二氧化氯對褐腐菌、酵母菌及大腸埃希菌的抑菌作用研究
2024-01-01 04:31:18幸紅霞姚倩張雨鋮廖俊米董娟余沁郭曉強
中國抗生素雜志 2023年8期
關鍵詞:酵母菌
幸紅霞 姚倩 張雨鋮 廖俊米 董娟 余沁 郭曉強
摘要:目的 考察二氧化氯對桃褐腐病主要致病菌褐腐菌(Monilinia fructicola)、酵母菌(Kazachstania exigua)以及大腸埃希菌(Escherichia coli)的抑制效果。方法 以褐腐菌、酵母菌、大腸埃希菌為實驗對象,采用固體培養法測定菌斑直徑和細菌存活率;測定胞外培養液電導率,蛋白質和DNA含量,檢測胞內丙二醛濃度和脫氫酶活性,并采用掃描電鏡進行形態學觀察。結果 隨著二氧化氯濃度升高,菌斑直徑和細菌存活率逐漸降低,胞外培養液電導率、蛋白質、DNA含量和胞內丙二醛含量增加,脫氫酶活性顯著下降;二氧化氯對褐腐菌的抑菌效果顯著高于酵母菌和大腸埃希菌(P<0.05)。電鏡掃描下可見細菌細胞膜邊界模糊,有明顯滲出液。結論 二氧化氯能有效抑制褐腐菌的生長,其機制與破壞細菌細胞膜,使細菌代謝酶失活有關。
關鍵詞:二氧化氯;褐腐菌;酵母菌;大腸埃希菌;抑菌機制
中圖分類號:R978.1 ?文獻標志碼:A
Antibacterial effects of chlorine dioxide on Monilinia fructicola, yeast and Escherichia coli
Xing Hongxia1, Yao Qian2, Zhang Yucheng2, Liao Junmi3, Dong Juan3, Yu Qin2, and Guo Xiaoqiang2
(1 School of Food and Biological Engineering, Chengdu University, Chengdu 610106; 2 School of Pharmacy, Chengdu University, Chengdu 610106; 3 Sichuan Shuangjinduotai Biotechnology Co., LTD, Chengdu 610100)
Abstract Objective To study the inhibitory effect of ClO2 on Monilinia fructicola , the primary pathogen of peaches, as well as yeast (Kazachstania exigua) and Escherichia coli (E. coli). Methods The plaque diameter and bacterial survival rate of Monilinia fructicola, yeast, and E. coli were measured using the solid culture method. The conductivity, protein, and DNA content of the extracellular culture medium, as well as the intracellular malondialdehyde content and dehydrogenase activity, were all tested using the same approach. Finally, scanning electron microscopy was applied to observe the bacterial morphology. Results The plaque diameter and bacterial survival rate gradually reduced as ClO2 concentration increased. However, extracellular conductivity, protein and DNA content as well as intracellular malondialdehyde content increased, while dehydrogenase activity declined dramatically. The inhibitory effect of ClO2 on Monilinia fructicola was notably stronger than that on yeast and E coli (P<0.05). The blurred boundary of bacterial cell membrane with visible intracellular effusion were observed under SEM. Conclusion The antibacterial mechanisms are associated with the impairment of cell membrane and inactivation of bacterial metabolic enzymes.
Key words Chlorine dioxide; Monilinia fructicola; Yeast; E. coli; Antibacterial mechanism
二氧化氯是一種新型的空氣消毒劑,具有適用面廣、使用劑量低、滅菌率高、持續時間長、無毒副作用及對環境無污染等優點,己被世界衛生組織列為A1級安全高效的綠色消毒劑[1],被公認為食品除臭和消毒最突出和最有效的化學品之一。二氧化氯對飲用水、水果和蔬菜進行消毒處理時通常采用水溶液或者氣體形式[2];研究表明,二氧化氯氣體處理后殘留的氯氣比其水溶液形式少,且表面的滲透速度也更快[3]。成都龍泉山脈區域是全國重要的水蜜桃生產基地之一,有“天下第一桃”的美譽[4]。桃的軟組織和薄果皮在采后貯藏期間易受到真菌感染引起疾病,褐腐菌是桃果實采后軟腐病的主要致病菌[5],由褐腐菌引起的褐腐病是桃收獲后最具破壞性的病害[6],在貯藏、運輸、銷售及消費過程中極易發生,使果實迅速腐敗,導致無法食用,造成巨大的經濟損失[7]。大腸埃希菌是食品主要污染源之一,食用后易引起腹瀉、嘔吐等消化道癥狀。酵母菌可利用果皮表面和傷口處的營養迅速大量繁殖,從而消耗傷口處營養且布滿整個傷口表面[8]。細菌加速了桃的腐爛變質,不利于貯藏和運輸。目前,桃果實采后主要依賴于化學殺菌劑防治微生物污染,但殺菌劑的過度使用會造成環境污染、對人體健康有一定的危害,為減少環境污染和滿足消費者對安全食品日益增長的需求,尋找新的褐腐菌防治方法迫在眉睫[9]。本文選擇水蜜桃貯存和運輸過程中常見的3種菌-褐腐菌、酵母菌和大腸埃希菌,采用固體反接培養方法考察二氧化氯對3種菌的抑制作用,以期更好地模擬果蔬貯藏的實際狀況,找到高效低毒的水蜜桃保鮮策略,為延長水蜜桃貨架期、提高水蜜桃營養價值提供有益指導。
1 材料與儀器
1.1 實驗菌株
大腸埃希菌BNCC336902(Escherichia coli)、酵母菌JC2636(Kazachstania exigua)均由成都大學四川省生物醫藥基礎實驗教學中心微生物實驗室提供。
1.2 試劑
丙二醛(malondialdehyde,MDA,AR,純度≥98%,廣東翁江化學試劑有限公司);營養瓊脂和LB肉湯購自青島海博生物技術有限公司;硫代巴比妥酸、乙酸異戊酯和2, 3, 5-三苯基氯化四氮唑(2, 3, 5-triphenyl tetrazolium chloride,TTC)購自上海泰坦科技股份有限責任公司;二氧化氯(AR,含量10%)由四川金科藥業有限公司提供。
1.3 儀器
UV5100B紫外可見分光光度計(上海元析儀器有限公司),FE30臺式電導儀(梅特勒-托力多儀器有限公司),SPX-250生化培養箱(常州邁科諾儀器有限公司),CT14D高速冷凍離心機(上海天美生化儀器設備工程有限公司),Quanta 200掃描電子顯微鏡(scanning electron microscope, SEM,美國FEI儀器有限公司)。
2 方法
2.1 褐腐菌的分離與復蘇
2.1.1 褐腐菌的分離
在龍泉水蜜桃病果上挑取少量腐爛部分,用無菌水連續稀釋,0.1 mL稀釋液置于PDA滅菌固體培養基,涂布均勻,25 ℃培養1 d,根據菌落形態挑選單個菌落,轉移至新的滅菌培養基中培養,重復3次。經形態學和PCR驗證[10],鑒定為褐腐菌。菌株置于PDA斜面上,4 ℃低溫保存。
2.1.2 菌種復蘇與生長
用接種環挑取褐腐菌、酵母菌、大腸埃希菌,分別接種于100 mL LB滅菌液體培養基中,分別在
25 ℃、28 ℃和37 ℃下以100 r/min振蕩培養24 h。調整其濃度為106 CFU/mL,吸取50 μL菌懸液置于PDA平板中央,涂布均勻,將培養皿分別置于25 ℃、
28 ℃和37 ℃下培養1 d備用[11]。
2.2 反接培養法
參照文獻[11]的方法并稍作改進。將滅菌的PDA培養基倒入培養皿中,冷卻備用,無菌打孔器(6 mm)取“2.1.2”項下含菌培養基,制得菌餅,將其反接于PDA平板中央;精確稱量二氧化氯固體,分別用蒸餾水配置成0.0125、0.025、0.05和0.1 mmol/L的二氧化氯溶液10 mL,將10 mL二氧化氯溶液倒入50 mL燒杯中;將燒杯和培養皿(加蓋)放置于10 L塑料盒中,將塑料盒分別置于25 ℃、28 ℃和37 ℃下培養16 h。取出培養皿,繼續培養直至空白組的菌落直徑大于培養皿直徑的2/3,結束培養。以未經二氧化氯處理的菌餅作為空白組。
2.3 抑菌活性測定
2.3.1 菌落直徑
培養結束后,用游標卡尺采用十字交叉法測定各組菌斑直徑3次,結果取平均值。
2.3.2 存活率
菌落培養結束后,用小刀刮下相同大小的菌斑(直徑約4 cm),置于試管中,用20 mL無菌PBS分散均勻,于600 nm波長處測吸光值,與空白組的吸光度值做比較,按下式計算存活率lg(Nt/N0)[12]:
lg(Nt/N0)=lg(At/A0)
其中,Nt和N0分別表示二氧化氯處理組和空白組的細菌數量,At和A0分別表示二氧化氯處理組和空白組細菌混懸液的吸光度值。
2.4 抑菌機制考察
2.4.1 電導率
取“2.3.2”項下培養結束后的菌斑,用無菌水分散樣品置于燒杯中,用電導儀測定電導值記為A1,測完后將所有樣品沸水浴15 min,冷卻后再次測其電導值記為A2,參照文獻[13]計算電導率。
電導率(%)=A1/A2×100
2.4.2 蛋白質和DNA
使用PBS沖洗反接處理培養后菌落,12000 r/min
離心30 min;吸取上清液,用紫外分光光度計測定A260和A280,DNA含量用260 nm波長處的吸光值表示,參照文獻[14]計算蛋白質含量。
蛋白質含量/(mg/mL)=1.45×A280 -0.74×A260
2.4.3 MDA測定
MDA含量測定參照文獻[15],并稍作改進。取反接菌斑,用PBS分散,隨后吸取1.5 mL,加入3 mL 10%三氯醋酸(w/v),在4 ℃、4500 r/min離心30 min;吸取上清液2 mL,加0.67% 硫代巴比妥酸(w/v)4.5 mL,沸水浴10 min,離心,于532 nm波長處測吸光值,根據MDA標準曲線計算含量。
2.4.4 脫氫酶活性測定
脫氫酶活性測定參照文獻[15],并稍作改進。取經PBS分散的菌液6 mL,加入2 mL Tris-HCl(pH 8.8)溶液,以及2 mL TTC-葡萄糖溶液,37 ℃水浴加熱2 h,分別加入7.5 mL甲苯和冰醋酸溶液,室溫放置1 h后,于485 nm波長處測吸光值;以未經二氧化氯處理的細菌作為陰性對照。
2.5 細菌形態觀察
取“2.1.2”項下培養的細菌,二氧化氯處理16 h,用6 mm無菌打孔器取菌餅,分散于滅菌蒸餾水中,8000 r/min離心5 min;棄上清液,加入2 mL 2.5%戊二醛(w/v),4℃下放置12 h;然后參照Tao等[16]方法進行梯度洗脫,樣品經自然干燥后,噴金、上樣,在掃描電鏡下觀察形態。
2.6 統計學分析
樣品測定重復3次,以平均值±標準偏差表示。使用Origin 2021軟件作圖,SPSS 26.0軟件進行統計分析[17],當P≤0.05時,組間有顯著性差異。
3 結果
3.1 抑菌活性
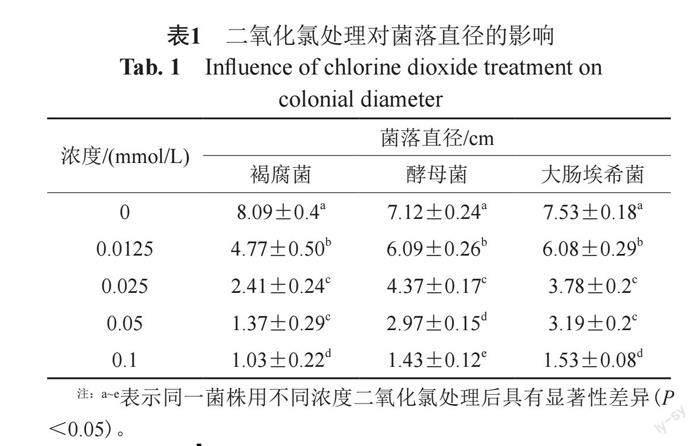
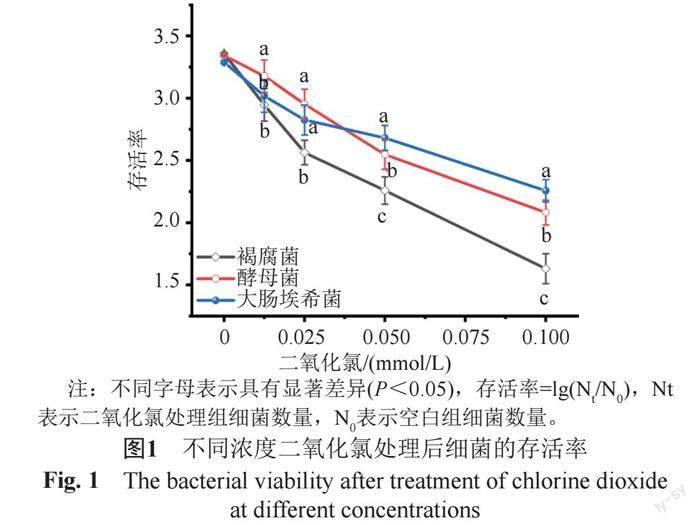
菌落直徑結果如表1所示,二氧化氯濃度與菌落直徑呈負相關,即濃度越大,菌落直徑越小。采用相同濃度的二氧化氯處理后,褐腐菌的菌落直徑均顯著小于酵母菌和大腸埃希菌,表明二氧化氯對褐腐菌的殺菌效果最好。細菌存活率表現了相同的趨勢,即隨著二氧化氯濃度的升高,細菌存活率下降,顯示了二氧化氯較強的殺菌作用(圖1);濃度≥0.025 mmol/L時褐腐菌的存活率顯著低于酵母菌和大腸埃希菌(P<0.05),濃度≥0.05 mmol/L時酵母菌存活率顯著低于大腸埃希菌(P<0.05)。
3.2 抑菌機制研究
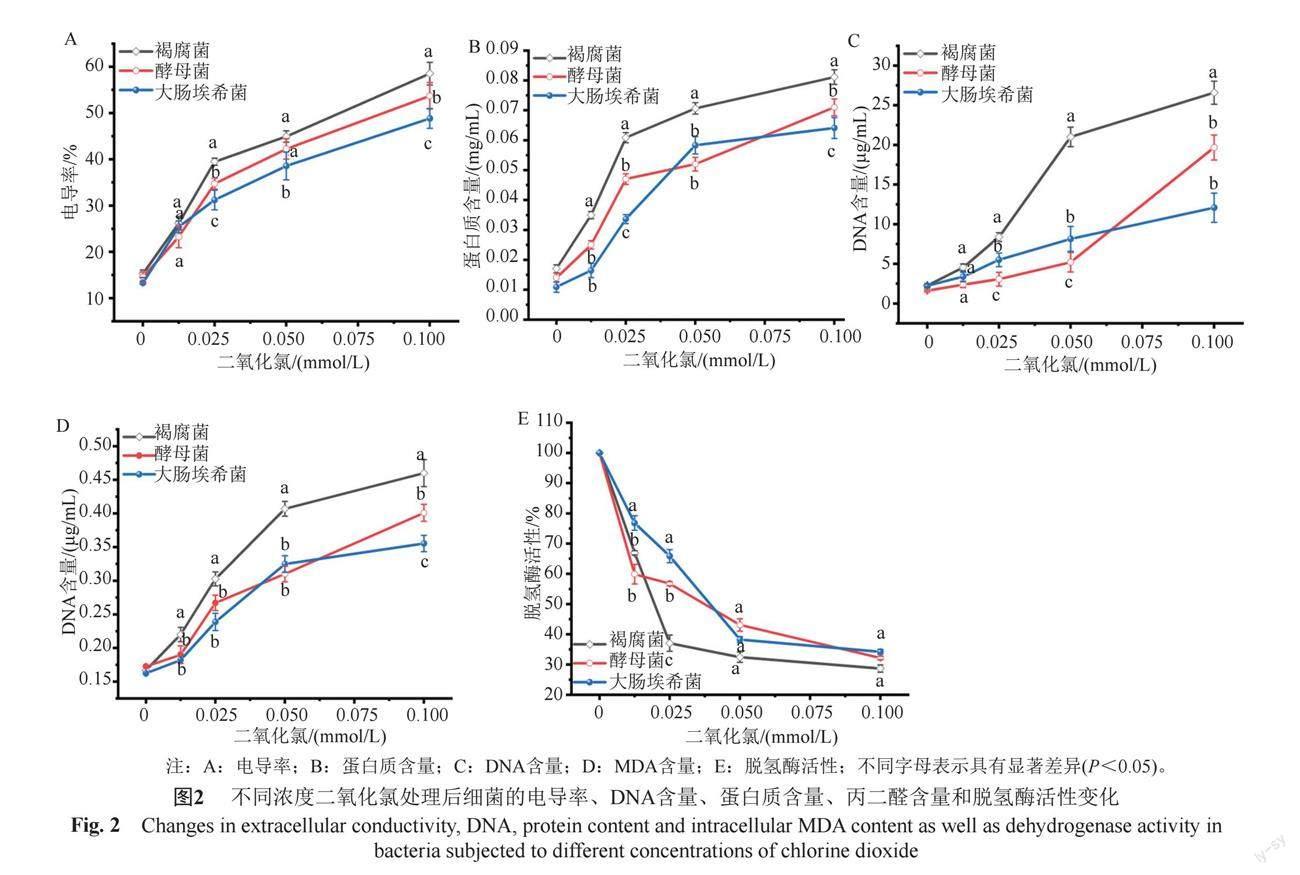
3.2.1 電導率
電導率測定結果如圖2(A)所示,隨著二氧化氯濃度的增加,細菌培養液的電導率不斷增加;在二氧化氯濃度≤0.0125 mmol/L時,3種菌株電導率無顯著性差異;二氧化氯濃度升至0.025 mmol/L,電導率順序為褐腐菌>酵母菌>大腸埃希菌。
3.2.2 蛋白質含量
培養基中細菌泄露的蛋白質含量測定結果如圖2(B)所示,二氧化氯處理后,褐腐菌、酵母菌和大腸埃希菌滲出的蛋白質含量與二氧化氯濃度呈正相關,在多數濃度下,蛋白質含量順序為褐腐菌>酵母菌>大腸埃希菌(P<0.05),褐腐菌胞外蛋白的含量均顯著高于酵母菌和大腸埃希菌。
3.2.3 DNA含量
細胞外DNA含量測定結果如圖2(C)所示,隨著二氧化氯濃度的增加,細菌胞外DNA含量逐漸增大,當二氧化氯濃度至0.025 mmol/L時,褐腐菌DNA含量急劇增大;二氧化氯濃度達0.05 mmol/L時,酵母菌胞外DNA開始迅速上升,而大腸埃希菌的DNA呈現勻速增大趨勢。二氧化氯濃度為0.05 mmol/L時,3種細菌DNA含量順序為褐腐菌>大腸埃希菌>酵母菌(P<0.05);當氣體濃度增至0.1 mmol/L時,DNA含量為褐腐菌>酵母菌>大腸埃希菌,褐腐菌胞外DNA濃度顯著高于其他兩種細菌(P<0.05)。
3.2.4 MDA含量
細菌MDA含量測定結果如圖2(D)所示,隨著二氧化氯濃度增加,細菌胞內MDA含量逐漸增大;在多數濃度作用下,MDA含量順序為褐腐菌>酵母菌>大腸埃希菌,濃度≥0.0125 mmol/L時褐腐菌胞內產生的MDA顯著高于酵母菌和大腸埃希菌(P<0.05)。
3.2.5 ? ?脫氫酶活性
脫氫酶活性測定結果如圖2(E),隨著二氧化氯濃度增加,3種細菌的脫氫酶活性迅速降低。二氧化氯濃度為0.0125 mmol/L時,大腸埃希菌的酶活性顯著高于褐腐菌與酵母菌(P<0.05),表明二氧化氯對大腸埃希菌酶的破壞作用相對較弱;二氧化氯濃度增至0.025 mmol/L時,3種菌株酶活性為大腸埃希菌>酵母菌>褐腐菌(P<0.05);二氧化氯濃度≥
0.05 mmol/L時,3種菌株的脫氫酶活性持續降低,組間均無顯著性差異(P>0.05),表明高濃度二氧化氯可使不同細菌的脫氫酶失活,從而引起細菌的死亡。
3.3 ? ?細菌形態變化
SEM掃描結果如圖3所示,空白組形態完整,表面較為光滑,基本沒有褶皺,SEM背景較干凈,說明無內容物流出。處理組中3種菌的部分形態發生改變,表面干癟粗糙、凹陷,細胞膜受損,邊界變得模糊,鏡下可見多處團聚體和滲出液,其中褐腐菌變形最嚴重,形態嚴重塌陷并有大量滲出液,這與滅活效果一致。
4 討論
大量研究表明,二氧化氯可抑制細菌、病毒、孢子形成的生物體,具有來源廣泛、安全性高、毒副作用小和環境友好等優勢。褐腐菌引起的褐腐病是桃果實采后的重要侵染性病害,二氧化氯對褐腐菌的抑菌研究較少。本實驗結果顯示,與酵母菌和大腸埃希菌相比,二氧化氯能較好抑制褐腐菌生長,原因可能是褐腐菌有菌絲體,二氧化氯能較快進入褐腐菌的菌絲細胞,抑制菌落生長。Yu等[18]報道二氧化氯氣體可抑制黃曲霉的菌絲體生長、孢子萌發和AFB1產毒能力。
二氧化氯促進微生物的死亡機制被認為有以下兩種,一種是由于細胞表面蛋白質巰基的氧化和細胞膜通透性的增加[19],另一種是通過影響細胞內部成分,如蛋白質和核酸含量,并干擾蛋白質的合成[20]。細菌懸浮液中的電導率可以反映細胞膜的滲透性改變,如果電解質滲漏,表明細胞膜被破壞,可檢測到胞外培養液電導率升高[21-22]。二氧化氯處理后3種菌株的電導率增加,說明二氧化氯對細菌細胞膜有一定程度的破壞作用,引起細胞膜內電解質外滲。二氧化氯處理后3種菌株的胞外蛋白質和DNA含量均有所增加,表明二氧化氯處理后細菌的細胞膜遭到破壞,使胞內物質泄露到細胞外。蛋白質和DNA是細菌維持生命活動的物質基礎,它與病原菌體內的正常生理活動和細胞周期等密切相關;細菌胞內蛋白質和DNA的滲出加速了細菌的死亡。MDA為細胞脂質膜的氧化產物,本實驗發現,經二氧化氯處理后,3種菌株的MDA含量均明顯增加,同時表現出了濃度依賴性,即隨著二氧化氯濃度升高,MDA含量增大,說明二氧化氯通過氧化脂質的形式使細菌細胞膜受損。同時,二氧化氯可抑制細菌相關代謝酶活性[20],TTC法測定脫氫酶活性可表征細菌呼吸活性的大小;本實驗顯示,低劑量的二氧化氯能使細菌脫氫酶活性急劇下降,在高劑量下脫氫酶活性降至40%以下,說明二氧化氯對細菌代謝酶有強烈的抑制作用,從而引起細菌的死亡。
通常,二氧化氯對真菌的抑制作用強于細菌。在本實驗中,當二氧化氯濃度高于0.025 mmol/L時,對酵母菌的抑制率顯著高于大腸埃希菌,但當濃度低于0.025 mmol/L時,二氧化氯對大腸埃希菌的抑制作用高于酵母菌(圖1),可能的原因為低濃度的二氧化氯即可破壞大腸埃希菌細胞核,使其DNA外溢;而酵母菌的細胞核在相同濃度作用下損傷較小,在培養液中僅檢出少量DNA,細胞核的完整性使酵母菌保持較強的活力(圖2C)。另外,不同真菌對藥物的敏感性存在差異,比如Pereira等[23]發現,氯氣對不同真菌的殺滅作用不一致,與培養真菌的基質有較大關系。本實驗所用培養基可能對酵母菌有一定保護作用,導致其對低濃度二氧化氯不敏感。
以上結果表明,二氧化氯對褐腐菌、酵母菌和大腸埃希菌均有抑制作用,其抑制機理可能是二氧化氯使細菌細胞膜氧化,導致細胞膜破損,胞內電解質、蛋白質和DNA滲出,引起細菌死亡;此外,二氧化氯對細菌代謝酶有強烈的抑制作用,研究結果為二氧化氯作為安全無毒抑菌劑的推廣應用提供了理論依據。二氧化氯的其他抑菌機制,如是否抑制蛋白的表達,是否改變細菌能量代謝和相關基因的表達等,有待進一步深入研究。
參 考 文 獻
康慧芳. 不同保鮮劑處理對葡萄交鏈孢霉腐病抑制效果的研究[D]. 上海: 上海師范大學, 2020.
Liu Z L, Cao Z Y, Wang J, et al. Chlorine dioxide fumigation: An effective technology with industrial application potential for lowering aflatoxin content in peanuts and peanut products[J]. Food Control, 2022, 136: 108847.
Kim S, Lee H, Ryu J H, et al. Inactivation of Bacillus cereus spores on red chili peppers using a combined treatment of aqueous chlorine dioxide and hot-air drying[J]. J Food Sci, 2017, 82(8): 1892-1897.
代婭. 水蜜桃氣調保鮮技術及不同處理保持果實硬度的機理研究[D]. 雅安: 四川農業大學, 2013.
李廣錦. 外源NO在中華壽桃應答褐腐菌脅迫中的作用及其蛋白組學研究[D]. 泰安: 山東農業大學, 2017.
Abate D, Pastore C, Gerin D, et al. Characterization of monilinia spp. populations on stone fruit in south Italy[J]. Plant Dis, 2018, 102(9): 1708-1717.
殷曉慧, 王慶國, 張暢, 等. 桃果實褐腐病拮抗菌的篩選、鑒定及其拮抗活性[J]. 食品工業科技, 2017, 38(9): 128-132.
趙魯寧. 水蜜桃炭疽病拮抗酵母菌的篩選、初步應用及其生防機理的研究[D]. 鎮江: 江蘇大學, 2018.
Tian S, Torres R, Ballester A R, et al. Molecular aspects in pathogen-fruit interactions: Virulence and resistance[J]. Postharvest Biol Tec, 2016, 122: 11-21.
陳淑寧. 桃褐腐病菌和炭疽病菌對DMI殺菌劑的抗性研究[D]. 武漢: 華中農業大學, 2017.
劉欣. 二氧化氯對指狀青霉致病力的影響及其機制研究[D]. 濟南: 齊魯工業大學, 2021.
Guan J W, Lacombe A, Tang J M, et al. Use of mathematic models to describe the microbial inactivation on baby carrots by gaseous chlorine dioxide[J]. Food Control, 2020, 123: 107832.
吳路芳. 影響致病疫霉孢子囊畸變的活性物的篩選與初步利用[D]. 保定: 河北大學, 2020.
Wen G, Xu X Q, Zhu H, et al. Inactivation of four genera of dominant fungal spores in groundwater using UV and UV/PMS: Efficiency and mechanisms[J]. Chem Eeg J, 2017, 328: 619-628.
Ersoya Z G, Dincb O, Cinar B, et al. Comparative evaluation of disinfection mechanism of sodium hypochlorite chlorine dioxide and electroactivated water on Enterococcus faecalis[J]. Lwt-Food Sci Technol, 2019, 102: 205-213.
Tao N G, Jia L, Zhou H E. Anti-fungal activity of Citrus reticulata Blanco essential oil against Peniillium italicum and Penicillium digitatum[J]. Food Chem, 2014, 153: 265-271.
Yao Q, Shen Y F, Bu L, et al. Ultrasound-assisted aqueous extraction of total flavonoids and hydroxytyrosol from olive leaves optimized by response surface methodology[J]. Prep Biochem Biotech, 2019, 49(9): 837-845.
Yu Y, Shi J Y, Xie B Y, et al. Detoxification of aflatoxin B1 in corn by chlorine dioxide gas[J]. Food Chem, 2020, 328: 127121.
Ulrike P, Herppich W B, Hassenberg K, et al. Aqueous chlorine dioxide treatment of horticultural produce: Effects on microbial safety and produce quality-A review[J]. Crit Rev Food Sci, 2018, 58(2): 318-333.
Bridges D F, Lacomeb A, Wu V C. Integrity of the Escherichia coli O157: H7 cell wall and membranes after chlorine dioxide treatment[J]. Front Microbiol, 2020, 11: 888.
Tang W W, Zeng X P, Zhao Y S, et al. Disinfection effect and its mechanism of electrolyzed oxidizing water on spores of Bacillus subtilis var. niger[J]. Food Sci Bioteclonol, 2011, 20(4): 889-895.
Zhu C H,Chen Z, Yu G Y. Fungicidal mechanism of chlorine dioxide on Saccharomyces cerevisiae[J]. Ann Microbiol, 2013, 63(2): 495-502.
Pereira V J, Marques R, Marques M, et al. Free chlorine inactivation of fungi in drinking water sources[J]. Water Res, 2013, 47(2): 517-523.
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
中國民間療法(2021年8期)2021-07-22 05:53:28
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
當代水產(2019年3期)2019-05-14 05:43:24
天然產物研究與開發(2018年9期)2018-10-08 03:25:32
煙草科技(2015年8期)2015-12-20 08:27:04
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
食品工業科技(2014年9期)2014-03-11 18:15:31
