新和種群延續[7-8]。因此,以鹽生植物為材料,解析鹽生樹種種子萌發期對于鹽脅迫的生理生化響應特征,闡釋耐鹽機理、確定耐鹽閾值對于理解植物種子萌發過程中的耐鹽機制、篩選種子萌發階段的耐鹽基因、提高植物種子萌發期耐鹽性,攻克鹽堿地植物天然更新障礙具有重要理論意義和現實價值。
紅砂(Reaumuria soongorica)是檉柳科鹽生植物(泌鹽植物),高15-25 cm,主要分布在我國西北部、內蒙古及東北西部鹽土荒漠、半荒漠沙地等具有不同程度的鹽漬化土壤中,紅砂一般可通過鹽腺將鹽分排出體外,以減少鹽分危害。
目前,國內外關于紅砂的研究主要集中在種子萌發的影響因素、外界因素對紅砂生長、分布的影響等方面[9-14]。朱恭等[15]發現,種子萌發的最低滲透閾值為-1.8 Mpa,土壤含水量在3%以上,黑暗條件下有利于紅砂種子的萌發,種子萌發的適宜溫度一般為15-25℃。牛宋芳等[16]發現經赤霉素處理的紅砂種子萌發率可高達78%。在有關鹽堿逆境條件下紅砂種子萌發研究方面,陳金元等[17]認為,混合鹽堿脅迫對于紅砂種子的萌發及萌發后幼苗生長均有抑制作用。高茜等[18]研究了2種不同鹽對紅砂種子吸脹過程中生理特性的影響及對種子的傷害程度發現,CaCl2的傷害大于NaCl。周航宇等[19]發現在高鹽條件下,紅砂能夠吸收大量Na+并將其絕大部分通過鹽腺排出體外或積累在地上部中,從而減輕Na+毒害。而在紅砂分子研究層面,Du等[20]研究發現,從長葉紅砂中克隆出的轉錄因子RtWRKY1可在擬南芥中進行轉化,且過表達植株中RtWRKY1主要通過調控抗氧化系統、離子轉運系統等提高擬南芥的耐鹽性。劉丹等[21]發現在干旱和UV-B雙重脅迫下,紅砂中產生了356個上調和248個下調的差異表達基因,且這些基因多在固碳作用等生物過程中顯著富集。
當前,關于鹽堿脅迫下紅砂的研究多集中在萌發后生長階段,聚焦萌發期的研究多停留在生長形態水平,而關于萌發期生理生化指標的研究及萌發期耐鹽閾值和萌發后生長閾值的計算等鮮有報道。本研究以泌鹽植物紅砂為材料,探究鹽脅迫對紅砂種子萌發及萌發后生長和生理生化指標的影響,確定紅砂萌發耐鹽閾值及最適鹽濃度、萌發后生長耐鹽閾值及最適鹽濃度,旨為解析紅砂萌發期耐鹽機制及后續研究提供數據支撐。
1 材料與方法
1.1 材料
供試紅砂種子于2017年9月收集于內蒙古巴彥淖爾市烏拉特中旗甘其毛都鎮S212省道南側(N:42°09′56.69″,E:107°43′21.95″),種子采集完成后,去雜、晾曬、置于室溫下干燥保存,紅砂種子千粒重為(1.137±0.121)g。
1.2 方法
1.2.1 實驗設計 紅砂種子萌發實驗于培養箱中進行。實驗采取NaCl脅迫處理,共設置7個濃度梯度(CK、50、100、150、200、300 和 400 mmol/L),每個處理3次重復。隨機選取顆粒飽滿、大小均勻、無破損的紅砂種子,經75%酒精消毒1 min后,用蒸餾水沖洗3-5次,吸水紙吸干水分備用,加入7 mL處理液置于墊有兩層濾紙的培養皿上,每皿放20粒種子,隨后,將添加處理液的培養皿置于培養箱中,在黑暗條件下設置溫度為25℃,相對濕度為55%進行培養。每天定時稱重補水以維持鹽溶液濃度的相對穩定,并以胚根明顯突破種皮1 mm左右為發芽標準,每天統計發芽數,試驗持續14 d。1.2.2 生長指標的測定 萌發率:正常發芽的種子數/供試種子總數×100%;相對萌發率:(處理種子發芽率/對照種子發芽率)×100%;萌發勢:發芽種子數達到高峰時正常發芽種子總數與供試種子總數的百分比;相對萌發勢:(處理種子發芽勢/對照種子發芽勢)×100%;相對鹽害率:(對照發芽率-鹽處理發芽率)/對照發芽率×100%;發芽指數=∑Gt/Dt,[Gt為不同時間(d)的發芽量,Dt為相應的發芽試驗天數];活力指數(VI)=S×GI[S:平均根長(mm),GI:萌發勢];株高和根長:在脅迫14 d后,每個重復取3棵測株高與根長,以3個重復的平均值作為株高和根長;地上鮮重:脅迫14 d后,每個重復隨機選取5株植株,除去根部、稱重記錄;地下鮮重:脅迫14 d后,每個重復隨機選取5株植株,保留根部、稱重記錄。
1.2.3 生理生化指標的測定 分別采用硫代巴比妥酸法、茚三酮染色法、創木酚染色法和氮藍四唑光還原法測定丙二醛含量(Malondialdehyde,MDA)、脯氨酸含量(Proline,Pro)、過氧化物酶含量(Peroxidase,POD)和超氧化物歧化酶含量(Superoxide dismutase,SOD)[22],每個處理測 5次,取均值。1.2.4 數據處理 采用Excel 2007進行數據整理、作圖,用SPSS17.0中的One-way ANOVA進行不同鹽濃度下待測指標間的差異性檢驗,LSD多重比較。耐鹽閾值的確定以NaCl濃度為自變量,相對萌發率、植株全長為因變量,以相對萌發率及植株全長下降到對照的50%為標準,擬合回歸方程,分別確定萌發耐鹽閾值及萌發后生長耐鹽閾值(所有數據的處理均做標準誤差分析)。
2 結果
2.1 NaCl脅迫對紅砂種子萌發的影響
運用不同濃度的NaCl對紅砂種子進行萌發實驗(圖1和表1),與對照相比,適當低濃度(50 mmol/L)NaCl處理后,紅砂種子的萌發數增加,相對萌發率升高且大于100%(P>0.05),相對鹽害率下降為負,萌發勢、相對萌發勢、萌發指數和活力指數增加。但是,隨著鹽濃度的升高,紅砂種子的萌發時間相比CK滯后2-7 d,且萌發率、發芽指數、活力指數顯著降低(P<0.05),相對鹽害率顯著增加,萌發勢下降。當NaCl濃度達到400 mmol/L的時候,紅砂種子在處理時間內已經不能萌發,相對鹽害率達到了100%。
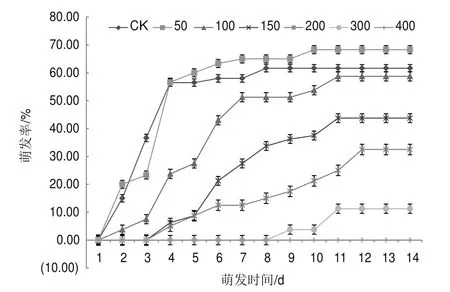
圖1 不同濃度NaCl處理下紅砂種子萌發率
為了探究脅迫下不能萌發的種子是否具有活力,分別將400 mmol/L NaCl處理15、25和35 d的種子進行復萌處理,處理時間仍為14 d,以復萌處理的第1天開始記錄。結果發現,復萌處理后,紅砂種子快速萌發,且萌發集中。復萌第1天的相對萌發率達105%,脅迫處理15、25和35 d的種子復萌情況相當,差異性不大,這符合自然條件下,紅砂種子在雨后快速、集中萌發的特征。
整體來看,紅砂種子的萌發具有以下特點:(1)條件適宜時,快速集中萌發,萌發速率先升高后降低;適當低濃度鹽促進萌發,高濃度鹽抑制萌發;(2)紅砂種子處理12 d左右,紅砂萌發達到穩定,此時間點可作為驗證紅砂種子是否能夠萌發的相對時間點;(3)脅迫條件下不能萌發的種子仍保持活性,可在條件適宜時快速集中萌發,以維持物種延續。
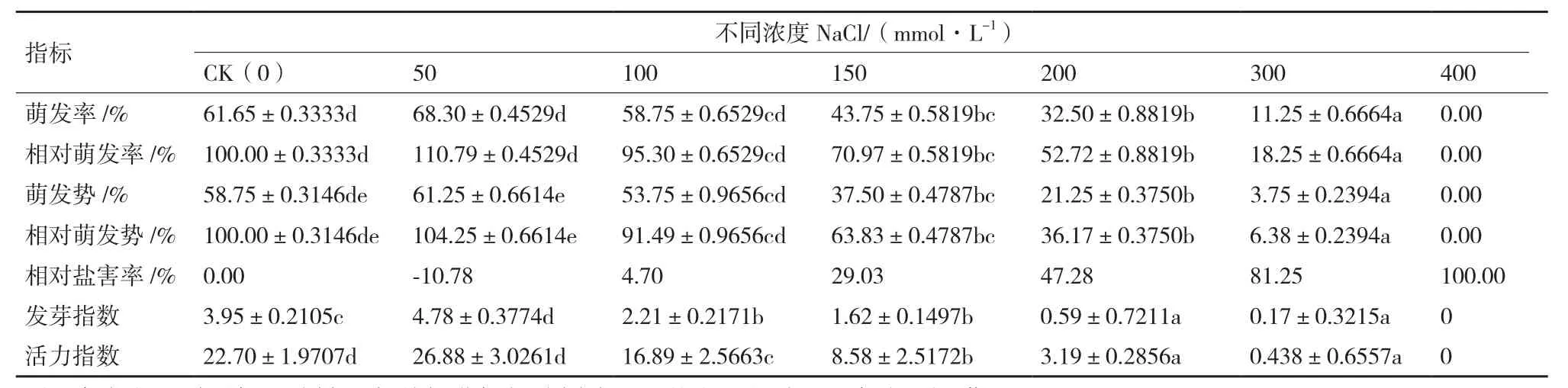
表 1 不同濃度NaCl下紅砂萌發指標
2.2 NaCl脅迫對紅砂生長的影響
為探究鹽脅迫對紅砂萌發及后期幼苗生長的影響,以萌發后期紅砂幼苗的根長和苗高作為指標(圖2-圖4),當鹽濃度為50 mmol/L時,根長增加13.55%;當NaCl濃度大于50 mmol/L時,其長度相比CK顯著降低(P<0.05);當NaCl濃度為300 mmol/L時,根長降低69.77%;當NaCl濃度為50 mmol/L時,株高增加36.37%;當NaCl濃度增加時,株高相比CK降低;當鹽濃度為300 mmol/L時,株高相比對照降低51.05%,差異性顯著(P<0.05)。當鹽濃度為50 mmol/L時,地上鮮重和地下鮮重相比CK分別3.54%和4.31%,但增加不顯著(P>0.05),隨著濃度增加,地上和地下鮮重較CK顯著下降。
整體來看,鹽濃度為50 mmol/L時,根長、株高相比CK顯著增加,但地上和地下鮮重增加不明顯;隨著鹽濃度的增加,根長、株高和鮮重下降顯著,但根長變化更為明顯,下降幅度更大。
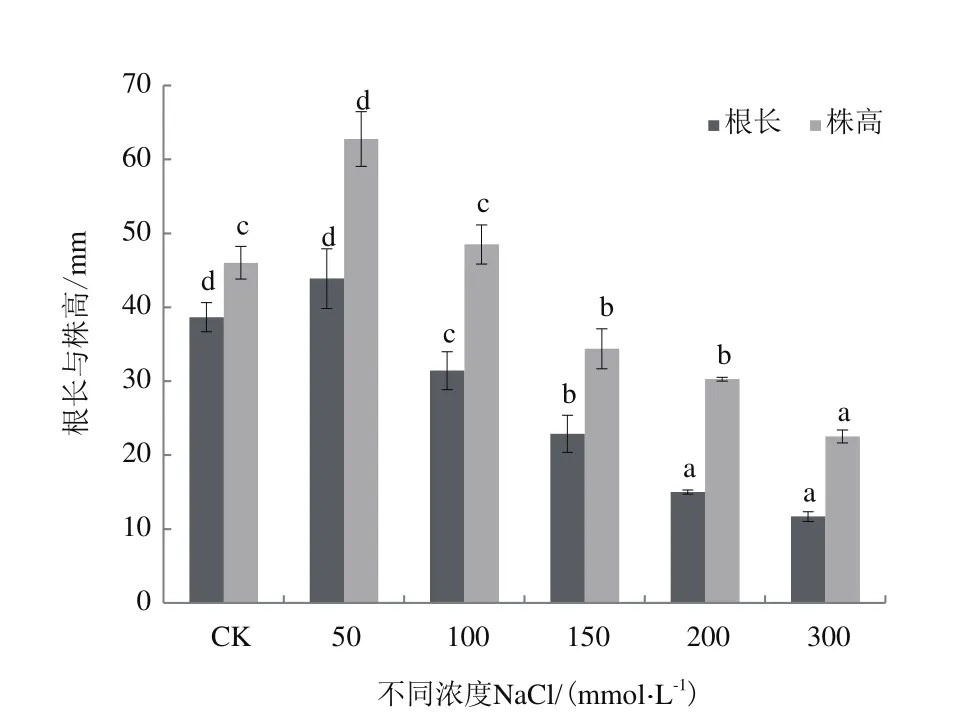
圖2 不同濃度NaCl處理下紅砂根長株高變化情況
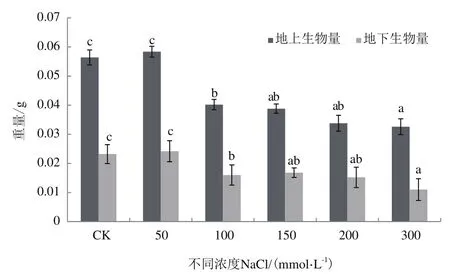
圖3 不同濃度NaCl處理下紅砂地上和地下鮮重
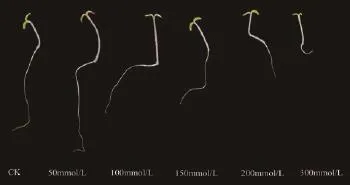
圖4 紅砂經400 mmol/L NaCl處理14 d后的生長表型圖
2.3 NaCl脅迫對紅砂萌發期生理生化指標的影響
2.3.1 NaCl脅迫對紅砂中MDA、Pro含量的影響 由圖5可知,隨著NaCl濃度增加,紅砂植株內的MDA含量較對照分別增加31.05%、164.03%、244.09%、282.12% 和 389.21%(P<0.05),MDA 的增加將會影響損傷細胞膜結構,影響膜滲透性,同時積累活性氧。
結果發現,在MDA增加的同時,作為滲透調節物質的Pro含量也顯著增加,與CK相比,不同NaCl濃度下的Pro含量分別增加25.95%、94.22%、91.59%、100.18%和105.22%,且差異性顯著(P<0.05),但 100、150、200和 300 mmol/L NaCl濃度下的Pro含量差異不大(P>0.05)。
2.3.2 NaCl脅迫對紅砂中酶活性的影響 由圖6可知,隨著NaCl濃度的增加,SOD的活性逐漸增強,與CK相比,不同濃度所對應的SOD活性分別增加237.23%、315.05%、631.35%、716.69%和699.58%,且增加顯著(P<0.05),但是 50和100 mmol/L之間、0和300 mmol/L之間的SOD活性差異不顯著。
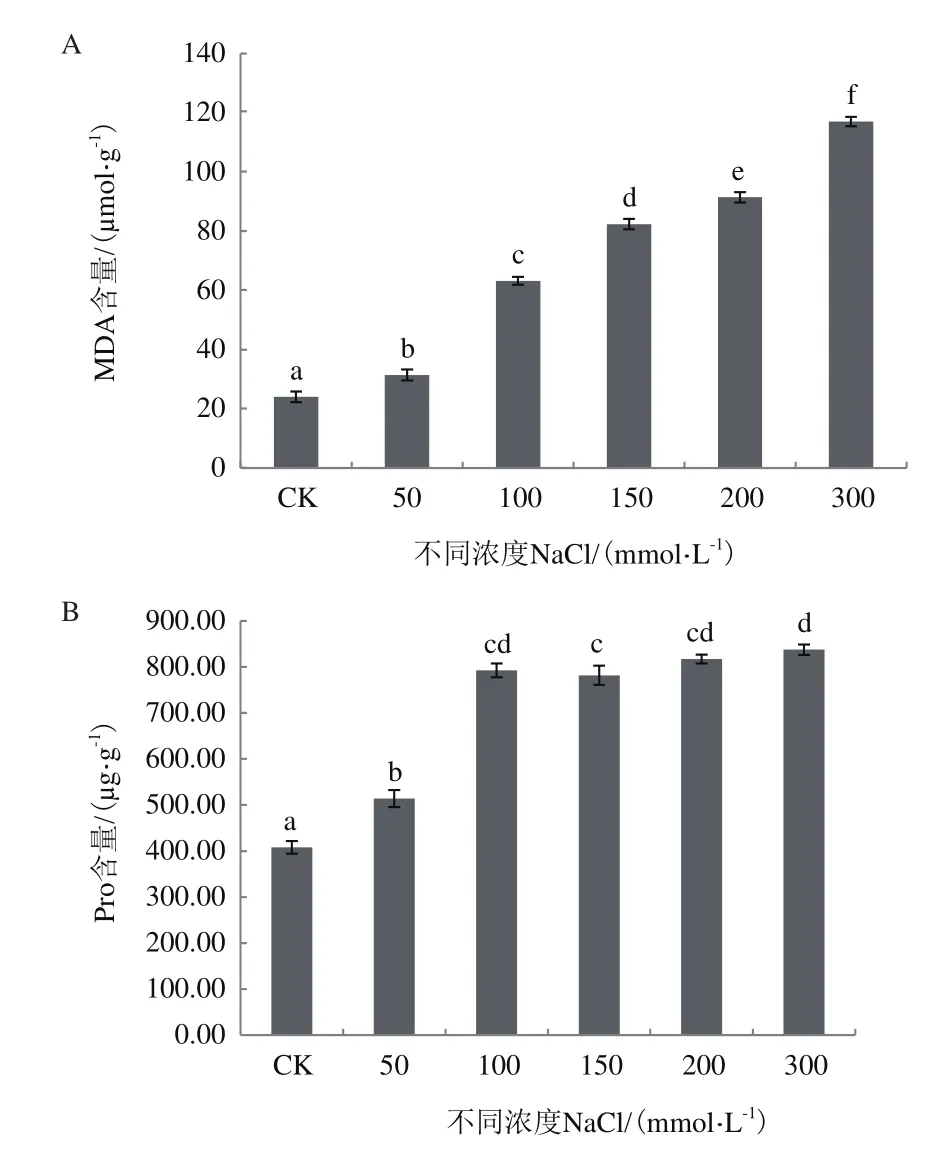
圖5 不同濃度NaCl處理下紅砂中的MDA和Pro含量變化
NaCl濃度增加的同時,POD的活性也整體呈現出增強趨勢,與CK相比,除了50 mmol/L外,其余濃度所對應的POD活性分別增加28.47%、52.18%、74.02%和81.41%,且增加顯著(P<0.05),但NaCl濃度為200和300 mmol/L所對應的POD活性差異不大,且NaCl濃度為50 mmol/L時,POD活性略有下降。
2.4 NaCl脅迫下紅砂萌發過程中耐鹽閾值的確定
分別以紅砂相對萌發率降為CK的50%時所對應的鹽濃度為萌發耐鹽閾值,植株全長降為對照的50%時所對應的鹽濃度為萌發后生長耐鹽閾值。
以NaCl濃度為自變量(x),以處理14 d后不同濃度下紅砂的相對萌發率、植株全長為因變量(y)建立回歸方程(表2),發現紅砂萌發耐鹽閾值濃度為273.04 mmol/L,生長耐鹽閾值為388.19 mmol/L,且擬合出的方程相關性較好。
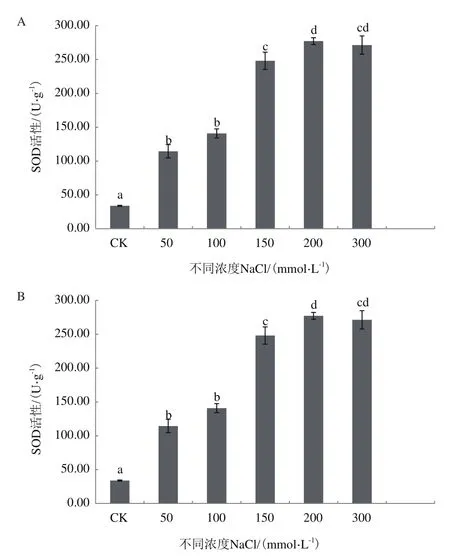
圖6 不同濃度NaCl處理下紅砂中的SOD和POD活性變化
與此同時,由于低濃度對于紅砂的萌發有一定的促進作用,分別以紅砂相對萌發率、植株全長為因變量y,NaCl濃度為自變量x,擬合出最適萌發曲線,經計算紅砂的最適萌發鹽濃度為45.78 mmol/L、最適生長鹽濃度為50.59 mmol/L。生長耐鹽閾值濃度明顯高于萌發耐鹽閾值濃度,最適萌發濃度和最適生長濃度差異性不大。
3 討論
鹽分是土壤組分的重要組成部分,對于植物的生長發育起著重要的作用,但過量的鹽分會導致植物種子萌發率降低、光合作用下降、活性氧代謝紊亂、蛋白質合成受阻等,最終造成植物生長發育遲緩[3-5]。本研究表明,低濃度的NaCl溶液處理,促進紅砂種子萌發,且萌發后的植株全長、根長、苗高較CK顯著增加,這種現象的出現可能是低濃度NaCl溶液促進細胞膜的滲透調節,或者是紅砂對低濃度鹽的一種適應機制,通過加速生長降低鹽濃度。
隨著NaCl溶液濃度的增加,紅砂的萌發時間滯后、相對鹽害率增加,相對萌發率、發芽指數、活力指數下降,植株全長、根長、苗高降低顯著,但根長的變化更為明顯,推測鹽脅迫主要影響紅砂根系建成與延長,影響水分吸收和養分供應;當鹽濃度為400 mmol/L時,紅砂不能萌發,但復萌處理發現,這些脅迫后的種子萌發迅速、集中,且脅迫35 d紅砂種子仍然具有萌發活力。這說明紅砂可以長時間忍受鹽堿環境,在鹽堿環境下,種子內部進行響應調節,以維持低耗、最佳萌發狀態(推測應為種子萌發的第Ⅱ階段),待條件適宜時,快速集中萌發。因此,在復萌階段,可將脅迫后待復萌種子進行生理生化指標分析,或者將待復萌的種子進行萌發期耐鹽基因篩選,以確定鹽生植物逆境條件下的“保活”機制。
通過擬合曲線分別確定了紅砂萌發期、生長期的耐鹽閾值及最適鹽濃度。從結果來看,紅砂萌發期的耐鹽閾值為273.04 mmol/L,萌發后生長耐鹽閾值為388.19 mmol/L,最適萌發鹽濃度為45.78 mmol/L,最適生長鹽濃度為50.59 mmol/L,萌發后生長耐鹽閾值明顯高于萌發耐鹽閾值,但最適生長和最適萌發鹽濃度差異性不大,這印證了低濃度鹽對鹽生植物的萌發和生長具有促進作用,也說明種子萌發階段是鹽生植物逆境條件下成活的關鍵階段,在這一階段,抗逆性最弱,對鹽脅迫比較敏感,種子萌發成功后對鹽環境適應性較強,可適應較高濃度的脅迫。萌發閾值及生長閾值的確定,為鹽生植物研究提供了不同發育層面的數據支撐。
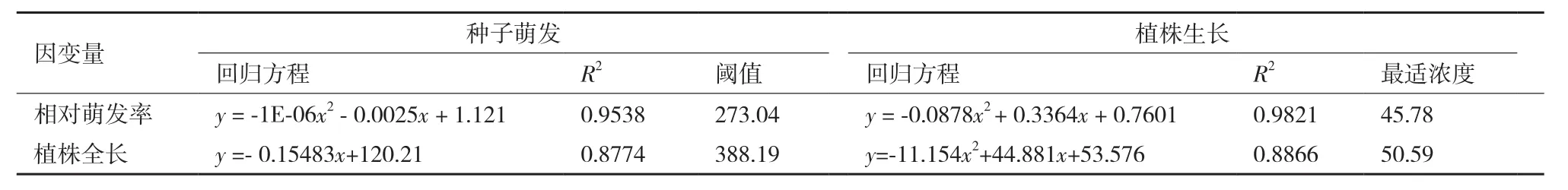
表2 NaCl條件下紅砂種子萌發及生長的耐鹽閾值與最適鹽濃度
當植株受到鹽脅迫時,植物體內活性氧代謝紊亂,MDA積累,細胞膜脂過氧化,細胞膜滲透性增大,細胞水分運輸失衡,同時Pro積累,起到滲透調節作用。本研究表明,隨著鹽濃度的增加,紅砂植株內MDA、Pro含量較對照顯著增加,但100、150、200和300 mmol/L之間Pro含量差異不大(P>0.05),可能是過高的鹽濃度導致植株僅依靠Pro不能進行有效的細胞膜滲透性調節,須有其他物質參與,共同響應并調節鹽脅迫下植株的各個過程。
植物在逆境下會積累MDA、活性氧(Reactive oxygen species,ROS)和自由基,導致膜脂過氧化、損傷大分子生命物質,進而導致生理生化紊亂。為了減少脅迫危害, 植株內的SOD 催化超氧化物的歧化反應,生成過氧化氫(H2O2)和氧氣(O2),但H2O2的積累對植株不利,需要POD 將SOD 歧化反應產生的H2O2分解[23-24]。結果表明,隨著鹽濃度的增加,SOD、POD的活性逐漸顯著增強,但200和300 mmol/L所對應的POD活性差異不大,推測原因為:鹽濃度過高,積累的較多的MDA,細胞膜滲透性發生了較大變化,MDA的積累激活SOD、POD相關信號通路,增加SOD、POD酶活性,加速歧化反應的進行,但SOD、POD相關信號通路對于MDA的響應存在一定的濃度區間。當MDA含量過低時,脅迫尚未造成直接影響,植株可通過自身滲透物質Pro進行調節,不激活酶活信號通路;當MDA濃度過高,超出植株自身調節范圍時不能激活相關信號通路,或是能激活相關信號通路進行調節,但歧化反應的發生需要消耗能量,種子內的儲存的物質有限,僅能維持一定程度內的歧化反應,最終導致植株脅迫致死或脅迫假休眠(非鹽生植物種子脅迫致死,鹽生植物種子脅迫假休眠)。因此,探究與脅迫響應相關酶聯信號通路和脅迫下鹽生植物種子保活機制尤為必要。
綜上所述,鹽濃度的增加,會導致紅砂種子萌發受阻,萌發時間滯后;萌發后生長受到抑制,且受抑制明顯;擬合方程發現種子萌發后生長耐鹽閾值顯著高于萌發耐鹽閾值,同時MDA、Pro積累,POD、SOD活性增加。這說明萌發期是紅砂完成生活是的關鍵時期,這一時期受到脅迫在一定程度內可通過提高POD、SOD活性清除MDA,增加Pro含量減少脅迫危害,維持正常的生理代謝。但這些指標的變化僅能幫助確定紅砂萌發期對鹽堿逆境的生理響應,并不能完全解析鹽生植物紅砂萌發期的耐鹽機制。因此,應進一步以這些生長生理指標為基礎,分別以閾值濃度和最適濃度做紅砂萌發期差異基因分析,開展鹽生植物種子復萌及脅迫后種子保活機制研究,這將為深入研究脅迫下鹽生植物種子萌發期信號通路、耐鹽機制奠定基礎,也將為通過分子育種解決鹽堿地區植被更新困難奠定理論基礎和提供基因資源。
4 結論
適當低濃度NaCl促進紅砂萌發及萌發后生長,反之則抑制,且萌發期耐鹽性較萌發后生長階段更為敏感,在生長階段,高濃度NaCl對紅砂根長的抑制作用更為明顯。紅砂可在脅迫環境下長期保持萌發活力,待萌發條件適宜時,可快速集中大量萌發;紅砂通過提高Pro含量維持滲透勢,增加POD和SOD活性清除MDA以維持正常的生理代謝。
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
