擬南芥RALF多肽家族的功能多樣性初步分析
2019-01-23 12:05:06強曉楠李鑫陳佳廖紅東于峰
生物技術通報 2019年1期
強曉楠 李鑫 陳佳 廖紅東 于峰
(湖南大學生物學院,長沙 410082)
小肽可作為植物生長發育與逆境適應性等細胞活動的調控信號,賦予植物適應復雜多變環境的能力[1]。快速堿化因子(Rapid alkalinization factor,RALF)首先在煙草中發現并被鑒定為一類5kD左右的小肽分子[2]。部分RALF可使細胞外圍環境快速發生堿化反應[3-4]。在模式植物擬南芥、水稻、楊樹和玉米等生物體內,針對RALF同源基因的克隆與功能分析顯示,RALF在主根中都抑制細胞生長,并提出RALF主要是通過使細胞壁堿化,從而抑制細胞生長[5]。RALF蛋白前體通常具有80-120 aa[2,6]。通過某種蛋白酶,如 S1P 蛋白酶[7],切割獲得RALF成熟多肽(約50 aa),成熟RALF小肽富含半胱氨酸,并在N末端存在一段保守的-YISY序列,推測其對于識別RALF對應的受體蛋白起重要作用,若將這段序列中的異亮氨酸突變成丙氨酸,則顯著抑制RALF功能[8-9]。在擬南芥中,至少具有35個組織特異性表達的RALF成員[10]。RALF4和RALF19在花粉管中高表達,RALF4/19能抑制花粉管的破裂[11-12],但RALF4對根長和下胚軸并沒有抑制作用[4]。這些結果暗示,RALF不同成員的功能可能具有組織特異性和多樣性。
受體激酶FERONIA(FER)是 RALF1信號分子的受體。RALF1與位于細胞膜表面的FER受體激酶結合并促進FER的磷酸化,抑制氫泵AHA2的活性,導致細胞壁堿化,從而抑制主根的生長[13]。本課題組的工作進一步發現FER通過與受體激酶RIPK相互作用響應RALF1信號,并抑制主根細胞生長[14]。同時,RALF1-FER/RIPK信號通路也在逆境響應中發揮重要作用。課題組前期研究發現FER能夠抑制逆境激素脫落酸(Abscisic acid,ABA)的響應,并介導RALF1與ABA在主根生長調節中的交叉會話[15]。在生物免疫方面,以絲裂原活化蛋白激酶(MAPK)磷酸化分析及活性氧迸發(Reactive oxygen species,ROS)作為檢測指標[16-19]。RALF23作為配體小分子,與受體FER相互作用,抑制免疫方面相關的受體復合物(EFR/BAK1/FLS2)的形成,負調控免疫信號的應答。在擬南芥中,過表達RALF23可以抑制flg22誘導的ROS產生,并且 對PtoDC3000(Pseudomonas syringaepv. tomato DC3000)的敏感性增加,相反,其突變體可以促進flg22誘導的ROS的產生,并且對PtoDC3000的敏感性降低[10],以上研究表明RALF23作為負調節因子,通過FER參與到由病原體分子模式引起的植物細胞免疫。綜上可知,RALF的活性分析具有以下幾類主要測定指標:(1)對根長等組織細胞生長的影響;(2)對flg22誘導的ROS產生和(3)對MAPK磷酸化的影響。
目前的研究結論表明,RALF1作為FER的配體的遺傳學證據只在根中成立,多項研究都認為RALF抑制根和葉片組織細胞的生長,而FER在不同組織中對細胞生長起到不同甚至完全相反的作用,比如FER和RALF1都在根中抑制細胞生長,但是FER在葉片中促進細胞生長[20-21],這與RALF1功能相反。為解釋以上悖論,目前推測認為不同RALF多肽成員在不同組織存在功能差異,甚至存在完全相反的功能,而這些功能相反的成員在不同的組織中與特異的受體結合,從而在不同組織起到差異甚至相反的遺傳學功能。為了驗證該推測,本研究克隆并分析了19種RALF小肽,系統性地研究不同RALF對根生長及下胚軸伸長的影響,發現并不是所有RALF都會對下胚軸伸長有抑制作用,RALF10會明顯促進下胚軸的伸長。該實驗結果表明不同RALF確實存在功能性的差異,甚至出現相反的功能。
1 材料與方法
1.1 材料
1.1.1 主要試劑 限制性核酸內切酶BamH 1(貨號:00455366)和EcoR I(貨號:00466339)、高保真酶(貨號:P505-d1-AA)、T4 DNA連接酶(貨號:00524509)為Thermo Fisher Scientific公司產品。氯化鈉(貨號:7647-14-5)、蛋白胨(貨號:73049-73-7)、酵母粉(貨號:8013-01-2)、Tris-堿(貨號:77-86-1)、異丙基 -β-D-硫代半乳糖苷(IPTG)(貨號:367-93-1)、咪唑(貨號:288-32-4)、卡那霉素(貨號:70560-51-9)以及氨芐霉素(貨號:69-52-3)均購于生工生物工程上海(股份)有限公司。載體與菌株:菌株E.coliDH5α,E.coliBL21由本實驗室保存,質粒pET-32a由本實驗室保存。
1.1.2 主要材料 擬南芥Col-0(哥倫比亞背景),將消毒好的Col-0點種于1/2 MS固體培養基上,垂直置于長日照培養箱里(16 h光照,8 h黑暗),溫度為22±4℃,4-5 d后取材進行根長處理及下胚軸處理實驗,7 d后取材進行MAPK磷酸化分析實驗。移苗于營養土中的Col-0在擬南芥生長室生長4-5周后取材進行ROS含量測定實驗。
1.2 方法
1.2.1 生物信息學分析 從Tair網站上獲得(https://www.arabidopsis.org/)RALF多肽氨基酸一級序列。使用帶有默認參數的ClustalW軟件對成熟的RALF多肽序列進行多序列比對。進化樹分析采用鄰近-連接方法,使用MEGA7.0軟件(參數為1 000 bootstrap 和 Poisson correction model)對多序列比對后的RALF多肽進行進化樹分析。
1.2.2 蛋白表達載體的構建 本研究中所選的表達載體為pET-32a,首先利用Primer 5.0軟件設計特異性引物,以野生型擬南芥cDNA為模板,進行PCR克隆擴增,獲得目的基因片段,設計的引物見表1。PCR 程序為 95℃ 3 min;95℃ 15 s,58℃ 15 s,72℃ 40 s,共36個循環;72℃ 5 min,22℃ ∞。然后進行瓊脂糖凝膠電泳回收目的條帶,以BamH I和EcoR I為酶切位點,對擴增產物和載體進行雙酶切(37℃,2 h)。然后用T4連接酶 4℃連接12 h后進行熱激轉化(42℃,90 s),涂布于含有氨芐霉素(Amp+)抗性的LB固體培養基上,37℃培養12 h,然后挑取單菌落進行菌落PCR檢測,挑取陽性菌落進行搖菌(37℃,12 h),提取質粒并測序。
1.2.3 蛋白誘導 將測序正確的重組質粒經過熱激轉化至宿主菌BL21中,涂布于含100 mg/mL氨芐霉素的LB固體篩選培養基上,37℃培養12-16 h后,挑取單菌落接種于6 mL含100 mg/mL的氨芐霉素的LB液體培養基中,37℃,220 r/min,活化10 h。次日將活化的菌液按1∶100的比例接種于100 mL 含100 mg/mL的氨芐霉素的LB液體培養基中,37℃,220 r/min振蕩培養2-3 h,至OD600=0.8,加入0.5 mmol/L的IPTG,28℃誘導6 h,離心收集菌體(5 000 r/min,22℃,5 min)。
1.2.4 蛋白純化 將菌體重懸于10 mL裂解緩沖液(50 mmol/L Tris-HCl(pH8),150 mmol/L NaCl,10 mmol/L imidazole,1 mmol/L PMSF),置于冰上,20 kH,5 min,超聲破碎,離心(5 000 r/min,4℃,5 min)收集上清,加入 500 μL Ni-NTA Agarose,4℃孵育12 h。離心(800 r/min,4℃,1 min)去上清,加入清洗緩沖液(50 mmol/L Tris-HCl,300 mmol/L NaCl,20 mmol/L imidazole),4℃孵育 10 min,重復清洗兩遍后去上清,加入洗脫緩沖液(50 mmol/L Tris-HCl,250 mmol/L imidazole),4℃孵育 1 h。收集上清(800 r/min,4℃,1 min),得到RALF蛋白。取少量純化重組蛋白,11% SDS-PAGE凝膠電泳,考馬斯亮藍染色觀察。
1.2.5 根長抑制及下胚軸抑制實驗 將擬南芥Col-0種子在75%酒精里消毒 1 min,無菌水沖洗兩次,然后加入15%的次氯酸鈉消毒 3 min,無菌水沖洗3次,將消毒好的種子置于4℃,春化2 d。根長抑制實驗,將消毒好的種子點種于1/2 MS固體培養基上,放置在22℃長日照培養箱中,使其生長4 d,然后將苗子移放到含有RALF小肽(1 μmol/L)的液體1/2 MS液體培養基中,正常生長3 d后進行根長觀察和統計。下胚軸抑制實驗,將春化后的種子點種于1/2 MS固體培養基上,使其生長4 d,然后將苗子移放到含有RALF小肽(1 μmol/L)的液體1/2 MS液體培養基中,放在黑暗條件下培養,4 d后進行下胚軸觀察和統計(對照組平均值-實驗組平均值/對照組平均值)。對照組植物與實驗組植物處于相同的生長條件。使用Image J軟件分別統計根長及下胚軸數據。
1.2.6 flg22誘導的ROS實驗 剪取未抽薹的野生型(Col-0)葉片(第七、八片對生葉),將葉片切成直徑為0.4×0.4 cm的正方形小塊,分別轉入每孔含有100 μL雙蒸水(pH5.8)的96孔板,用錫箔紙遮蓋,避光,過夜反應(8-12 h),消耗因機械損傷引起的ROS,并恢復葉片生理狀態。第2天,吸去雙蒸水,加入反應液 17 μg/mL luminol(Sigma,貨號 CAS :521-31-3),10 μg/mL HRP(Sigma),0.1 μmol/L flg22(Sigma),1 μmol/L RALF,陽性對照加入 flg22,實驗組加入flg22及相應的1 μmol/L RALF小肽,然后迅速放入儀器Fluoroskan Ascent FL(Thermo Scientific),檢測。
1.2.7 MAPK磷酸化水平分析實驗 每組稱取0.1 g生長了7 d的擬南芥幼苗,置于含有1/2 MS液體培養基的1.5 mL EP管中(實驗組含2 μmol/L RALF小肽),置于光照培養箱處理5 min,15 min,30 min,去除反應液進行蛋白提取,蛋白提取試劑為(150 mmol/L NaCl,10 mmol/L MgCl2,50 mmol/L HEPES(pH7.5),10 mmol/L KCl,5 mmol/L EDTA,1%Tritonx-100,10% glycerol,1% PMSF,1% Cocktail),Western 檢測。用 β-actin(CMC,貨號:AT0004)檢測上樣平衡,使用p-MAPK(Cell Signaling,貨號:#4370)抗體檢測MAPK磷酸化水平。WB灰度值使用image J 軟件進行測定。
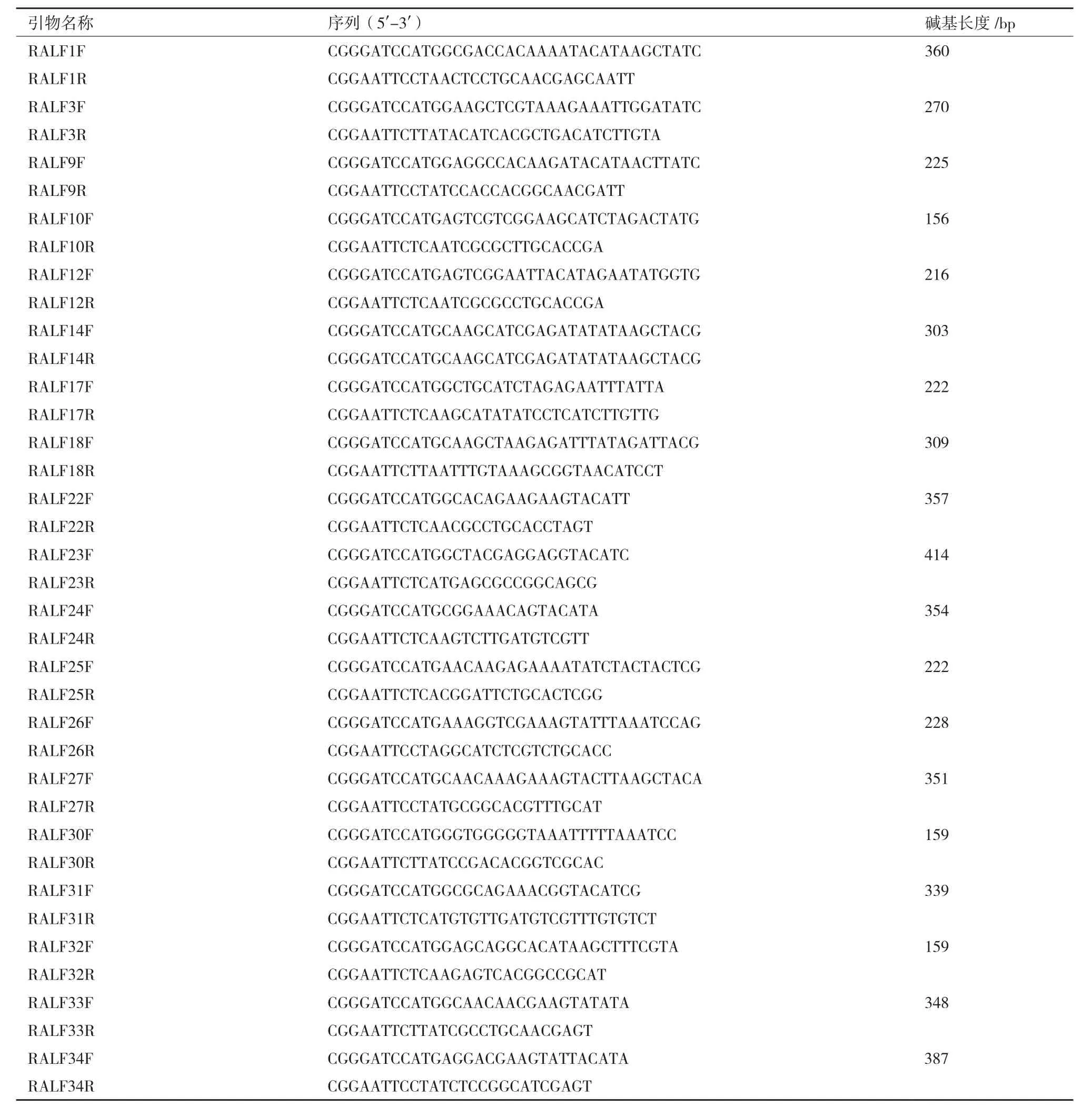
表1 RALFs引物
2 結果
2.1 重組His-RALF的純化
在擬南芥中,RALF家族至少含有35個成員,并有著不同的分布與功能,為系統性研究這些RALF的功能,首先根據其蛋白一級結構序列上的相似性,將其分為四大類并分別命名為I、II、III和IV。
本研究中,我們選擇具有代表性的19個RALF進行后續的研究,研究的RALF蛋白當中,以RALF1為代表的第I類共8個成員中我們選取其中6個成員進行研究;以RALF34為代表的第II類共9個成員中我們選取其中7個成員進行研究;以RALF17為代表的第III類共14個成員中我們選取其中3個成員進行研究;以RALF18為代表的第IV類共4個成員中我們選取其中3個成員進行研究(圖1-A)。
首先對這19個RALF進行目的片段的PCR克隆,亞克隆進入原核蛋白表達載體并轉入BL21大腸桿菌中進行原核表達和誘導,純化并獲得原核表達的重組蛋白。通過考馬斯亮藍染色分析,發現RALF目的蛋白是純化后蛋白的主要成分,分子量與預測大小吻合,表明RALF純化成功(圖1-B)。
2.2 RALF抑制主根生長實驗的分析
RALF對主根生長的調控是其活性的主要體現指標之一。已有研究報道發現RALF1處理能夠有效的抑制主根的生長,后續的研究發現RALF23同樣也能夠有效的抑制主根的生長。為了驗證除RALF1和RALF23外的其他17種RALF是否都具有抑制根長的作用,我們用1 μmol/L RALF處理擬南芥野生型Col-0(WT)幼苗,發現所有RALF均能不同程度地對根的生長產生抑制(圖 2)。從實驗結果來看,不同的RALF對根長的抑制效果存在一定的差異,RALF1/23均能對根的生長產生抑制,其抑制率分別為26.42%和18.56%(圖 2),該結論與前期研究報道的實驗結論一致。
RALF27對根生長的抑制作用相對于其它RALF最低,只有7.46%的相對抑制率。而RALF34的相對抑制率達到了26.63%,對根的抑制作用最明顯。以上實驗結果表明,這19種RALF均能對根的生長產生抑制,不同的RALF對根的抑制作用存在一定的差異。

圖1 RALF家族成員進化分析及蛋白純化
同時,進一步探究在fer-4背景下,不同RALF處理對根生長的抑制作用。fer-4為T-DNA插入到FER胞外域后得到的突變體植株,相比于野生型,其植株矮小,根毛缺失,表皮毛減少,并且結實率顯著降低,fer-4突變體根長生長對RALF1和RALF23不敏感。fer-4實驗結果表明,在fer-4背景下,RALF對根生長的抑制與WT存在差異。與前期研究的結論一致,RALF1和RALF23在fer-4背景下,和WT相比對根生長的抑制不敏感,RALF1抑制率為9.65%,RALF23抑制率為7.63%(圖 2),表明這些RALF對根生長的抑制可能由FER介導。
而RALF34抑制率為25.82%,RALF10抑制率為25.34%,和對照WT的抑制率大體類似,表明這些RALF介導的對根生長的抑制可能由其它受體激酶介導。以上實驗結果表明,RALF可能與其特異特定受體蛋白結合介導對主根生長的抑制。

圖2 RALF對Col-0及fer-4主根生長的抑制
2.3 RALF誘導MAPK磷酸化的分析
RALF對絲裂原活化蛋白激酶(MAPK)磷酸化水平的調控是其生物活性的另外一個體現指標。活性氧迸發(ROS burst)和MAPK磷酸化是病原體觸發的植物細胞免疫(PTI)引起的兩個早期事件。為研究RALF是否也參與到植物細胞免疫的信號通路,我們以MAPK磷酸化變化來驗證RALF在植物細胞免疫調節中的作用。前期的研究表明,真菌內分泌的RALF類似小肽可激活MPK3/6的磷酸化。
基于此,本研究分別從每一大類中選取3個RALF進行MAPK磷酸化水平分析實驗。實驗結果(圖3)表明,RALF9/10/14/18/23/30/32/33/34處理Col-0幼苗5 min后,MPK3/6的磷酸化顯著增強,RALF1/3在處理15 min后,MPK3/6的磷酸化顯著增強,然而,隨著處理時間的增加,MAPK磷酸化顯著下降,表明RALF1/3/9/10/14/18/23/30/32/33/34可能參與到了由MAPK介導的植物細胞免疫,這種由RALF1/3/9/10/14/18/23/30/32/33/34參與引起的MAPK磷酸化是一種快速的響應機制。而RALF17對MPK3/6的磷酸化沒有顯著性的影響(圖3-B)。
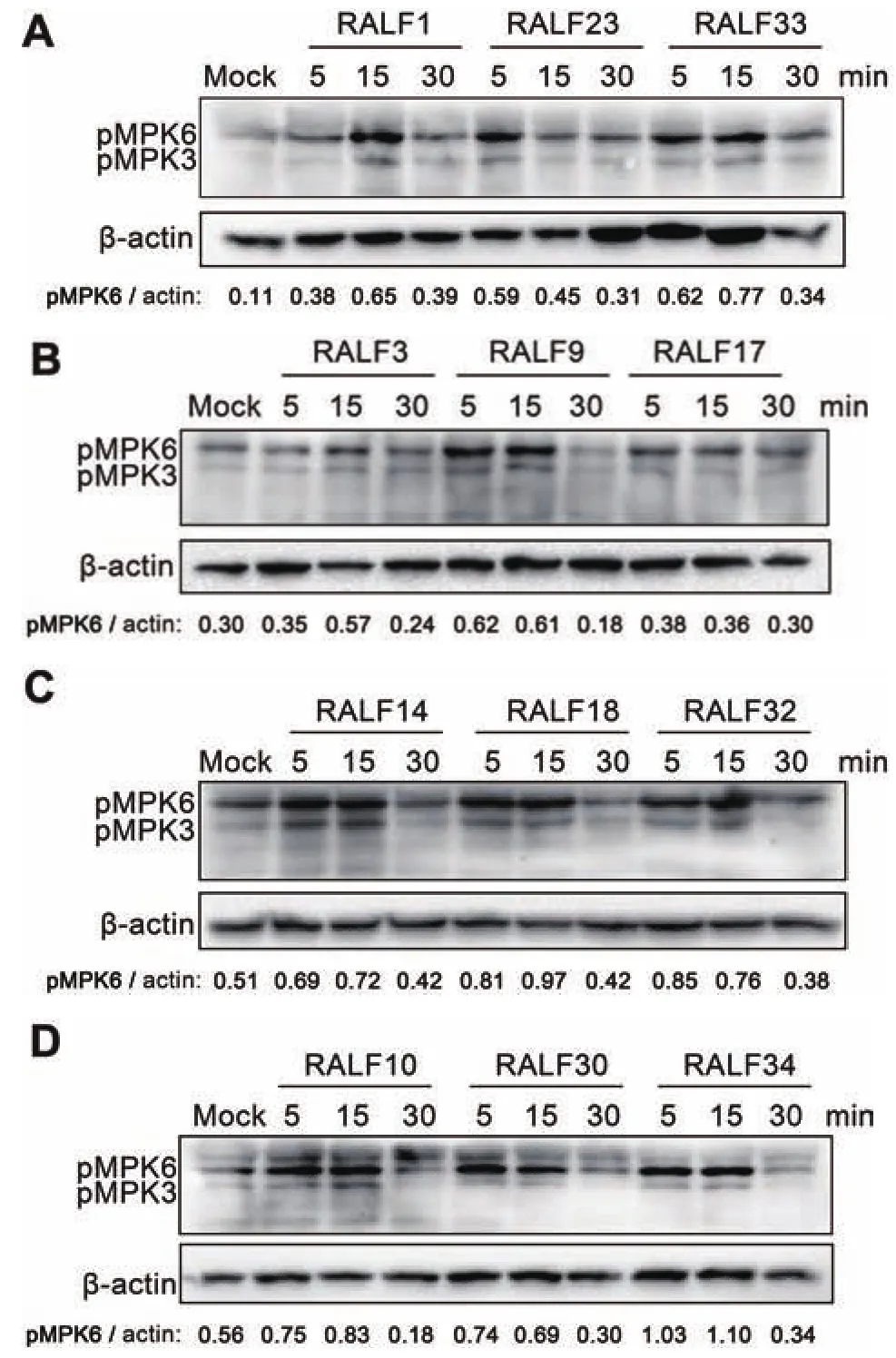
圖3 RALF對MAPK磷酸化的影響
2.4 RALF對flg22誘導的ROS生成影響分析
RALF可以通過調節flg22誘導的ROS來調控植物的免疫系統,是RALF活性的一個重要指標。ROS,作為一種第二信使分子,參與到對植物細胞免疫的響應,病原體分子模式誘導植物細胞內ROS的釋放。
圖4 RALF調控flg22觸發的ROS釋放
基于此,本研究想進一步探索不同的RALF對病原體分子模式引起的ROS的釋放的影響。在含有flg22的反應體系中添加不同RALF,探索RALF對flg22誘發的ROS釋放的影響。實驗結果(圖 4)表明,分析的16種RALF(RALF1/9/10/12/14/18/22/23/24/25/26/27/30/31/33/34)均不同程度地抑制由flg22誘導的ROS的釋放。表明其可能在植物細胞免疫中起負調控作用。
2.5 RALF對下胚軸伸長的影響分析
FER突變后,在黑暗生長情況下,下胚軸變短,這暗示RALF可能參與下胚軸細胞的伸長調控過程,但是目前尚未有相關研究報道RALF對下胚軸伸長的作用。基于此,本研究想研究這19個不同的RALF對下胚軸伸長的影響。分別用1 μmol/L不同的RALF處理擬南芥幼苗,置于黑暗培養箱培養4 d后觀察實驗結果,實驗結果(圖 5)表明,不同的RALF對下胚軸的抑制存在極大的差異,與對照組相比,RALF3和RALF9的抑制作用相對不明顯,抑制率分別為12.86%和11.51%。RALF10則是促進下胚軸的伸長,其促進伸長的百分比為41.99%,其它RALF則均能顯著的抑制下胚軸的伸長。
該實驗結果表明,不同的RALF參與對下胚軸伸長抑制存在功能性的差異,大部分的RALF能夠顯著抑制下胚軸伸長,但也有少部分的RALF則能促進下胚軸的伸長,這暗示不同RALF的功能存在組織差異性。
3 討論

圖5 RALF對Col-0下胚軸伸長的影響
RALF小肽最早從煙草中發現并提取,它可以引起煙草細胞懸浮液的堿化、并可以激活體內的MAPK反應、引起Ca2+流的上升。同時,RALF同源物也從多個物種中被分離出來,如,雙子葉植物,單子葉植物,裸子植物等[5]。RALF作為進化保守的多肽分子,以基因家族形式存在,在植物不同器官的表達模式存在差異,這暗示RALF不同成員之間功能可能具有功能特異性,可能影響特定的信號通路。本研究的實驗結果發現,不同的RALF對下胚軸細胞的伸長過程起到截然相反的作用,RALF1,23等抑制下胚軸細胞伸長,而RALF10則促進下胚軸細胞伸長。最近的研究發現,RALF 4和RALF19作為BUP1/2和AUX1/2的配體,在維持花粉管的穩定中起著非常重要的作用,當花粉管靠近珠孔時,從雌配子體釋放的RALF34與RALF4/19競爭BUP1/2和ANXUR1/2受體而干擾其信號,從而導致花粉管的破裂,釋放精細胞完成雙受精[12]。這讓我們猜想到,FER在不同組織中其功能存在差異是否也涉及類似具有不同功能的RALF成員之間的競爭結合,最終產生截然相反的功能。基于以上的研究,我們有理由猜測FER在不同部位具有相反功能的原因之一可能是由于FER與不同的RALF小肽結合從而引起下游不同的生物學響應。
基于蛋白一級結構序列進行分類的RALF成員在參與到相同的信號響應過程中存在很大差異,這表明,對RALF多肽的生物學功能研究從一級結構層面進行研究的策略還需要調整,如需要更多結構學研究來系統闡述一級序列相似,而功能不同的RALF成員是否具有結構差異性。此外,RALF多肽具有生物學活性需要其從前體肽切割釋放出成熟的功能性多肽,而這種從前體肽到成熟肽的加工過程需要特異的蛋白酶的參與和介導,是否存在除了S1P之外的的蛋白酶對不同的RALF前體肽進行切割,從而產生相應的生物學功能,這可能是今后需要關注和研究的方向。為了更為具體的研究不同RALF多肽的生物學功能,建議可以從基因敲除的層面入手,對特定的RALF進行特定的研究,從而從遺傳學的角度探明不同RALF的生物學功能。目前我們的工作還不完善,更多更深入的研究還需要開展,同時需要把重點落實到遺傳學層面,從而確認不同RALF的生物學功能和生物學意義。此外,關于不同RALF調控不同信號轉導的研究也是今后的研究方向和重點。
4 結論
本研究以模式生物擬南芥為研究材料,分別克隆出了19個RALF多肽,發現19個RALF均能抑制擬南芥主根的生長;RALF1/3/9/10/14/18/23/30/32/33/34影響MAPK的磷酸化變化;RALF1/9/10/12/14/18/22/23/24/25/26/27/30/31/33/34抑制由flg22引起的ROS的釋放。下胚軸伸長抑制實驗,發現RALF10促進下胚軸的伸長,推測RALF10可能通過不同于其他RALF的信號通路參與到對下胚軸伸長的調控。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
文苑(2018年22期)2018-11-19 02:54:14
中國科技論壇(2017年7期)2017-07-25 08:49:53
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
