持續自然降溫對不同甘蔗品種形態及生理指標的影響
2017-11-30 18:14:59李素麗史曉朋李志剛楊麗濤李楊瑞
江蘇農業科學 2017年20期
關鍵詞:形態
李素麗+史曉朋+李志剛+楊麗濤+李楊瑞
摘要:為篩選耐寒優良甘蔗品種,在廣西霜、雪、冰期最早、最長的縣份之一的資源縣,選用8個甘蔗品種進行試驗,在甘蔗成熟期降溫后期甘蔗受害最嚴重的時期采樣,觀察甘蔗在低溫過程中的形態和生理變化,結合相關性檢驗,采用隸屬函數法對8個甘蔗品種進行抗寒性綜合評價。試驗結果表明,不同甘蔗品種對長時間持續低溫的反應各異,品種間莖的水漬狀節數、綠葉數、錘度、糖分含量、莖尖褐變指數、葉綠素含量等指標均達差異明顯。根據隸屬函數法的分析結果,將供試品種劃分為3級,其中Ⅰ級高抗寒型品種2個,Ⅱ級中抗寒型品種2個,Ⅲ級低抗寒型品種4個。由結果可知,甘蔗的抗寒能力存在著品種間差異,選擇多個形態和生理化指標,采用隸屬函數法,綜合評估甘蔗工藝成熟期的抗寒性是可行的,可以較好地揭示指標性狀與抗寒性的關系,為耐寒甘蔗品種篩選及生產提供參考依據。
關鍵詞:甘蔗;抗寒性;形態;生理;隸屬函數;抗寒性評價
中圖分類號: S566.101 文獻標志碼: A 文章編號:1002-1302(2017)20-0098-06
甘蔗是喜溫作物,種植于熱帶亞熱帶地區,需要適宜的溫度、濕度才能正常生長。隨著割手密等育種材料中抗逆基因的導入,甘蔗種植已從熱帶地區擴大到亞熱帶和溫帶地區[1]。我國近年來氣候變化異常,出現極端最低氣溫,氣候變化不穩定性增加,寒冷災害更加嚴重,寒害和冷害使甘蔗生產和制糖工業生產受到嚴重阻礙,每年都會造成不同程度的經濟損失[1-6],低溫寒害嚴重影響甘蔗生產以及制糖工業和其他用糖企業的穩定發展。因此,研究甘蔗抗寒性對提高甘蔗抗寒能力具有重要的意義。
受凍害甘蔗主要表現為心葉先死亡呈黑色,然后是生長點死亡,葉片受害干枯呈灰白色,不能進行光合作用,蔗莖上部嫩芽受凍害后變為黑褐色,受凍害嚴重的蔗莖,組織全部或部分呈水漬狀,節間有紅褐色斑,有時節間開裂,蔗糖分逐漸水解,有酸腐味,莖內組織變質由上而下逐漸擴展,嚴重降低原料蔗品質[7-13],譚宗琨等將凍害對甘蔗的傷害癥狀分為重級、中級、輕級和基本正常4個等級[2,14]。
在甘蔗成熟期,比較容易遭受自然低溫傷害的一般是在其生長期或成熟期,從而導致蔗莖呈水漬狀,嚴重影響蔗糖生產。因此,研究甘蔗成熟期對低溫的忍受能力及機制,針對不同冷敏感型甘蔗抗寒性的差異研究,選擇抗寒性差異顯著的品種為研究材料,對于尋求甘蔗抗寒育種理論上的突破與創新具有重要意義。本研究對8個不同甘蔗品種在持續自然降溫后的形態和生理變化進行比較,再用隸屬函數法對其抗寒性的強弱進行評價和排序,為深入研究甘蔗抗寒機制、品種推廣以及常遭受寒害侵襲的蔗區選擇栽培品種提供參考依據。
1 材料與方法
1.1 試驗地概況
試驗區所在地資源縣位于廣西壯族自治區北部,地處110°13′~110°54′E、25°48′~26°16′N之間,屬亞熱帶季風濕潤氣候區。由于地形地貌復雜,境內小氣候多樣,具有明顯的山地氣候特征。全縣年均氣溫16.4 ℃,年均降水量 1 761.1 mm,年均日照時數1 307.6 h,年均無霜期 300 d,是廣西霜、雪、冰期最早、最長的縣份之一。試驗地位于資源縣城附近,土壤為栗鈣土,土層深厚,土質肥沃,有灌溉條件。試驗采取隨機區組設計,3次重復,單行區,行長12 m,行距 1 m,小區面積12 m2。供試甘蔗于2013年3月11日種植,下種量為16芽/m,試驗田的管理按照常規措施進行。
1.2 試驗材料
共8個甘蔗品種,詳見表1。
1.3 取樣時間和方法
于2014年2月4日(最低氣溫為5.2 ℃)采樣,每個品種隨機選取5個生長一致的甘蔗,取+1葉(甘蔗最高可見肥厚帶葉片)。
1.3.1 莖水漬狀節數的計算 肉眼看到有水漬狀現象的計算為產生水漬狀節。
1.3.2 甘蔗莖尖褐變指數的計算 根據莖尖顏色的深淺和組織軟硬的程度,將甘蔗莖尖褐變程度用褐變指數表示,分為5級:0級為生長點完好,顏色淺黃色,組織有一定硬度;1級為生長點有輕微褐色,組織還有一定硬度;2級為生長點有較深褐色,組織還有一定硬度;3級為生長點深褐色并擴展到幼葉部位,組織變軟;4級為生長點深度褐變并擴展到幼葉和生長點以下幼莖部位,組織變軟;5級為生長點及幼葉和生長點以下幼莖深度褐變腐爛。
1.3.3 綠葉數的計算 肉眼觀察葉片呈綠色,而且不萎蔫或干枯的部分為綠葉,整張葉片都是以上特征的計算為1張綠葉,有部分枯黃的葉片按綠色葉片占葉全長的比例計算綠葉數。
1.3.4 葉綠素含量的測定 用SPAD-502葉綠素測定儀測每個品種葉片中部葉綠素相對含量(SPAD值)(SPADL)。
1.3.5 蔗汁錘度和甘蔗糖分含量的測定 按張篤思的方法[15]測定:蔗汁錘度用比重計法,甘蔗糖分含量用二次旋光法。
1.4 數據處理與分析
采用Excel和SPSS統計軟件統計分析試驗數據,參照張文娥等的方法[16-19],計算隸屬函數值、平均隸屬度,分析評價品種抗寒性。隸屬函數公式如下:
隸屬函數=(Xi-Xmin)/(Xmax-Xmin)。
(1)
式中:Xi為指標測定值,Xmin、Xmax分別為所有參試材料某一指標的最小值、最大值。如果某一指標與抗寒性呈負相關,則用反隸屬函數進行轉換:
反隸屬函數=1-(Xi-Xmin)/(Xmax-Xmin)。
(2)
2 結果與分析
2.1 資源縣氣象資料和甘蔗田間生長情況
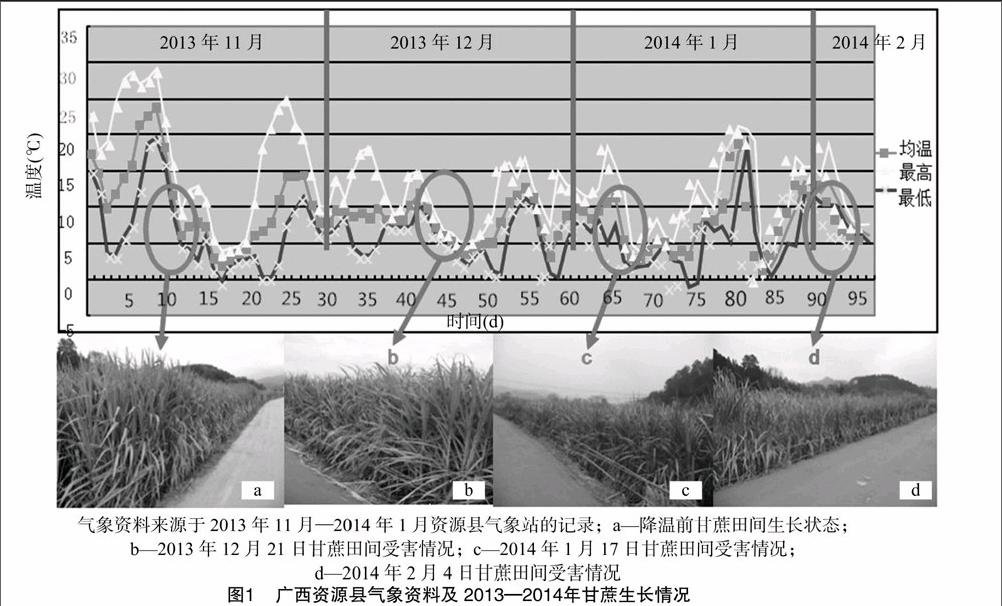
資源縣相關氣象資料顯示,從2013年11月中旬開始降溫,11月17日受較強冷空氣影響,出現雨夾雪天氣,降溫幅度較大,12月中下旬又出現低溫。2014年1月初再次出現寒潮天氣,一直持續低溫,出現當年日最低溫度-3 ℃(圖1)。降溫前,甘蔗生長狀況良好,葉片油綠(圖1-a),隨著降溫時間延長,甘蔗寒害程度加深,葉片枯黃(圖1-b~圖1-d),其中2014年2月4日甘蔗嚴重受害,大部分葉片枯黃(圖1-d)。endprint
2.2 不同甘蔗品種的形態表現
在2014年2月,廣西資源縣已達到當地的最低氣溫 -3 ℃,在種植的8個試驗品種中,發現大部分甘蔗品種已嚴重受冷害,從圖2可以看出,不同甘蔗品種的抗寒能力存在明顯差異,受害較輕的品種,綠葉數多,莖無水漬狀,莖尖無褐變現象;相反,受害較嚴重的品種,葉片枯黃,莖水漬狀嚴重,莖尖褐變也嚴重。
2.3 不同甘蔗品種水漬狀的差異
低溫會導致甘蔗細胞結冰,細胞脹破死亡產生組織水漬狀現象[20]。從圖3可以看出,不同甘蔗品種在降溫后期莖組織產生水漬狀現象的嚴重程度有明顯差異,本試驗中不同甘蔗品種中產生水漬狀現象的嚴重程度依次為GT28 2.4 不同甘蔗品種莖尖褐變指數的差異 低溫下發生的組織褐變是由于冰晶的增大使細胞膜結構破壞而打破了細胞內酚-酚酶區域化分布后,酚、酚酶和氧相互接觸,發生氧化褐變[21-23]。從圖4可以看出,不同甘蔗品種在持續降溫后期莖尖組織產生的褐變現象有明顯差異,供試品種中產生水漬狀現象的嚴重程度依次為GT28 2.5 不同甘蔗品種綠葉數的差異 葉片主要進行光合和蒸騰作用,綠葉數越多,光合能力就越強[24],綠葉數是鑒定植物抗寒性的重要指標之一[25]。從圖5可以看出,降溫后期品種間綠葉數達明顯差異,供試品種綠葉數按多少排序依次為GT28>GT21>GAN18>FN15>ROC22>TT98>YL8>YL6。由圖6可以看出,+1綠葉占全葉的比例按高低排序依次為GT28>GAN18>ROC22>TT98>GT21>FN15>YL8>YL6,其中GT28綠葉數最多,有5張,+1葉綠葉完整;而YL6最嚴重,無1張完整綠葉,+1葉也幾乎全部枯黃。 2.6 不同甘蔗品種葉綠素含量的差異 葉綠素含量是反映植物光合作用能力的間接指標,低溫會導致葉綠素含量降低[26-27]。從圖7可以看出,不同甘蔗品種在降溫后期葉綠素含量有明顯差異, 供試品種中葉綠素含量按高低排序依次為GT28>GT21>GAN18>FN15>ROC22>TT98>YL8>YL6,其中GT28葉綠素含量最高,而YL6葉綠素含量最低。 2.7 不同甘蔗品種蔗汁錘度的差異 錘度是指可溶性固形物含量的百分比,植物中可溶性固形物含量高能增強細胞滲透勢,降低細胞溶液的冰點[28]。從圖8可以看出,不同的甘蔗品種在降溫后期蔗汁錘度下降程度有明顯差異,供試品種中錘度降低嚴重程度依次為GT28 2.8 不同甘蔗品種蔗糖含量的差異 低溫脅迫下細胞蔗糖的積累有和錘度相同的生理功能,蔗糖含量是植物抗寒性研究中普遍采用的抗寒性指標之一[29-30]。從圖9可以看出,不同甘蔗品種在降溫后期糖分含量下降程度有明顯差異,不同甘蔗品種間差異明顯,供試品種中蔗糖含量降低嚴重程度依次為GT28 2.9 不同抗寒指標的相關性 自然降溫后不同甘蔗品種莖水漬狀節數、莖尖褐變指數、綠葉數、+1葉綠葉占全葉比例、葉綠素含量、錘度下降比例、蔗糖分下降比例等農藝性狀均表現出不同程度的差異,不同指標之間的相關性見表2。 從表2可以看出,莖水漬狀節數與莖尖褐變指數、錘度下降比例、蔗糖下降比例呈極顯著正相關,與綠葉數、+1葉綠葉占全葉比例、葉綠素含量呈顯著、極顯著負相關;莖尖褐變指數與甘蔗錘度下降比例、蔗糖下降比例均呈顯著正相關,與綠葉數、+1葉綠葉占全葉比例、葉綠素含量呈極顯著負相關。綠葉數與+1葉綠葉比例、葉綠素含量呈正相關,與錘度下降比例、蔗糖下降比例呈極顯著負相關;葉綠素含量與錘度下降比例、蔗糖下降比例呈負相關,錘度下降比例與蔗糖分下降比例呈極顯著正相關。莖水漬狀節數是衡量低溫傷害的比較直接的鑒定指標[5],分析結果表明,莖水漬狀節數與其他指標呈負相關,起負面效應,該指標越高,抗寒性越低。葉綠素含量、綠葉數、+1葉綠葉占全葉比例對甘蔗抗寒起正向作用,含量越高,甘蔗越抗寒;而莖尖褐變指數、錘度下降比例、蔗糖下降比例和莖水漬狀節數之間呈明顯正相關性。以上結果表明,低溫脅迫對甘蔗的影響是多方面的,莖水漬狀節數、莖尖褐變指數、綠葉數、+1葉綠葉占全葉比例、葉綠素含量、錘度下降比例、蔗糖下降比例可作為不同甘蔗品種抗寒性鑒定的重要指標。 2.10 利用隸屬函數評價甘蔗抗寒性 采用模糊數學中隸屬函數法,以水漬狀節數、莖尖褐變指數、綠葉數、+1葉綠葉占全葉比例、葉綠素含量、錘度下降比例、蔗糖下降比例等7個農藝性狀指標對8個甘蔗品種抗寒性進行綜合評價,不同甘蔗品種的平均隸屬度見表3。參照前人研究方法[16-18]結合大田實際情況,按照平均隸屬度將抗寒性分為3級,Ⅰ級:0.60~1.00,為高抗寒品種;Ⅱ級:0.30~0.59,為中抗寒品種;Ⅲ級:0~0.29,為低抗寒品種。8個品種的耐寒能力大小依次為:GT28>GT21>FN15>GAN18>YL8>TT98/0432>ROC22>YL6,其中桂糖28號最耐寒,園林6號最不耐寒。 3 討論與結論 溫度是植物生長發育過程中十分重要的生態因子,在低溫環境中,植物形態和生理等方面都發生不同程度的改變[31-32]。本研究結果表明,持續自然降溫后甘蔗形態和生理等發生很大變化,不同甘蔗品種莖的水漬狀節數、莖尖褐變指數、綠葉數、葉綠素含量、錘度和蔗糖在冷害前和冷害后的差值均有明顯差異,表明不同甘蔗品種受冷害的程度不一樣,甘蔗抗寒能力存在品種間的差異。
組織呈水漬狀的原因是由于溫度低到凍結狀態時細胞間隙的水分結冰,使細胞原生質的水分析出,冰塊逐漸加大致使細胞脫水,或使細胞解離死亡而造成的[20,33]。組織水漬狀的程度是鑒定植物抗寒性的重要指標之一[5]。本研究結果顯示,自然降溫后抗寒性較強的甘蔗品種桂糖28號、桂糖21號組織水漬狀節數較少,其次為中抗品種葉片;而抗寒性較弱的甘蔗品種ROC22、園林8號,園林6號水漬狀嚴重。表明桂糖28號最耐寒,而園林6號最不耐寒。
組織褐變是植物細胞酚、酚酶、氧接觸后產生的現象[21-23,34]。低溫環境下,一方面超氧化物歧化酶(SOD)等一些清除自由基的酶活性下降,體內自由基產生和清除平衡受破壞,導致丙二醛的積累,破壞膜的結構[35];另一方面由于霜凍導致組織結冰,對細胞組織產生傷害,膜結構的破壞酚、酚酶和氧相互接觸,發生氧化褐變[21,36]。組織褐變是細胞膜受傷的產物,其褐變程度是鑒定植物抗寒性的重要指標之一[37]。本研究結果顯示,自然降溫后抗寒性較強的甘蔗品種桂糖28號、桂糖21號組織褐變最少,其次為福農15號、贛蔗18號的葉片,而抗寒性較弱的甘蔗品種如園林8號,園林6號葉片組織褐變嚴重。
葉片是植物進行光合和蒸騰作用的器官。正常植物葉肉細胞中的葉綠體膜上分布著葉綠素a和葉綠素b,所以葉片呈綠色。綠葉數越多,光合能力就越強。低溫環境下,葉綠體膜受破壞,葉綠體解體,葉綠素被分解,葉片變黃,不能行使其正常的功能。植物中綠葉數是鑒定植物抗寒性的重要指標之一[25]。本研究結果顯示,自然降溫后抗寒性最強的甘蔗品種桂糖28綠葉數最多,其次是桂糖21號、福農15號和贛蔗18號;而抗寒性較弱的甘蔗品種如ROC22、臺糖98/0432和園林8號、園林6號綠葉數較少,其中園林6號的綠葉數最少。
葉綠素含量是反映植物光合作用能力的一個間接指標,葉綠體是植物體內冷敏感性很強的細胞器,長期低溫會導致葉綠素的分解加快,葉綠素含量降低[38-41]。相關報道顯示,葉片葉綠素含量是鑒定作物抗寒性的指標之一[26,42]。在本研究中,自然降溫后期抗寒性較強的甘蔗品種桂糖28號、桂糖21的葉綠素含量較高;而抗寒性較弱的甘蔗品種如ROC22、臺糖98/0432和園林8號、園林6號含量較低。其中葉綠素含量最高的是桂糖28號,最低的是園林6號。
錘度是指可溶性固形物含量的比例,植物中可溶性固形物含量高能增強細胞滲透勢,降低細胞溶液的冰點,還可緩沖細胞質過度脫水,從而減少逆境脅迫對細胞的傷害[28]。糖還能清除羥自由基,抵抗逆境的傷害,可溶性糖在低溫脅迫下積累是各種植物的抗寒性研究中普遍采用的抗寒性指標[29-30]。本研究表明,甘蔗在自然降溫后期,錘度和糖分含量大部分都降低,但不同的品種降低的幅度不一樣,桂糖28號、桂糖21、福農15和贛蔗18糖分含量降低幅度明顯低于新臺糖22、臺糖98/0432和園林8號、園林6號,其中降幅最低是桂糖28號,降幅最高的是園林6號。
低溫對甘蔗的影響是復雜的,不僅表現在形態上,同時也表現在生理生化過程中。由于供試品種不同,蔗株對低溫的適應能力各異。因此,在進行甘蔗品種抗寒性鑒定時,不能使用單一的指標,而應以多個指標綜合評價。隸屬函數法已經成功應用于葡萄[16]、玉米[17]、小麥[43]、苜蓿[18]、月季[19]、甘薯[44]等其他作物的抗逆性評價,并證明其對抗性篩選的可靠性。本研究綜合前人的經驗,將隸屬函數分析法應用于自然低溫條件下的大田甘蔗抗寒性研究,應用多個形態和生理指標對不同甘蔗品種進行綜合抗寒性評價,提高了評價的準確性,在理論和實踐中都有一定的指導作用。
不同甘蔗品種對長時間持續低溫的反應各異,本研究根據7個相關性狀指標,運用隸屬函數法的分析方法對供試的8個甘蔗品種成熟期的抗寒性進行綜合評價,劃分為3級,其中Ⅰ級高抗寒型品種2個,Ⅱ級抗寒型品種2個,Ⅲ級低抗寒型品種4個,其中桂糖28號抗寒性最強,園林6號抗寒性最弱。
參考文獻:
[1]許文花,楊清輝. 甘蔗抗寒性研究進展[J]. 甘蔗,2003,10(3):8-12.
[2]譚宗琨,黃城華,孟翠麗,等. 甘蔗寒凍害等級指標及災損指標的初步研究[J]. 中國農學通報,2014,30(28):169-181.
[3]李茂枝. 淺談甘蔗抗寒性及防寒措施[J]. 中國糖料,1998(2):42-45.
[4]陳能武,楊榮仲,吳才文. 甘蔗品種抗寒性鑒定技術研究[J]. 甘蔗糖業,1996(4):1-9.
[5]張木清,陳如凱. 甘蔗抗寒性的研究——Ⅰ.低溫對甘蔗苗期生長的影響[J]. 福建省農科院學報,1992,7(1):14-22.
[6]孟翠麗,譚宗琨,李紫甜. 廣西甘蔗寒凍害空間反演模型研究[J]. 中國農業資源與區劃,2016,37(6):15-21,43.
[7]王鑒明. 中國甘蔗栽培學[M]. 北京:農業出版社,1985:292-296.
[8]覃蔚謙. 從建國后數次凍害看今冬凍害甘蔗糖的損失[J]. 廣西蔗糖,2000(1):14-18.
[9]梁廣焜. 甘蔗霜凍害及其防抗措施[J]. 甘蔗糖業,1994(6):7-8.
[10]馬 麗,王麗萍,蔡 青,等. 自然霜凍后甘蔗種質耐霜性鑒定[J]. 亞熱帶農業研究,2005,1(4):1-4.
[11]Singh O,Ralham N. A review of low temperatures stress resistance in sugarcane[J]. Indian Sugar,1998,48(6):421-430.
[12]柯仿鋼. 甘蔗霜害識別與預防[J]. 農藥信息市場,2012(7):51.endprint
[13]譚宗琨,歐釗榮,何 燕. 廣西蔗糖發展主要氣象災害分析及蔗糖產業優化布局的研究[J]. 甘蔗糖業,2006(1):17-21,33.
[14]王鑒明,保國裕,陳景形. 對甘蔗的新認識[J]. 甘蔗糖業,1980(8):3-7.
[15]張篤思. 甘蔗制糖工業分析[M]. 北京:中國輕工業出版社,1991:3-42.
[16]張文娥,王 飛,潘學軍. 應用隸屬函數法綜合評價葡萄種間抗寒性[J]. 果樹學報,2007,24(6):849-853.
[17]許桂芳,張朝陽,向佐湘. 利用隸屬函數法對4種珍珠菜屬植物的抗寒性綜合評價[J]. 西北林學院學報,2009,24(3):24-26.
[18]魏永勝,梁宗鎖,山 侖,等. 利用隸屬函數值法評價苜蓿抗旱性[J]. 草業科學,2005,22(6):33-36.
[19]許 鳳,張 顥. 楊春梅利用隸屬函數值法評價月季耐寒性[J]. 西南農業學報,2012,25(5):1870-1873.
[20]陳亞華,沈振國,劉友良. 低溫、高pH脅迫對水稻幼苗根系質膜、液泡膜ATP酶活性的影響[J]. 植物生理學報,2000,26(5):407-412.
[21]胡桂兵,陳大成,李 平,等. 荔枝果皮色素、酚類物質與酶活性的動態變化[J]. 果樹科學,2000,17(1):35-40.
[22]郁志芳,彭貴霞,夏志華,等. 鮮切山藥酶促褐變機理的研究[J]. 食品科學,2003,24(5):44-49.
[23]陳艷樂,申秀英,丁炳揚,等. 薯蕷多酚氧化酶特性及褐變控制[J]. 食品科學,2003,24(10):59-62.
[24]王 忠. 植物生理學[M]. 2版.北京:中國農業出版社,2009:124-125.
[25]焦雨歆,趙 琦,王雪英,等. 環境因子對植物葉綠體結構的影響[J]. 生物技術通報,2008(2):5-10.
[26]馬春平,宋麗萍,崔國文. 紫花苜蓿抗寒生理指標的此較研究[J]. 黑龍江畜牧獸醫,2006(6):47-48.
[27]李 進. 低溫脅迫下水稻幼苗抗寒生理指標分析[J]. 安徽農學通報,2014,20(12):21-22,68.
[28]利容千,王建波. 植物逆境細胞及生理學[M]. 武漢:武漢大學出版社,2002:160-164.
[29]羅煥亮,徐位力,李建忠,等. 馬占相思對低溫脅迫適應性的研究[J]. 華南農業大學學報(自然科學版),2002,23(2):51-53.
[30]劉麗杰,蒼 晶,王軍虹,等. 外源ABA對苗期低溫下冬小麥蔗糖含量及其關鍵酶基因表達的影響[J]. 麥類作物學報,2015,35(10):1333-1340.
[31]曹慧明,史作民,周曉波,等. 植物對低溫環境的響應及其抗寒性研究綜述[J]. 中國農業氣象,2010,31(2):310-314,319.
[32]Ren J,Huang Z L,Zeng L X,et al. A review of physiological reaction mechanism of plants exposed to low temperature stress[J]. World Forestry Research,2013,26(6):15-20.
[33]周慶鑫. 低溫下冬小麥活性氧清除物及膜脂組分對質膜穩定性影響的研究[D]. 哈爾濱:東北農業大學,2014:6-7.
[34]魏送送. 混濁梨汁酶促褐變機理及控制技術研究[D]. 西安:陜西科技大學,2014:55-56.
[35]Stewart G R,Larher F,Miflin B J. Accumulation of amino acids and related compounds in relation to environmental stress[J]. Biochem Plant,1980,5:609-635.
[36]薛楚然. 影響荔枝及荔枝酒酶促褐變的因素及控制研究[D]. 楊凌:西北農林科技大學,2015:34-35.
[37]姚 焱,劉向東,汪珍春,等. 低溫預處理對水稻花藥培養中花藥壁褐變的結構影響[J]. 熱帶亞熱帶植物學報,2006,14(4):307-311.
[38]王國莉,郭振飛. 水稻不同耐冷品種光呼吸對低溫的反應[J]. 作物學報,2005,31(5):673-676.
[39]陳 潔. 3種含笑屬植物抗寒性研究[D]. 長沙:中南林業科技大學,2015.
[40]安飛飛,簡純平,楊 龍,等. 木薯幼苗葉綠素含量及光合特性對鹽脅迫的響應[J]. 江蘇農業學報,2015,31(3):500-504.
[41]李春雷,倪德江. 氟對幼齡茶樹葉綠素含量及抗氧化酶活性的影響[J]. 江蘇農業學報,2015,31(5):1149-1153.
[42]陳 潔,金曉玲,寧 陽,等. 3種含笑屬植物抗寒生理指標的篩選及評價[J]. 河南農業科學,2016,45(2):113-118.
[43]陳榮敏,楊學舉,梁鳳山,等. 利用隸屬函數法綜合評價冬小麥的抗旱性[J]. 河北農業大學學報,2002,25(2):7-9.
[44]孫 凱,張勝利,朱弘博,等. 利用隸屬函數法對不同基因型甘薯耐鹽堿能力的分析與評價[J]. 東北師大學報(自然科學版),2015,47(2):115-119.endprint
猜你喜歡
中國綠色畫報(2016年11期)2017-02-18 15:21:32
中州學刊(2016年11期)2017-01-06 02:17:50
中國綠色畫報(2016年7期)2016-12-26 16:37:18
藝術科技(2016年10期)2016-12-14 20:02:46
文藝生活·下旬刊(2016年11期)2016-12-12 08:14:33
商情(2016年40期)2016-11-28 11:50:53
藝術科技(2016年9期)2016-11-18 18:35:07
人間(2016年27期)2016-11-11 15:56:02
中國科技博覽(2016年13期)2016-07-13 02:25:11
科技視界(2016年7期)2016-04-01 12:24:45
