消旋-3-正丁基苯酞對溴化乙錠誘導脫髓鞘模型作用及其機制研究
2019-01-11 07:54:52吳月娟廖宇晗
中風與神經疾病雜志 2018年12期
關鍵詞:手術
董 樂, 吳月娟, 黃 琪, 廖宇晗, 韋 雷, 吳 原
髓鞘由蛋白質層和磷脂層構成,髓鞘堿性蛋白(MBP)是中樞神經系統(CNS)髓鞘的主要蛋白質,位于髓鞘漿膜面,維持CNS髓鞘結構和功能的穩定,具有神經組織特異性[1]。研究表明少突膠質細胞(oligodendrocytes,OLs)缺失是脫髓鞘疾病的核心機制[2,3]。OLs起源于胚胎發育的早期階段,主要位于側腦室(LV)、第四腦室的室下區(SVz)和脊髓神經管腹側[4,5]。人們認為線粒體功能障礙和細胞凋亡可能在脫髓鞘過程中發揮更大的作用[6],消旋-3-正丁基苯酞(dl-3-n-Butylphthalide,NBP)可改善脫髓鞘的癥狀[7]。可能的機制是通過抗細胞凋亡作用保護髓鞘[8]。本研究通過注射溴化乙錠(ethidium bromide,EB)至SD大鼠側腦室中制作脫髓鞘模型,給予NBP干預,觀察大鼠的脫髓鞘程度和病理學改變,探討NBP在脫髓鞘病變中的作用機制,為臨床治療提供依據。
1 材料與方法
1.1 材 料
1.1.1 實驗動物及分組 正常成年健康雄性Sprague-Dawley大鼠36只(9~10周齡,體重200~250 g,SPF級),由廣西醫科大學實驗動物中心提供。所有大鼠在室溫22 ℃~25 ℃的自然光照下進行濕度控制的室內飼養,并自由進食和飲水。并隨機分為3組:EB組(n=12)、NBP組(n=12)和假手術組(n=12)。
1.1.2 主要試劑 兔抗鼠MBP多克隆抗體、兔抗鼠cytochrome C多克隆抗體、兔抗鼠caspase-9多克隆抗體購于美國Abcam公司;辣根過氧化物酶標記山羊抗兔二抗購于碧云天公司;胞漿蛋白抽提試劑盒、ECL化學發光試劑盒、免疫組織化學試劑盒、LFB髓鞘染色試劑盒、DAB顯色試劑盒購于索萊寶公司。
1.2 方 法
1.2.1 EB誘導脫髓鞘模型的建立 所有大鼠腹腔注射10%水合氯醛(3 ml/kg)麻醉,固定于立體定位儀。EB組和NBP組:用微量注射器以3 μl/min的速度把5 μl 濃度為0.05%的溴化乙錠溶液通過直接單次注射的方式注射至右側側腦室,并留針5 min以保證藥物充分彌散。具體坐標為(AP=-0.6;ML=+1.3;DV=-3.8)。假手術組:通過相同的方法注射5 μl生理鹽水。縫合頭皮,復溫蘇醒后回籠飼養。NBP組中的大鼠以腹腔注射的方式給予NBP干預,劑量為20 mg/(kg·d),EB組和假手術組以相同的方式注射相同量的生理鹽水。
1.2.2 動物取材和石蠟切片的制備 各組大鼠在14 d后用10%水合氯醛麻醉,打開胸腔暴露心臟,經左心室快速灌注生理鹽水,后改用4%多聚甲醛灌注進行前固定,然后斷頭取腦放于4%多聚甲醛進行后固定。常規石蠟包埋,進行3 μm連續冠狀位切片。
1.2.3 牢固藍(LFB)染色 常規脫蠟至水,95%酒精稍洗;Luxol Fast Blue染液室溫過夜,95%酒精清洗,蒸餾水沖洗,分化液分色15 s,蒸餾水沖洗,伊紅復染,蒸餾水沖洗,酒精脫水,二甲苯透明,中性樹膠封片,光學顯微鏡下觀察照相。通過脫髓鞘評分標準對各組大鼠進行脫髓鞘評分:正常髓鞘結構無脫髓鞘1分;輕度或輕度脫髓鞘(保留>50%髓磷脂染色)2分;重度脫髓鞘改變(髓鞘染色<50%保存)3分。記錄各組分值。
1.2.4 HE染色 切片常規脫蠟至水;蘇木素染色5 min,水洗10 min;1%鹽酸酒精30 s,水洗30 s;0.5%伊紅液染色3 min,蒸餾水洗30 s;酒精脫水、二甲苯透明、中性樹膠封片,光學顯微鏡下觀察照相。
1.2.5 Tunel法 常規脫蠟至水,PBS洗;蛋白酶K工作液37 ℃孵育,PBS洗;Tunel 反應混合液孵育,PBS洗;POD工作液孵育,PBS洗。DAB顯色,酒精脫水,二甲苯透明,中性樹膠封片,光學顯微鏡下觀察照相。選擇5個具有代表性的視野,計數陽性細胞數和陰性細胞數,并計算陽性表達百分率[陽性表達百分率=陽性細胞數/(陽性細胞數+陰性細胞數)×100%]。
1.2.6 免疫組織化學染色 常規脫蠟至水,PBS洗;0.3%H2O25 min,PBS沖洗后,放入檸檬酸緩沖液中進行高壓抗原修復20 min,取出,室溫下自然冷卻至35 ℃以下,PBS洗;正常羊血清室溫10 min;兔抗鼠MBP多克隆抗體(1∶200)、兔抗鼠Cyt C多克隆抗體(1∶5000)、兔抗鼠caspase-9多克隆抗體(1∶500)室溫30 min、4 ℃冰箱過夜、室溫復溫30 min, PBS洗;二抗37 ℃孵育60 min, PBS洗;DAB顯色;酒精脫水,二甲苯透明,中性樹膠封片,光學顯微鏡下觀察照相。免疫組織化學染色結果用Image Pro plus圖像分析軟件測定積分光密度(integral optical density,IOD)值和陽性面積(area)。計算出各組平均光密度(IOD/area)并記錄。
1.2.7 Western blot 取腦白質組織200 mg,依據胞漿蛋白抽提試劑盒說明書提取胞漿蛋白,煮沸變性,取50 μg蛋白,變性后行聚丙酰胺凝膠電泳,轉至硝酸纖維膜,TBST液洗3次,5%脫脂奶粉室溫封閉1 h,TBST液洗3次,入兔抗鼠MBP(1∶500)或Cyt C抗體(1∶250),4 ℃過夜,TBST液洗3次,加入辣根過氧化酶結合的抗兔IgG(1∶4000),TBST液洗3次,ECL化學發光試劑檢測,X線片顯影。采用條帶圖像分析系統測定各條帶灰度值。并以Image J軟件進行半定量分析,結果以相對光密度值×面積表示。蛋白的相對含量=目的蛋白條帶/β-tubulin條帶。記錄各組結果。
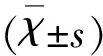
2 結 果
2.1 髓鞘LFB染色結果 假手術組灰質、白質界限明顯,染色深且均勻,髓鞘結構清晰、無脫失。與假手術組相比,EB組可見大小不等片狀髓鞘脫失,甚至出現大片的空泡變性脫失區域,NBP組藍色髓鞘區域相對完整,空泡變性區域相對減少(見圖1);與假手術組相比,EB組的LFB脫髓鞘評分明顯高于假手術組(P<0.05);與EB組相比, LFB脫髓鞘評分明顯低于EB組P<0.05) (見圖2)。
2.2 HE染色結果 假手術組細胞排列整齊有序,組織排列致密,形態結構正常,染色清晰;與假手術組相比,EB組組織排列紊亂,炎性細胞浸潤,白質結構松散、破壞;NBP組的無明顯的結構破壞,但結構較假手術組松散,存在少量空泡(見圖1)。
2.3 Tunel 法染色結果 EB組可見大量凋亡細胞,凋亡細胞數明顯高于假手術組,NBP組凋亡細胞較EB組減少(見圖1)。與假手術組相比,EB組凋亡細胞陽性表達百分率高于假手術組(P<0.05);與EB組相比,NBP組凋亡細胞陽性表達百分率明顯少于EB組(P<0.05)(見圖3)。
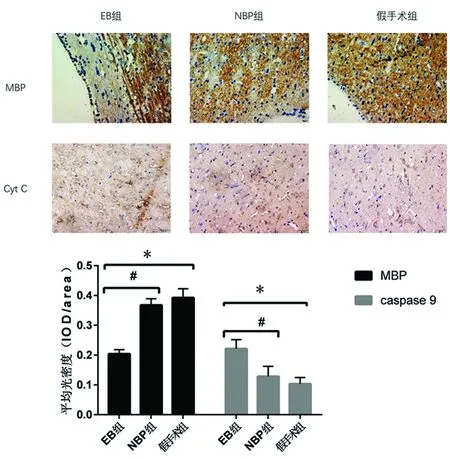
2.4 免疫組織化學染色結果 與假手術組相比,EB組MBP表達量明顯低于假手術組(P<0.05);與EB組相比,NBP組MBP表達量明顯高于EB組(P<0.05);與假手術組相比,EB組caspase-9表達量明顯高于假手術組(P<0.05);與EB組相比,NBP組caspase-9表達量明顯低于EB組(P<0.05)(見圖4)。
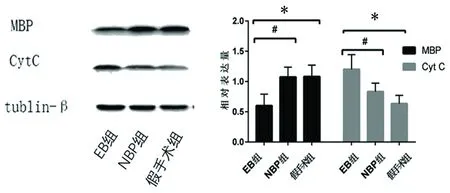
2.5 Western blot結果 與假手術組相比,EB組MBP表達水平顯著低于假手術組(P<0.05);與EB組相比,NBP組MBP表達高于EB組(P<0.05);與假手術組相比,EB組Cyt C表達水平顯著高于假手術組(P<0.05);與EB組相比,NBP組Cyt C表達水平低于EB組(P<0.05) (見圖5)。

圖1 各組大鼠腦白質區HE染色、LFB染色、TUNEL染色(10×40)

與假手術組相比*P<0.05;與EB組相比#P<0.05
圖2 LFB染色檢測各組大鼠腦白質脫髓鞘程度

與假手術組相比*P<0.05;與EB組相比#P<0.05
圖3 TUNEL染色法檢測各組大鼠腦白質陽性表達百分率
與假手術組相比*P<0.05;與EB組相比#P<0.05
圖4 免疫組織化學法檢測各組大鼠腦白質MBP、Cyt C的表達(10×40)
與假手術組相比*P<0.05;與EB組相比#P<0.05
圖5 Western blot 法檢測各組大鼠腦白質MBP、Cyt C的表達
3 討 論
EB誘導脫髓鞘模型是由Yajima和Suzuki在1979年研發出來,主要用于研究局灶性脫髓鞘疾病。既往研究發現,在2~4 w內該模型可以在目的區域產生明顯脫髓鞘病變,并且可見OLs死亡。OLs和星形膠質細胞丟失都是EB誘導脫髓鞘病變中心的標志[9]。建立EB誘導脫髓鞘模型后72 h內出現OLs死亡[10]。Mahdi Goudarzvand等指出EB具有誘導細胞凋亡的作用[11]。證明這個模型的穩定性。
MBP是特異性表達于髓鞘具有抗原性的蛋白質,是髓鞘蛋白的主要成分[12]。主要分布在胼胝體、海馬、以及小腦和皮質下區域。因此,MBP也被認為是成熟髓鞘的標志性蛋白[12]。只有成熟的OLs大量表達MBP,才有對軸突髓鞘化的能力[13],脫髓鞘病變時,病變區髓鞘崩解脫失,組織結構松散,炎性細胞浸潤, MBP表達減少。本實驗在EB側腦室注射后14 d, EB組出現髓鞘淡染,髓鞘崩解,髓鞘脫失;組織結構松散,神經纖維紊亂,炎性細胞浸潤,MBP表達減少。與EB組相比,NBP組髓鞘染色深染,髓鞘相對完整,髓鞘脫失減輕。組織結構完整,纖維排列整齊,炎癥細胞減少。NBP組MBP表達增高,表明EB誘導CNS脫髓鞘模型建立成功,而且NBP能夠減輕組織脫髓鞘和炎癥反應,保護髓鞘完整。
在CNS內,正常的軸突運輸過程和神經元存活依賴于適當的髓鞘形成[14]。有髓神經纖維的髓鞘由OLs的突起形成。在脫髓鞘過程中存在大量OLs凋亡[15],對12例因急性復發而死亡的MS患者早期病變形成的研究顯示,在沒有外周免疫細胞浸潤的情況下,病變包含廣泛的OLs凋亡[16],因此,抑制OLs凋亡過程能夠有效減輕脫髓鞘程度。Cyt C是12.3 kDa,核DNA編碼的蛋白質。在細胞質中游離核糖體合成[17]。Cyt C正常時位于線粒體內外膜之間,不能通過線粒體外膜進入胞漿,脫髓鞘病變時,OLs線粒體功能障礙,線粒體通透性轉換孔(PTP)開放[18]。持續的病理性PTP開放誘導胞質流入線粒體基質,導致線粒體膜電位的喪失和離子梯度的失衡,膜電位去極化促進線粒體基質擴張,線粒體腫脹和膜破裂,導致Cyt C釋放[19]。Cyt C從線粒體釋放到胞質,此為線粒體介導凋亡途徑的關鍵步驟,與Apaf 1、ATP/dATP結合形成凋亡體,Apaf 1通過分子中的胱天蛋白募集域(CARD)與caspase-9前體原域中的CARD相互作用而募集caspase-9前體,caspase-9前體自我激活后活化下游的caspase-3,引起OLs細胞凋亡[20]。EB組CNS存在大量細胞凋亡,而且Cyt C、caspase-9細胞質表達明顯上調,這些都表明在EB誘導的CNS脫髓鞘模型中,存在OLs線粒體介導的細胞凋亡。與EB組相比,NBP組Cyt C、caspase-9表達減少,凋亡細胞明顯減少,表明NBP可以有效減少EB模型中Cyt C釋放,抑制caspase-9的表達,抑制OLs內線粒體途徑凋亡途徑,維持OLs生理功能和髓鞘的完整。本實驗證實NBP對EB誘導的CNS脫髓鞘有保護作用。可能與抑制OLs內線粒體介導的內源性凋亡有關。
猜你喜歡
環球時報(2022-12-23)2022-12-23 09:28:37
昆明醫科大學學報(2022年1期)2022-02-28 07:45:04
中老年保健(2021年11期)2021-08-22 03:13:36
昆明醫科大學學報(2021年2期)2021-03-29 07:42:46
河北畫報(2020年10期)2020-11-26 07:20:50
小學閱讀指南·低年級版(2017年1期)2017-03-13 20:07:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:47
中國醫療美容(2015年1期)2015-07-12 10:06:38
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:54
西南軍醫(2014年5期)2014-04-25 07:42:48
