NIH3T3細胞中miR-29b表達變化對mPer1和Egr2的影響*
2016-08-09 02:41:34徐濤后望汪宇輝成姝婷劉延友江舟肖靜郭慧玲王正榮
西部醫學 2016年7期
徐濤 后望 汪宇輝 成姝婷 劉延友 江舟 肖靜 郭慧玲 王正榮
(四川大學華西基礎醫學與法醫學院·衛生部時間生物學重點實驗室,四川 成都 610041)
?
·論著·
NIH3T3細胞中miR-29b表達變化對mPer1和Egr2的影響*
徐濤后望汪宇輝成姝婷劉延友江舟肖靜郭慧玲王正榮
(四川大學華西基礎醫學與法醫學院·衛生部時間生物學重點實驗室,四川 成都 610041)
【摘要】目的探討NIH3T3細胞中miR-29b對血清誘導的立早基因Egr2 mRNA表達的影響及其機制。方法將體外培養NIH3T3細胞分為實驗組、陰性對照組和空白對照組。實驗組用miR-29b轉染,陰性對照組空轉,而空白對照組則不做任何處理。RT-qPCR檢測近日節律基因mPer1與立早基因Egr2 mRNA的表達水平。結果實驗結果顯示,mPer1和Egr2的表達均發生了較明顯的變化。與陰性對照組相比,空白對照組中mPer1和 Egr2的mRNA表達水平無明顯差異;而實驗組中mPer1的mRNA表達降低了42%,Egr2的mRNA表達升高了49%。結論miR-29b能上調 Egr2的表達,其途徑可能是通過抑制NIH3T3細胞近日節律基因mPer1的表達實現。
【關鍵詞】mPer1;miR-29b;Egr2;NIH3T3細胞
近日節律系統是生物體的基本系統之一[1]。它能夠幫助生物體與每日外界環境的變化相協調,保證生物體的生理活動正常進行[2]。該系統由一系列基因及其表達產物相互作用形成的負反饋環路構成。其中Pers、Bmal1、Clock和Crys是其核心組成[3]。在維持生物節律穩定的同時,不同的核心節律基因在諸如細胞分化、生物發育等領域都扮演了重要角色[4]。
MicroRNAs(miRNAs)是一類廣泛存在于生物體內、長度在19~25核苷酸(nt)之間、高度保守的非編碼小RNA,是包括細胞增殖、分化、凋亡、發育、免疫調節等在內的眾多生物學進程的重要調節因子[5-6]。目前,越來越多的研究發現miRNA對近日鐘系統有非常重要的調節作用。例如,miR192/194和miR-29b基因簇可以負性調節per1、per2和per3的表達[6-7]。miR-142-3p 能夠直接作用于Bmal1的3′UTR區域抑制Bmal1的表達[8]。這些發現提示我們,miRNA可能是近日鐘調節的重要組成部分。
早期生長應答基因2(early growth response 2,Egr2)是20世紀80年代在果蠅胚胎發育分節現象的研究中發現的一種轉錄因子[9]。它包括一個C2H2型的鋅指結構,在中腦的發育中起著非常重要的作用[10]。研究顯示,Egr2受節律基因Per2和Dbp的調控且具有節律性[11]。而我們的前期實驗提示miR-29b可能調節節律基因per1的表達。這種在調節區域和參與的生物學事件的高度重合提示了miR-29b可能通過影響Egr2表達水平,從而對上述生理活動發揮調節作用。
1材料與方法
1.1PCR引物合成PCR引物由英濰捷基(上海)貿易有限公司和廣州銳博生物科技有限公司合成。引物序列,見表1。
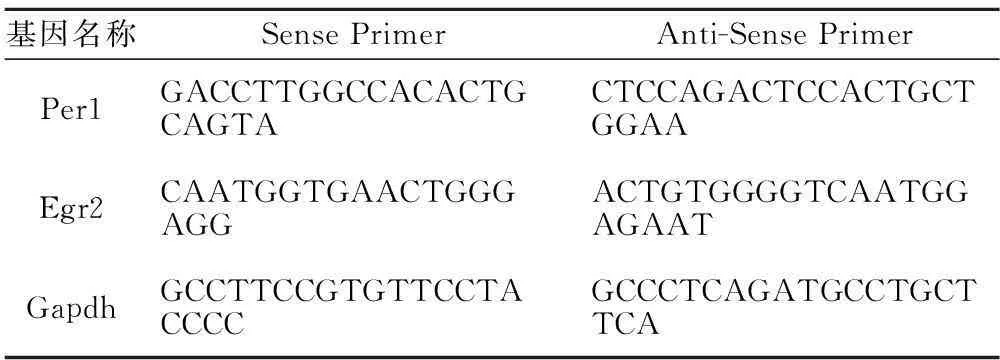
表1 PCR引物序列
1.2細胞培養細胞培養條件:選取NIH3T3細胞,用DMEM高糖培養基加入10%新生牛血清作為完全培養基,5% CO2和37℃條件下進行培養。在對數生長期用胰酶消化后以5×105/孔接種到六孔板中,待生長到對數生長期時進行細胞轉染。細胞轉染:選取對數生長期的NIH3T3細胞,用Opti-MEM無血清培養基清洗兩次;加入1.5ml的Opti-MEM無血清培養基待用。對照組:直接加入0.5ml的Opti-MEM無血清培養基;陰性對照組:配制0.5ml的只含lipo 2000的轉染液,孵育20min后加入待用的六孔板中;實驗組:配制0.5ml的含有miR-29b和lipo 2000的轉染液,孵育20min后加入待用的六孔板中。孵育5~6小時后更換含有血清的培養基,48小時后提取總RNA。
1.3實時熒光定量PCR根據Eastep?Super總RNA提取試劑盒說明書提取NIH3T3細胞的總RNA,用微量分光光度計NanoDrop 2000檢測RNA的濃度和純度。分別取1μg的RNA用逆轉錄試劑盒反轉錄合成cDNA。合成的cDNA稀釋成5倍的量待用。RT-qPCR反應體系:9μl的SYBR Green Mix,1μl上游引物,1μl下游引物,2μl cDNA模板,補加高壓滅菌蒸餾水至總體積20μl。反應條件,預變性:95℃,10min,擴增條件:95℃,10s;55~65℃,20s;72℃,20s;40個循環;融解曲線:65~95℃,5s。其中Gapdh、mPer1和Egr2各自的退火溫度分別為56.9℃、62.5℃和62℃。而miRNA的逆轉錄反應需要用特定的RT引物,反轉錄合成的cDNA不需要稀釋。RT-qPCR反應體系及反應條件同上。以U6作為檢測的內參基因。
1.4統計學分析mPer1與Egr2基因的mRNA相對表達水平以2-△△Ct方法計算。數據用SPSS 13.0軟件分析,采用單因素方差分析法,當P<0.05為差異有統計學意義。
2結果
2.1miR-29b轉染NIH3T3細胞前后細胞狀態的變化轉染前細胞的狀態正常(見圖1);轉染期的細胞空白對照組的細胞狀態正常(見圖2A),陰性對照組(見圖2B)和實驗轉染組(見圖2C)的細胞都出現了明顯的死亡,觸角變短。
2.2miR-29b顯著抑制了NIH3T3細胞內mPer1基因的表達以U6為內參基因(銳博公司提供),采用RT-qPCR法檢測空白對照組、陰性對照組和實驗轉染組中miR-29b mRNA的相對表達。實驗組(29b):脂質體轉染試劑加miR-29b轉染NIH3T3 細胞;陰性對照組(NC):脂質體轉染試劑加陰性對照miRNA轉染NIH3T3 細胞;空白對照組(Blank):正常生長的NIH3T3細胞。結果顯示,相對于空白對照組和陰性對照組,實驗組miR-29b的相對表達明顯升高(P<0.001),而兩個對照組則沒有顯著差異(見圖3)。其中空白對照組miR-29b的相對表達量為0.22,陰性對照組miR-29b的相對表達量為0.11,實驗組miR-29b的相對表達量為1.04。
圖1轉染前NIH3T3細胞的狀態
Figure 1State NIH3T3 cells before transfection
注:A.為空白對照組;B.為陰性對照組;C.為實驗轉染組

圖2轉染后NIH3T3細胞的狀態
Figure 2State NIH3T3 cells after transfection
注:A.為空白對照組;B.為陰性對照組;C.為實驗轉染組
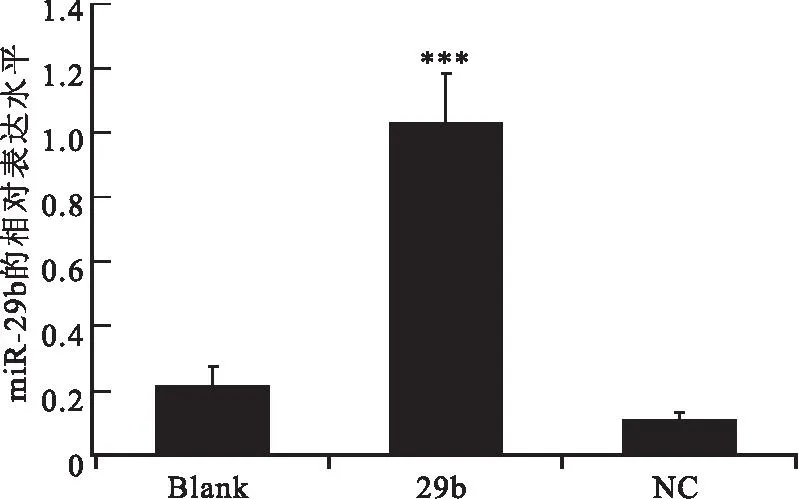
圖3miR-29b在NIH3T3細胞中的相對表達
Figure 3The relative expression of miR-29b in NIH3T3 cells
注:miR-29b的表達明顯升高(P<0.001),其中Blank為空白對照組,NC為陰性對照組,29b為實驗組
以Gapdh為內參基因,RT-qPCR證實相對于空白對照組和陰性對照組,實驗組mPer1的相對表達明顯降低(P<0.05),而空白對照組和陰性對照組之間沒有差異(見圖4)。其中空白對照組mPer1的相對表達量為1.07,陰性對照組mPer1的相對表達量為1.05,實驗組mPer1的相對表達量為0.63。
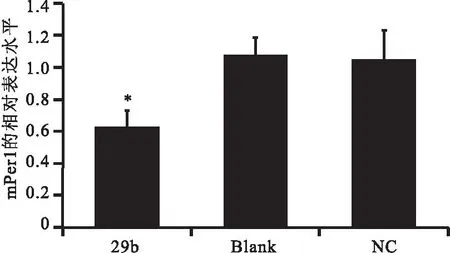
圖4mPer1在NIH3T3細胞中的相對表達
Figure 4The relative expression of mPer1 in NIH3T3 cells
注:mPer1的表達明顯降低(P<0.05),其中Blank為空白對照組,NC為陰性對照組,29b為實驗組
2.3miR-29b上調Egr2基因表達以Gapdh為內參基因,采用RT-qPCR法檢測陰性對照組、空白對照組和實驗組Egr2基因的mRNA的相對表達水平。實驗組(29b):脂質體轉染試劑加miR-29b轉染NIH3T3 細胞;陰性對照組(NC):脂質體轉染試劑加陰性對照miRNA轉染NIH3T3 細胞;空白對照組(Blank):正常生長的NIH3T3細胞。結果顯示,相對于空白對照組和陰性對照組,實驗組Egr2的相對表達明顯升高(P<0.05),而兩個對照組之間則不存在顯著性差異(見圖5)。其中空白對照組Egr2的相對表達量為0.80,陰性對照組Egr2的相對表達量為0.96,實驗組Egr2的相對表達量為1.45。

圖5Egr2在NIH3T3細胞中的相對表達
Figure 5The relative expression of Egr2 in NIH3T3 cells
注:Egr2的表達明顯升高(P<0.05),其中Blank為空白對照組,NC為陰性對照組,29b為實驗組
3討論
miRNA是一類非編碼的小RNA分子,能在轉錄后水平上改變基因的表達[12]。最近的研究表明,miRNA能夠調控近日節律系統。miR-219能夠調控近日周期的長短和速度,敲除體內miR-219能夠延長晝夜周期[13]。miR-132能夠調節近日鐘基因的表達和光誘導重置近日節律的過程[14]。這些研究結果表明,miRNA可以作為近日節律系統的一項重要的效應器。miR-29s家族可以結合Per1的3′UTR區域來抑制Per1的表達[15]。同時已有報道證實mPer1也在動物代謝調節和生物發育及細胞分化方面扮演了重要角色[16]。
Egr2的表達受多個信號通路如Wnt、FGF等信號通路的調控[17],進而對動物脂肪細胞的分化及神經系統的發育產生影響[18]。有研究報道,Egr2表達呈現出節律性,并且與近日節律基因Per2和Dbp的變化趨勢類似[11]且由于Egr2在中腦的發育過程中至關重要,而生物節律的中央調控位于下丘腦的SCN區域,加之二者所參與的生物學事件高度重合,由此可以推測,Per1和 Egr2之間可能存在一定的關系。但是Per1是通過何種機制來刺激Egr2的表達卻沒有研究。而近日節律基因的紊亂與某些細胞增殖與凋亡的發生息息相關,Per1作為核心的近日鐘基因能夠參與到該過程之中[4]。實驗結果表明,miR-29b轉染NIH3T3細胞影響了Per1和Egr2的表達,具體表現為Per1的表達降低,而Egr2的表達升高。這提示miR-29b能上調Egr2的表達,而這種改變有可能是通過Per1來介導的。但這還需要實驗進一步證實。
4結論
本研究證實,通過miR-29b抑制NIH3T3細胞內mPer1基因表達同時顯著上調Egr2的表達。該結果提示了miR-29b可通過影響Egr2基因表達的方式在諸多生理活動中發揮調控作用,這將有助于相關疾病的檢測、預防與治療,為病理性疾病的預防和治療提供了新的思路和理論基礎。
【參考文獻】
[1] Oike H,Oishi K,Kobori M.Nutrients,Clock Genes,and Chrononutrition[J].Current Nutrition Reports,2014,3(3):204-212.
[2]Mohammed S,Harrison D A.The clocks that time us are not the same:A theory of temporal diversity,task characteristics,and performance in teams [J].Organ Behav Hum Dec,2013,122(2):244-256.
[3]Albrecht,Urs.Timing to Perfection:The Biology of Central and Peripheral Circadian Clocks[J].Neuron,2012,74(2):246-260.
[4]Bae K,Jin X,Maywood ES,et al.Differential Functions of mPer1,mPer2,and mPer3 in the SCN Circadian Clock[J].Neuron,2001,30(2):525-536.
[5]Bartel DP.MicroRNAs:genomics,biogenesis,mechanism,and function[J].Cell,2004,116(2):281-297.
[6]Zhao Xiyan,Zhu Xueqiang,Cheng Shuting,et al.MiR-29a/b/c regulate human circadian gene hPER1 expression by targeting its 3′UTR[J].Acta Biochim Biophys Sin,2014,46 (4):313-317.
[7]Nagel R,Clijsters L,Agami R.The miRNA-192/194 cluster regulates the Period gene family and the circadian clock [J].The FEBS journal,2009,276(19):5447-5455.
[8]Shende V R,Neuendorff N,Earnest D J.Role of miR-142-3p in the post-transcriptional regulation of the clock gene Bmal1 in the mouse SCN [J].PloS one,2013,8(6):e65300.
[9]Weger N,Schlake T.Igf-I signalling controls the hair growth cycle and the differentiation of hair shafts [J].Invest Dermatol,2005,125(5):873-882.
[10] Shampa D,Jack E,Turman,Jr.Krox-20 gene expression:Influencing hindbrain-carniofacial developmental interaactions[J].Arch Histol Cytol,2005,68(4):227-234.
[11] Alvarez JD,Dechun Chen,Elizabeth Storer,et al.Non-cyclic and Developmental Stage- Specific Expression of Circadian Clock Proteins During Murine Spermatogenesis[J].Biology of Reproduction,2003,69(1):81-91.
[12] Cheng H-YM,Papp JW,Varlamova O,et al.microRNA modulation of circadian clock period and entrainment[J].Neuron,2007,54(5):813-829.
[13] Kocerha J,Faghihi M A,Lopez-toledano M A,et al.MicroRNA-219 modulates NMDA receptor-mediated neurobehavioral dysfunction [J].Proceedings of the National Academy of Sciences of the United States of America,2009,106(9):3507-3512.
[14] Alvarez-saavedra M,Antoun G,Yanagiya A,et al.miRNA-132 orchestrates chromatin remodeling and translational control of the circadian clock [J].Human molecular genetics,2011,20(4):731-751.
[15] Nicola Amodio,Marco Rossi,Lavinia Raimondi,et al.miR-29s:a family of epi-miRNAs with therapeutic implications in hematologic malignancies[J].Oncotarget,2015,6(15):12837-12861.
[16] Ansari N,Agathagelidis M,Lee C,et al.Differential maturation of circadian rhythms in clock gene proteins in the suprachiasmatic nucleus and the pars tuberalis during mouse ontogeny[J].The European journal of neuroscience,2009,29(3):477-489.
[17] Gabet Y,Baniwal SK ,Leclerc N,et al.Krox20/EGR2 deficiency accelerates cell growth and differentiation in the monocytic lineage and decreases bone mass[J].Blood,2010,116(19):3964-3971.
[18] Zhu C,Javier I,Bruce M,et al.Krox20 stimulates adipogenesis via C/EBPβ-dependent and -independent mechanisms[J].Cell Metab,2005,1(2):93-106.
基金項目:國家自然科學基金(31371180);國家自然科學青年基金(3150035)
通訊作者:汪宇輝,Email:wangyuhui1972@163.com
【中圖分類號】R 392;R 393
【文獻標志碼】A
doi:10.3969/j.issn.1672-3511.2016.07.003
(收稿日期:2016-01-19;編輯:母存培)
The effects of miR-29b expression change on mPer1 and Egr2 in NIH3T3 cells
XU Tao,HOU Wang,WANG Yuhui,et al
(Key Laboratory of Chronobiology,Ministry of Health (Sichuan University) ,West China School of Preclinical and Forensic Medicine,Sichuan University,Chengdu 610041,China)
【Abstract】ObjectiveTo investigate the effects of miR-29b on serum-induced immediate early gene Egr2 mRNA expression in NIH3T3 cells.MethodsNIH3T3 cells cultured in vitro were divided into experiment group,negative control group and blank control group.Experimental group transfected with miR-29b,negative control group idling,while the control group received no treatment.The expression of mPer1 and Egr2 were detected by RT-qPCR.ResultsThe experimental results show that mPer1 and Egr2 expressions were more obvious changes.Compared with the negative control group,the mRNA expression level of mPer1 and Egr2 were not obvious difference in the blank control group; but,the mRNA expression of mPer1 was reduced 42%,the mRNA expression of Egr2 was increased 49% in the experimental group.ConclusionmiR-29b can increase Egr2 expression,the approach is probably achieved by inhibiting circadian gene mPer1 expression in NIH3T3 cells.
【Key words】mPer1; mir-29b; Egr2; NIH3T3 cells
