在大腸桿菌內引入甲羥戊酸途徑高效合成抗瘧藥青蒿素前體——紫穗槐-4,11-二烯
2011-02-09 09:31:38吳濤吳勝明殷晴戴紅梅李樹龍董芳庭陳必鏈方宏清
生物工程學報 2011年7期
吳濤,吳勝明,殷晴,戴紅梅,李樹龍,董芳庭,陳必鏈,方宏清
1 福建師范大學 生命科學學院,福州 350108
2 軍事醫學科學院 生物工程研究所,北京 100071
3 軍事醫學科學院 國家生物醫學分析中心,北京 100850
青蒿素 (Artemisinin),一種倍半萜內酯過氧化物,是我國科學工作者從植物黃花蒿 Artemisia annua L.中分離并鑒定的抗瘧有效單體[1]。青蒿素與一種或多種其他抗瘧藥物配伍后得到一種抗瘧特效藥——以青蒿素為基礎的聯合藥物療法(Artemisinin-based combination therapies,ACTs)。2005年世界衛生組織 (WHO) 將ACTs推薦為治療非復雜惡性瘧疾的第一線用藥[2]。隨著 WHO的推薦,青蒿素的市場需求增大、價格迅速上漲,但青蒿素供應不足,極大地限制了ACTs的廣泛使用[3]。目前,從植物中提取仍是獲得青蒿素的主要途徑。大量種植黃花蒿能提高青蒿素的產量,但這是與糧食種植爭奪有限的土地資源,且易受氣候等外界因素的影響。化學方法合成青蒿素曾被寄予厚望,但青蒿素具有7個手性碳原子,存在128個異構體,得率低難以產業化[3]。利用微生物細胞合成青蒿素是替代植物提取和化學合成的方法之一,具有很好的應用前景。
2003年,Martin等[4]率先在大腸桿菌中引入人工合成的紫穗槐-4,11-二烯合酶基因 (ADS) 并重建釀酒酵母Saccharomyces cerevisiae H.來源的甲羥戊酸 (Mevalonate,MVA) 途徑,成功獲得了抗瘧藥青蒿素的前體——紫穗槐-4,11-二烯 (Amorpha-4,11-diene,AD)。之后,他們通過優化代謝途徑酶基因的密碼子、增加限速酶的表達水平、消除有毒代謝中間物或副產物的積累以及優化培養基和培養條件等[5-8],使紫穗槐-4,11-二烯產量極大提高,搖瓶培養產量為275 mg/L,上罐發酵產量達到27.4 g/L。 2006年,Lindahl等[9]采用構建質粒表達載體和同源重組基因組兩種方法,在釀酒酵母中引入黃花蒿來源的紫穗槐-4,11-二烯合酶基因,結果都獲得了紫穗槐-4,11-二烯,且前者產量達到600 μg/L,是后者的6倍。同年,Ro等[10]通過改造釀酒酵母內源的MVA途徑,并引入黃花蒿來源的紫穗槐-4,11-二烯合酶和細胞色素P450單加氧酶 (CYP71AV1) 基因,獲得了青蒿素的另一重要前體——青蒿酸 (Artemisinic acid),產量為100 mg/L。2008年,Ro等[11]以釀酒酵母為宿主,通過單個質粒表達紫穗槐-4,11-二烯合酶、紫穗槐-4,11-二烯氧化酶和細胞色素 P450還原酶等 3個植物來源酶的基因,使青蒿酸產量提高到1 g/L。這種微生物半合成青蒿素的方法——先通過改造微生物合成途徑獲得青蒿素的前體紫穗槐-4,11-二烯或青蒿酸等,再采用相對簡單的化學催化步驟可將這些前體轉化成青蒿素[12-13],可以大規模制備青蒿素,解決資源短缺問題。
大腸桿菌是應用最廣泛的原核表達系統,它具有操作簡單、周期短、成本低和產量高等優點。且大腸桿菌遺傳背景清楚,次生代謝產物簡單,目的代謝產物易于分離與純化。另外,大腸桿菌內存在以脫氧木酮糖磷酸 (Deoxyxylulose-5-phosphate,DXP) 途徑為基礎的類異戊二烯代謝途徑,能合成少量的輔酶Q等。因此,大腸桿菌非常適合作為生物合成類異戊二烯化合物的宿主菌。
本研究在大腸桿菌中引入人工合成的紫穗槐-4,1 1-二烯合酶基因并構建原核的糞腸球菌Enterococcus faecalis來源的MVA途徑,獲得了抗瘧藥青蒿素前體——紫穗槐-4,11-二烯的高表達。與文獻報道相比,本研究避開了眾所周知的真核生物來源的基因在原核細胞表達的各種難題。
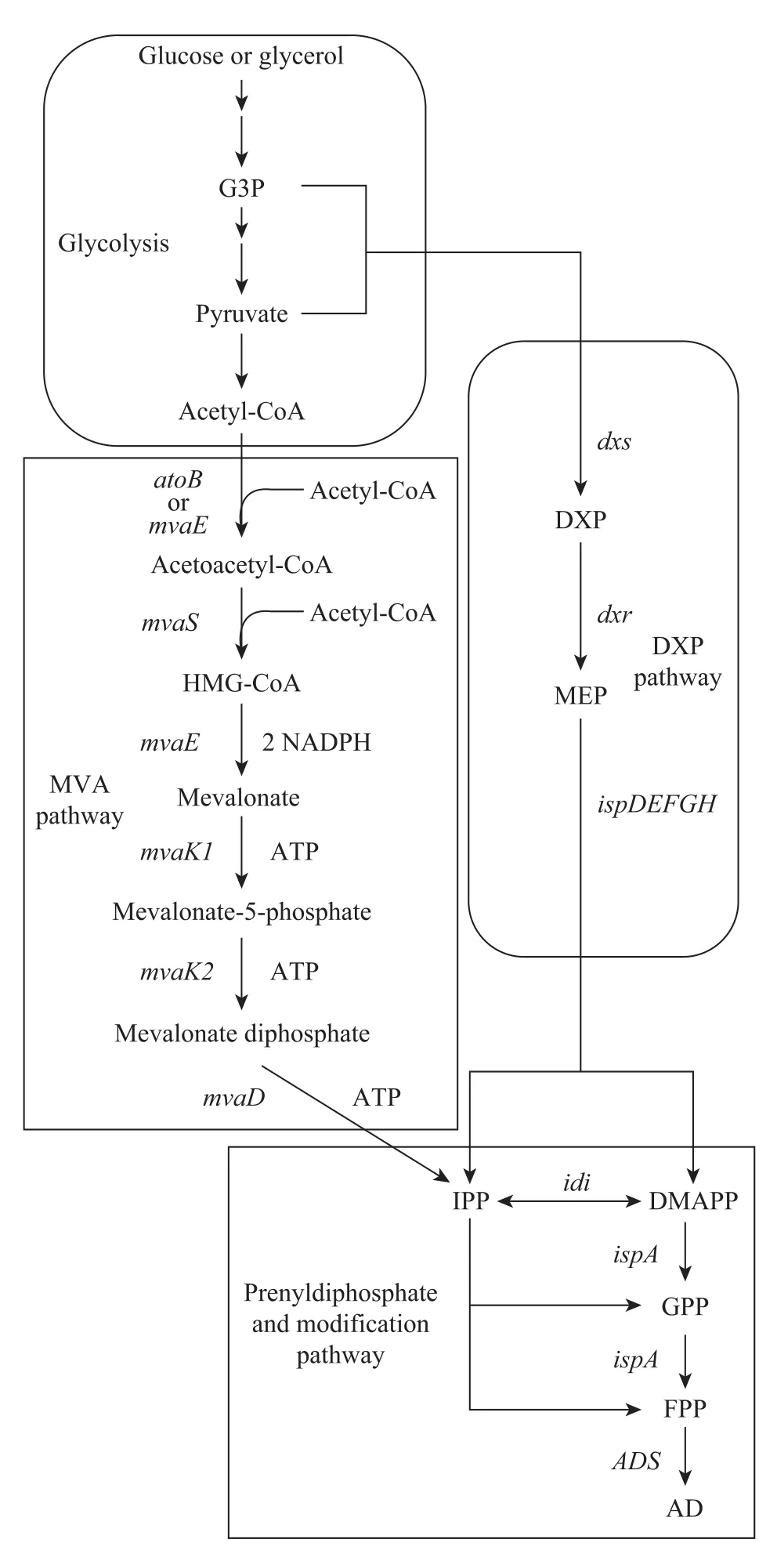
圖1 本實驗構建的重組菌的紫穗槐-4,11-二烯合成途徑Fig. 1 Synthesis of amorpha-4,11-diene in recombination E. coli from the building blocks of IPP and DMAPP, which are synthesised using DXP and MVA pathways. The native pathways are in round frames, and the heterologous pathways reconstructed in this paper are in square frames. The abbreviations are used: G3P, glyceraldehyde 3-phosphate; DXP, 1-deoxy-D-xylulose 5-phosphate; MEP, 2-C-methyl-D-erythritol 4-phosphate; HMG-CoA, 3-hydroxy-3-methyl-glutaryl-CoA; IPP, isopentenyl pyrophosphate; DMAPP, dimethyl allyl pyrophosphate; GPP, geranyl pyrophosphate; FPP, farnesyl pyrophosphate; AD, amorpha-4,11-diene.
1 材料與方法
1.1 實驗材料
1.1.1 菌株、質粒和培養基
宿主菌E. coli DHGT7為本室構建,表達載體pAOC3ANL為本實驗構建;糞腸球菌與質粒載體pET28a、pACYCDuet-1和pET3b由本室保存;菌株E. coli DH5α和BL21(DE3) 購于北京全式金生物技術公司。
LB培養基 (10 g/L胰蛋白胨,5 g/L酵母提取物,10 g/L NaCl) 用于大腸桿菌普通培養;FM2G培養基 (15 g/L胰蛋白胨,12 g/L酵母提取物,3 g/L NaH2PO4·2H2O,7 g/L K2HPO4·3H2O,2.5 g/L NaCl,0.5 g/L MgSO4,0.2%葡萄糖,1.5%甘油) 用于搖瓶發酵。
1.1.2 主要試劑
Prime STARTMHS DNA聚合酶、限制性內切酶和 T4 DNA連接酶購于寶生物工程公司;質粒提取試劑盒和DNA片段膠回收試劑盒購于北京博大泰恒生物技術公司;IPTG購于Promega公司;L-阿拉伯糖購于 Merck公司;標準品石竹烯((-)-trans-Caryophyllene) 購于Sigma-Aldrich公司。其他化學試劑均為國產或進口分析純。
1.1.3 相關基因的獲得
ADS根據文獻[14]提供的基因序列進行全基因合成。mvaE和 mvaS根據已公布的基因序列(GenBank Accession No. AF290092.1),MD (包含連續排列在基因組上 4個酶的基因:idi、mvaK2、mvaD 和 mvaK1) 和 ispA根據已公布的基因序列(GenBank Accession No. AE016830.1) 分別設計引物 (表1),以糞腸球菌的染色體 DNA為模板PCR擴增獲得。基因和引物由上海生工生物工程公司合成,基因測序由北京中科希林公司完成。
1.2 載體構建、目的基因表達
采用常用分子克隆技術,如PCR擴增、酶切、連接和轉化等構建質粒表達載體 (表 2),具體方法參考文獻[15]。
將表達載體轉化宿主菌E. coli DHGT7,接入含相應抗生素 (Kn、Cm均為25 μg/mL) 的液體LB培養基中,37 ℃培養至OD600=0.3~0.4,加入0.2% L-阿拉伯糖誘導,繼續培養 4~5 h。取菌體全蛋白作SDS-PADE分析,具體方法參考文獻[15]。

表1 本文使用的引物Table 1 Primers used in this study

表2 本文使用的菌株和質粒Table 2 Strains and plasmids used in this study
1.3 搖瓶培養方法
挑取新轉化的單克隆,接入含相應抗生素的液體 LB培養基中,37 ℃振蕩培養過夜。取過夜培養菌液轉接40 mL抗性FM2G培養基,使初始菌體濃度為OD600=0.05,37 ℃、220 r/min振蕩培養至OD600為0.3~0.4時,加入0.2% L-阿拉伯糖或0.5 mmol/L IPTG誘導,并加入20% (V/V) 無菌的十二烷覆蓋在培養基的表面[5],隨后轉至30 ℃繼續培養48~60 h。選取不同時間點測定菌體濃度和紫穗槐-4,11-二烯產量。
1.4 GC-MS法測定AD含量
1.4.1 儀器設備和樣品處理
氣相色譜-質譜聯用儀:HP6890氣相色譜和飛行時間質譜 (Waters公司);色譜柱:DB-5 (30 m× 250 μm×0.25 μm,Agilent公司)。
樣品處理方法:培養液離心分層,取10 μL十二烷層溶液加入到190 μL含10 μg/mL石竹烯為內標物的乙酸乙酯溶液中,渦旋混勻。
1.4.2 樣品檢測條件
氣相色譜條件:載氣:氦氣;流速:1.0 mL/min;程序升溫:70 ℃保持1 min,15 ℃/min升溫至285 ℃,保持2 min,30 ℃/min升溫至310 ℃;進樣口溫度:150 ℃;進樣方式:不分流進樣;進樣量:0.3 μL。
質譜條件:電離方式:電子轟擊電離 (EI);電離能量:70 eV;電子源溫度:220 ℃;掃描模式:全掃描。
2 結果與分析
2.1 表達ADS產紫穗槐-4,11-二烯
將全基因合成的 ADS克隆到質粒載體 pET28a上,得到表達載體 pET28-ADS;轉化 E. coli DHGT7,以pET28a為對照,進行SDS-PAGE分析,結果在約 56 kDa處有可見的紫穗槐-4,11-二烯合酶(ADS) 的條帶 (圖2)。
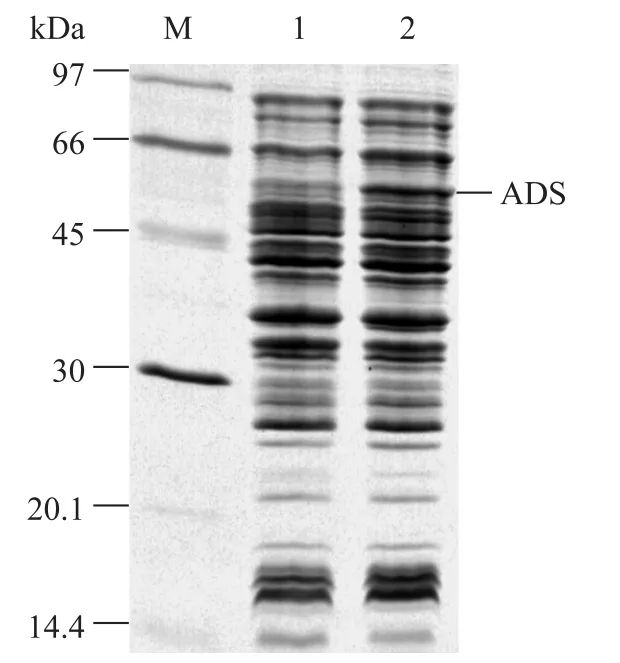
圖2 ADS誘導表達的菌體全蛋白SDS-PAGE分析結果Fig. 2 SDS-PAGE analysis showing the expressing of the synthetic ADS gene in DHGT7. M: protein marker; 1: pET28a/ DHGT7; 2: pET28-ADS/DHGT7.
新轉化的菌株pET28-ADS/DHGT7進行搖瓶培養,采用GC-MS法測定紫穗槐-4,11-二烯的含量。通過GC檢測,分別在8.42 min和8.83 min出現內標物石竹烯 (IS) 和紫穗槐-4,11-二烯 (AD) 的保留峰 (圖3A)。對樣品AD作MS檢測,質譜圖與紫穗槐-4,11-二烯的標準圖譜比對,二者一致 (圖3B、C)。根據內標石竹烯的濃度對樣品 AD含量進行定量,結果搖瓶培養48 h的紫穗槐-4,11-二烯產量僅為11.3 mg石竹烯當量/L。我們推測大腸桿菌內源的FPP水平太低,限制了紫穗槐-4,11-二烯的產量。由于大腸桿菌僅存在 DXP途徑,為避免對內源途徑的影響,后面引入了異源的MVA途徑來提高前體供給。
2.2 構建異源MVA途徑
以低拷貝質粒 pACYCDuet-1的骨架為基礎,PCR擴增質粒pET3b的多克隆位點 (MCS),并通過PCR引物引入NotⅠ和ApaⅠ限制性酶切位點,將PCR擴增的MCS序列連接到pACYCDuet-1的SphⅠ和 EcoRⅠ之間,構建了一個具有特殊 MCS的質粒載體 pAOC3ANL,用于多基因合成途徑的構建。
將糞腸球菌來源的 mvaE、mvaS、MD和 ispA與ADS分別構建在質粒載體pAOC3ANL和pET28a上,得到表達載體 pAOC3-ES-MD和 pET28-ispAADS,共轉化E. coli DHGT7,搖瓶培養48 h紫穗槐-4,11-二烯產量達到 151 mg/L,是利用內源 FPP的13.3倍。
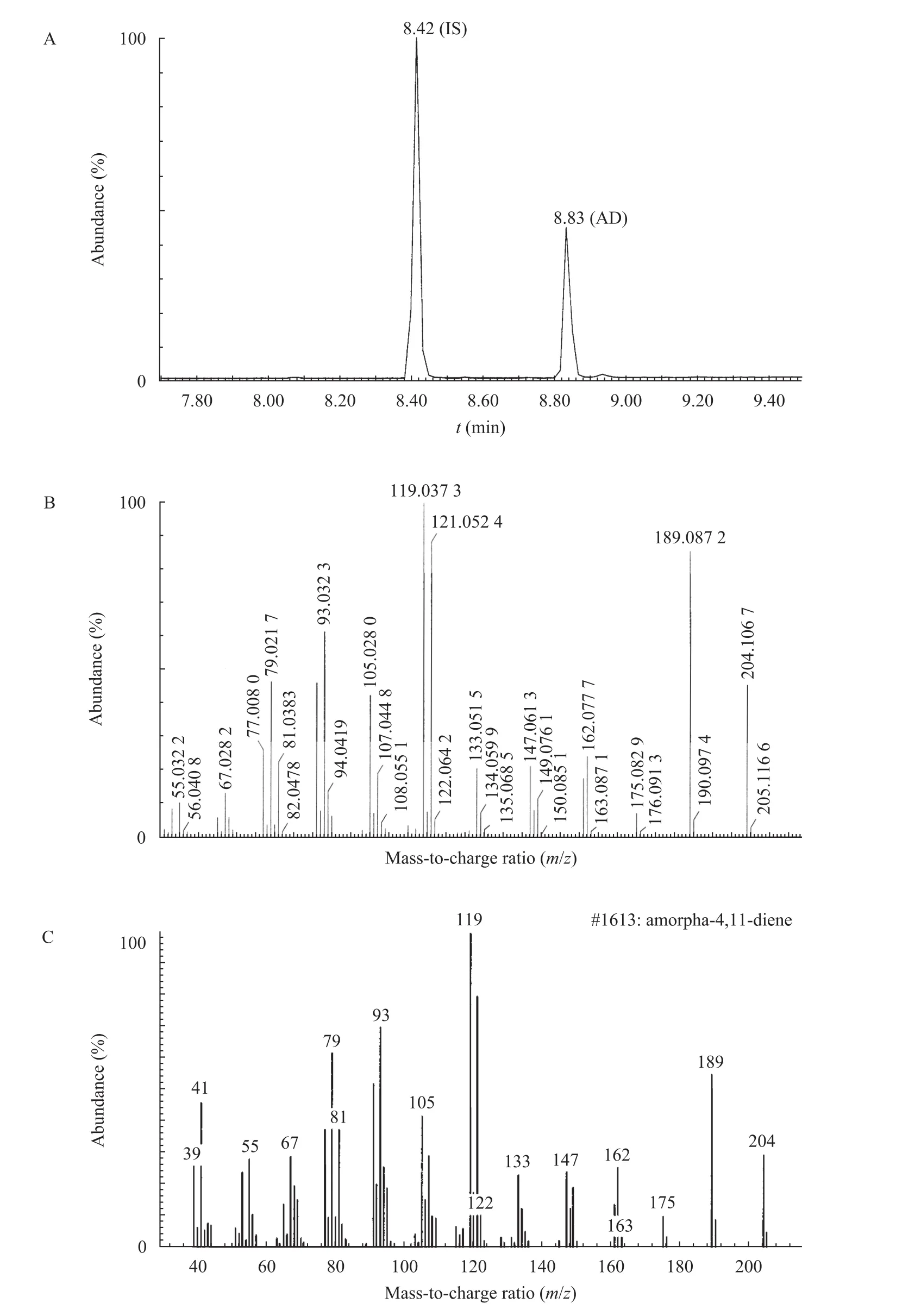
圖3 紫穗槐-4,11-二烯的GC-MS測定結果Fig. 3 GC-MS analysis of amorpha-4,11-diene produced by pET28-ADS/DHGT7. (A) Total ion chromatogram of pET28-ADS/ DHGT7. (B) Mass spectrum of the peak of retention time at 8.83 min in A. (C) Mass spectrum of amorpha-4,11-diene[16].
2.3 優化紫穗槐-4,11-二烯合成途徑
為進一步提高紫穗槐-4,11-二烯的產量,必須消除代謝途徑的限制因素,如提高限制酶的水平、消除毒性中間物或副產物等,因此我們作了如下優化。
將整個紫穗槐-4,11-二烯合成途徑的基因構建到單個質粒載體 pAOC3ANL上,得到表達載體pAOC3-ES-MD-ispA-ADS (圖 4),轉化 E. coli DHGT7,紫穗槐-4,11-二烯產量為32.5 mg/L,僅為使用 2個質粒共表達時的 1/5,菌體的生長也受到嚴重抑制 (圖5A)。我們推斷這是因為當使用2個質粒共表達時ADS是構建在高拷貝的pET28a上,而以單個質粒表達時 ADS是構建在低拷貝的pAOC3ANL上,間接地降低了ADS的拷貝數,使得胞內FPP累積,引起內源FPP合成途徑的反饋調控作用,從而抑制菌體生長。
因此,我們提高了ADS的拷貝數,將pET28-ADS與 pAOC3-ES-MD-ispA-ADS 共轉化 E. coli DHGT7,結果紫穗槐-4,11-二烯產量提高了4.3倍,達到140 mg/L (圖5B),菌體生長抑制也得到一定程度的緩解。
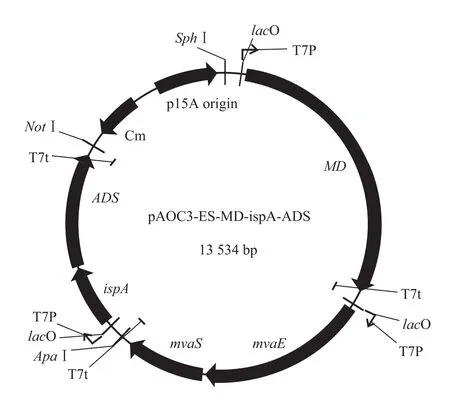
圖4 表達載體pAOC3-ES-MD-ispA-ADS的示意圖Fig. 4 Map of plasmid pAOC3-ES-MD-ispA-ADS. T7P and T7t represent the T7 promoter and T7 terminator respectively.
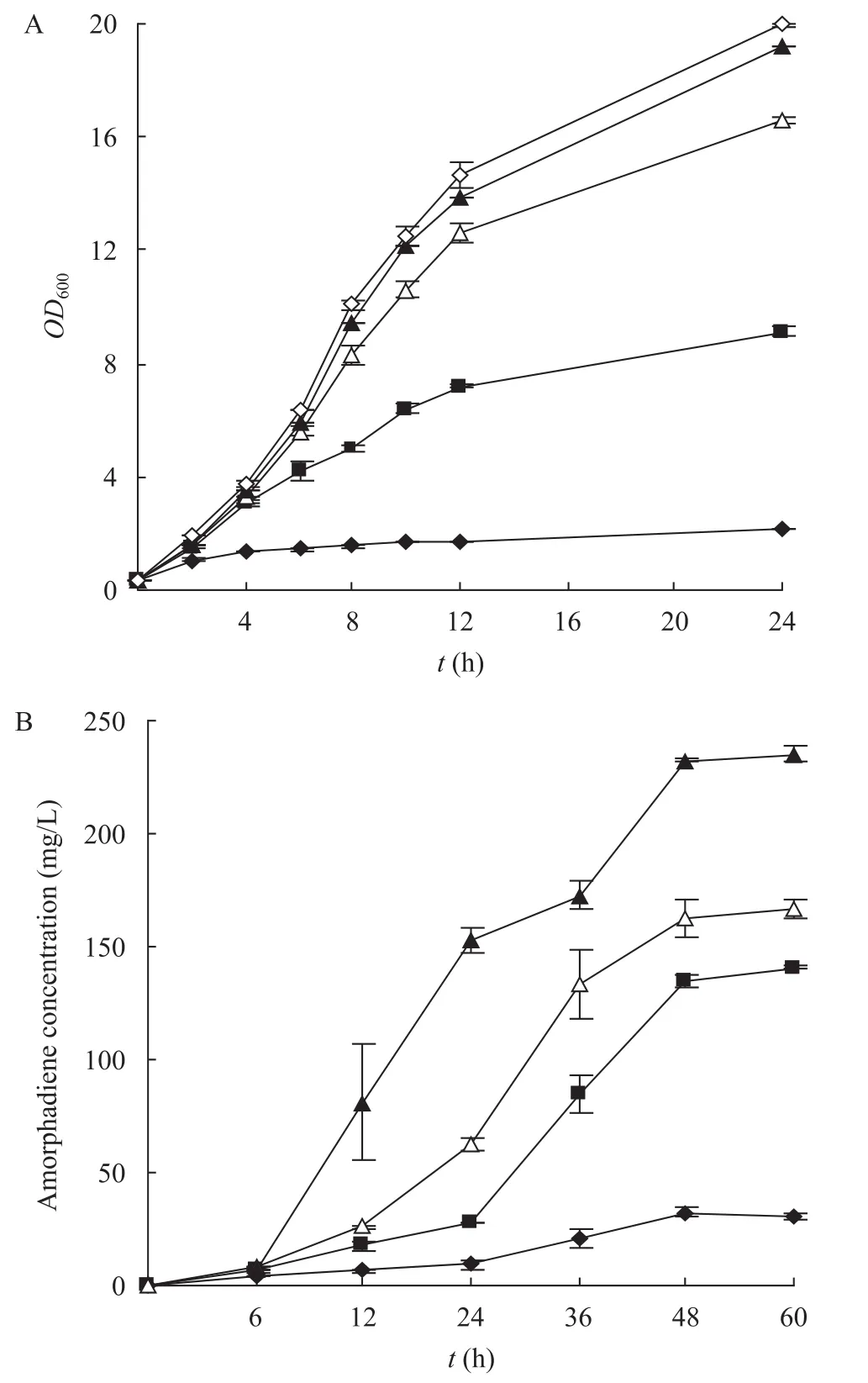
圖5 菌體生長曲線 (A) 和紫穗槐-4,11-二烯產量測定曲線 (B)Fig. 5 Growth curves and amorpha-4,11-diene production analysis of engineered strains. (A) Growth curves of E. coli DHGT7 showing the inhibition effect was eliminated by increasing another high copy of limiting enzymes gene. (B) Amorpha-4,11-diene production was measured from E. coli DHGT7 containing different expressing vectors. The symbols represent host cell E. coli DHGT7 containing different expressing vectors: pAOC3-ES-MD-ispA-ADS (?); pET28-ADS and pAOC3-ES-MD-ispA-ADS (■); pET28-E-ADS and pAOC3-ES-MD-ispA-ADS (?); pET28-E-MK-ADS and pAOC3-ES-MD-ispA-ADS (▲); and pAOC3ANL (?).
據Pitera等[6]報道,HMG-CoA還原酶是類異戊二烯化合物生物合成過程中的關鍵酶,它催化HMG-CoA轉化為甲羥戊酸的反應是不可逆的,HMG-CoA還原酶活性不足會導致HMG-CoA在胞內累積產生毒性。因此,我們提高HMG-CoA還原酶水平,構建了表達載體pET28-E-ADS,與pAOC3-ES-MD-ispA-ADS共轉化E. coli DHGT7,結果菌體生長抑制得到進一步緩解,紫穗槐-4,11-二烯產量也再次提高。
另據 Anthony等[7]報道,提高胞內甲羥戊酸激酶 (MK) 的水平可以極大地提高紫穗槐-4,11-二烯產量。因此我們構建了表達載體pET28-E-MK-ADS,與pAOC3-ES-MD-ispA-ADS共轉化E. coli DHGT7,結果紫穗槐-4,11-二烯產量達到235 mg/L,是優化前的7.2倍,菌體生長抑制基本消除。
2.4 不同宿主合成紫穗槐-4,11-二烯
為考察優化過的紫穗槐-4,11-二烯合成途徑對不同宿主的影響,我們以菌株E. coli BL21(DE3) 為宿主,共轉化表達載體 pET28-E-MK-ADS和pAOC3-ES-MD-ispA-ADS,搖瓶培養60 h的紫穗槐-4,11-二烯產量為201 mg/L,菌體生長正常。說明我們構建青蒿素前體的合成途徑對不同的大腸桿菌具有通用性。
3 討論
本研究以基因組改造過的菌株 E. coli DHGT7為宿主,構建質粒載體引入異源的類異戊二烯合成途徑,成功合成了目的產物青蒿素前體——紫穗槐-4,11-二烯,產量達到235 mg/L,是Kong等[17]報道的國內最高產量的近2倍,稍低于2009年Tsuruta等[8]報道的最高產量275 mg/L。同時,我們以E. coli BL21(DE3) 為宿主,也獲得了紫穗槐-4,11-二烯。說明我們構建的青蒿素前體的合成途徑對不同的大腸桿菌具有通用性。
大腸桿菌能通過內源的DXP途徑合成FPP,但水平較低。我們在大腸桿菌中引入人工合成的紫穗槐-4,11-二烯合酶基因,內源的FPP不足限制了紫穗槐-4,11-二烯產量,因此提高胞內FPP水平是提高紫穗槐-4,11-二烯產量的關鍵。為了避免對內源 DXP途徑的干擾,我們引入了糞腸球菌來源的 MVA途徑,以提高胞內FPP的水平,使紫穗槐-4,11-二烯產量提高了13.3倍,達到151 mg/L。
在類異戊二烯合成途徑中,中間代謝產物如HMG-CoA、IPP、DMAPP和FPP等積累,會抑制菌體的生長[6]。Anthony等[7]證明甲羥戊酸激酶是合成途徑的限速酶,提高胞內甲羥戊酸激酶的水平可以極大地提高紫穗槐-4,11-二烯產量;而當甲羥戊酸激酶不足時,會導致甲羥戊酸的積累,但甲羥戊酸幾乎不影響菌體生長。我們通過提高紫穗槐-4,11-二烯合酶、HMG-CoA還原酶和甲羥戊酸激酶等3個酶的水平,使紫穗槐-4,11-二烯產量提高了7.2倍。
值得指出的是,在大腸桿菌中引入異源的MVA途徑合成類異戊二烯需要消耗大量的乙酰輔酶 A,但這并不會對菌體生長產生不利影響,也不會因為乙酰輔酶A不足降低產物產量。Tabata等[18]在大腸桿菌中構建甲羥戊酸合成途徑,以內源的乙酰輔酶A為底物合成甲羥戊酸的產量達到47 g/L。Choi等[19]和 Kang等[20]也分別利用大腸桿菌內源的乙酰輔酶 A合成 PHB達到 89.8 g/L和細胞干重的85.8%。
另外,我們以低拷貝的質粒載體pACYCDuet-1為基礎,構建了具有特殊多克隆位點的表達載體pAOC3ANL。采用單個低拷貝質粒載體表達整個合成途徑的基因,用另一相容的高拷貝表達載體pET28a來增加限速酶基因的拷貝數,協調合成途徑的表達,避免了多質粒載體的不相容性影響。
我們采用FM2G培養基搖瓶培養,在其表面覆蓋 20%的十二烷,這是因為目的產物紫穗槐-4,11-二烯在水相中易揮發,超過97%的紫穗槐-4,11-二烯可被覆蓋在培養基表面的十二烷有機層“捕獲”[5],而十二烷本身不會被菌體代謝,且對菌體的生長無副作用[21]。Newman等[5]利用這種含十二烷的兩相分配封閉反應器獲得紫穗槐-4,11-二烯產量達到0.5 g/L。
本研究構建的合成紫穗槐-4,11-二烯的工程菌以阿拉伯糖為誘導物,避免了文獻中大量采用的IPTG誘導的缺點。IPTG是有毒且有異味的物質,不適宜在大規模工業化生產中應用。而阿拉伯糖則不存在上述缺點,而且來源豐富,在規模化生產上將具有明顯的優勢。
本研究利用大腸桿菌為宿主,構建異源紫穗槐-4,11-二烯從頭合成途徑,并對合成途徑進行優化,通過提高紫穗槐-4,11-二烯合酶、HMG-COA還原酶和甲羥戊酸激酶等3個酶的水平,消除了抑制菌體生長的不利因素,使紫穗槐-4,11-二烯產量達到235 mg/L。繼續對培養基和培養條件等進行優化,或通過上罐發酵,將獲得更高的紫穗槐-4,11-二烯產量。這為高效生物合成抗瘧藥青蒿素前體——紫穗槐-4,11-二烯奠定了基礎。
REFERENCES
[1] Klayman DL. Qinghaosu (artemisinin): an antimalarial drug from China. Science, 1985, 228(4703): 1049?1054.
[2] Olumese P. Guidelines for the treatment of malaria. Geneva: World Health Organization Press, 2006: 1?2.
[3] White NJ. Qinghaosu (artemisinin): the price of success. Science, 2008, 320(5874): 330?334.
[4] Martin VJJ, Pitera DJ, Withers ST, et al. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat Biotechnol, 2003, 21(7): 796?802.
[5] Newman JD, Marshall J, Chang M, et al. High-level production of amorpha-4,11-diene in a two-phase partitioning bioreactor of metabolically engineered Escherichia coli. Biotechnol Bioeng, 2006, 95(4): 684?691.
[6] Pitera DJ, Paddon CJ, Newman JD, et al. Balancing a heterologous mevalonate pathway for improved isoprenoid production in Escherichia coli. Metab Eng, 2007, 9(2): 193?207.
[7] Anthony JR, Anthony LC, Nowroozi F, et al. Optimization of the mevalonate-based isoprenoid biosynthetic pathway in Escherichia coli for production of the anti-malarial drug precursor amorpha-4,11-diene. Metab Eng, 2009, 11(1): 13?19.
[8] Tsuruta H, Paddon CJ, Eng D, et al. High-Level production of amorpha-4,11-diene, a precursor of the antimalarial agent artemisinin, in Escherichia coli. PLoS ONE, 2009, 4(2): e4489.
[9] Lindahl AL, Olsson ME, Mercke P, et al. Production of the artemisinin precursor amorpha-4,11-diene by engineered Saccharomyces cerevisiae. Biotechnol Lett, 2006, 28(8): 571?580.
[10] Ro DK, Paradise EM, Ouellet M, et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature, 2006, 440(7086): 940?943.
[11] Ro DK, Ouellet M, Paradise EM, et al. Induction of multiple pleiotropic drug resistance genes in yeast engineered to produce an increased level of anti-malarial drug precursor, artemisinic acid. BMC Biotechnol, 2008, 8: 83.
[12] Acton N, Roth RJ. On the conversion of dihydroartemisinic acid into artemisinin. J Org Chem, 1992, 57(13): 3610?3614.
[13] Haynes RK, Vonwiller SC. Cyclic peroxyacetal lactone, lactol and ether compounds: US, 5420299. 1995-05-30.
[14] Wallaart TE, Bouwmeester HJ. DNA encoding amorpha-4,11-diene synthase: EP, 0982404. 2000-03-01.
[15] Sambrook J, Russell DW. Molecular cloning: a laboratory manual. 3rd ed. New York: Cold Spring Harbor Laboratory Press, 2001: 1232?1235.
[16] Bouwmeester HJ, Wallaart TE, Janssen MH, et al. Amorpha-4,11-diene synthase catalyses the first probable step in artemisinin biosynthesis. Phytochemistry, 1999, 52(5): 843?854.
[17] Kong JQ, Wang W, Wang LN, et al. The improvement of amorpha-4,11-diene production by a yeast-conform variant. J Appl Microbiol, 2009, 106(3): 941?951.
[18] Tabata K, Hashimoto S. Production of mevalonate by a metabolically-engineered Escherichia coli. Biotechnol Lett, 2004, 26(19): 1487?1491.
[19] Choi JI, Lee SY. High level production of supra molecular weight poly(3-hydroxybutyrate) by metabolically engineered Escherichia coli. Biotechnol Bioprocess Eng, 2004, 9(3): 196?200.
[20] Kang Z, Wang Q, Zhang HJ, et al. Construction of a stress-induced system in Escherichia coli for efficient polyhydroxyalkanoates production. Appl Microbiol Biotechnol, 2008, 79(2): 203?208.
[21] Daugulis AJ, Janikowski TB. Scale-up performance of a partitioning bioreactor for the degradation of polyaromatic hydrocarbons by Sphingomonas aromaticivorans. Biotechnol Lett, 2002, 24(8): 591?594.
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中國果業信息(2019年10期)2019-11-13 01:21:34
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
聚氯乙烯(2018年9期)2018-02-18 01:11:34
中國衛生(2016年3期)2016-11-12 13:23:26
