內(nèi)皮祖細(xì)胞損傷來源微粒對內(nèi)皮祖細(xì)胞的影響機(jī)制研究*
2023-12-25 14:16:08馬藝萍阿不都熱合曼米吉提袁玉娟吐爾孫阿依依斯米提拉阿卜拉江艾合麥提穆葉賽尼加提
重慶醫(yī)學(xué) 2023年23期
關(guān)鍵詞:檢測
馬藝萍,阿不都熱合曼·米吉提,袁玉娟,吐爾孫阿依·依斯米提拉,阿卜拉江·艾合麥提,穆葉賽·尼加提
(1.新疆醫(yī)科大學(xué)研究生院,烏魯木齊 830000;2.喀什地區(qū)第二人民醫(yī)院心血管內(nèi)科,新疆喀什 844099;3.新疆維吾爾自治區(qū)人民醫(yī)院急救中心,烏魯木齊 830000)
心血管疾病(cardiovascular disease,CVD)是一種與內(nèi)皮細(xì)胞(endothelial cell,EC)水平相關(guān)的疾病[1]。EC損傷、血小板黏附和循環(huán)血液中的白細(xì)胞侵襲內(nèi)皮下層是動脈粥樣硬化形成的關(guān)鍵早期事件。內(nèi)皮祖細(xì)胞(endothelial progenitor cell,EPC)是一種多功能干細(xì)胞,具有向EC分化的潛能[2]。當(dāng)組織缺血或損傷時(shí),EPC從骨髓被動員到外周血中進(jìn)行分化。研究發(fā)現(xiàn),EPC能有效減輕心絞痛和心力衰竭,改善左心室射血分?jǐn)?shù),并顯示出組織器官再生和恢復(fù)缺血相關(guān)器官功能的前景[3]。微粒(microparticles,MPs)是由各種活化和凋亡細(xì)胞產(chǎn)生、直徑為100~1 000 nm的小膜囊泡[4-6]。MPs是細(xì)胞間物質(zhì)交換和信號轉(zhuǎn)導(dǎo)的重要介質(zhì),攜帶表面抗原、蛋白質(zhì)和各種生物活性分子,在細(xì)胞通訊中發(fā)揮關(guān)鍵作用[7]。有研究發(fā)現(xiàn),EPC來源MPs可以保護(hù)心肌細(xì)胞免受血管緊張素-Ⅱ誘導(dǎo)的肥大和凋亡[8],并將微小RNA轉(zhuǎn)移到EPC和EC,修復(fù)CVD中的組織損傷[9]。WU等[10]發(fā)現(xiàn)miR-126通過細(xì)胞外調(diào)節(jié)蛋白激酶(extracellular signal-regulated kinase,ERK)1/2信號通路來增強(qiáng)氧化應(yīng)激下EPC的生物學(xué)功能,這可能為急性心肌梗死的治療提供新的策略。然而,損傷的EPC來源MPs對EPC的作用及具體作用機(jī)制尚缺乏深入研究,因此,本研究通過體外實(shí)驗(yàn),損傷處理EPC,提取MPs對EPC進(jìn)行干預(yù),為減少細(xì)胞組織損傷,開發(fā)阻斷損傷EPC來源MPs試劑盒提供理論依據(jù),現(xiàn)報(bào)道如下。
1 材料與方法
1.1 一般材料
EPC購于武漢普諾賽生命科技有限公司;異硫氰酸熒光素(fluorescein isothiocyanate,FITC)-CD34(11-0349-42)購于美國eBioscience公司;內(nèi)皮細(xì)胞生長培養(yǎng)基(endothelial cell growth medium,EGM)-2(Promocell,C22121)、胎牛血清(Wisent,086-150)及青鏈霉素(Wisent,450-201-EL)均購于北京畢特博生物技術(shù)有限責(zé)任公司;腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)重組蛋白購于上海澤葉生物科技有限公司。檢測設(shè)備:AcurriC6流式細(xì)胞儀購于美國BD公司;JEM1011透射電鏡儀購于日本JEOL公司;Multiskan 51119000酶標(biāo)儀購于美國Thermo fisher公司;Ts2R倒置顯微鏡購于日本Nikon公司);Veriti PCR儀購于美國Applied Biosystems公司;Mini Protean 3十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)儀器購于美國Bio-Rad公司;化學(xué)發(fā)光成像分析儀購于上海天能生命科學(xué)有限公司。
1.2 方法
1.2.1細(xì)胞培養(yǎng)
EPC為貼壁細(xì)胞,培養(yǎng)于EGM-2中,含有10%胎牛血清,100 U/mL的青霉素,100 μg/mL鏈霉素,放置于37 ℃、5% CO2的細(xì)胞培養(yǎng)箱中。當(dāng)細(xì)胞長至80%~90%時(shí),去除培養(yǎng)基,加入磷酸鹽緩沖液(PBS)洗1~2次,去除PBS后,加入1 mL胰酶消化1~3 min后,加入3 mL完全培養(yǎng)基中和胰酶終止消化,將消化的細(xì)胞轉(zhuǎn)移至15 mL離心管中,1 000 r/min離心5 min。倒掉上清液,加入3 mL培養(yǎng)基重懸細(xì)胞,1∶3傳代至培養(yǎng)皿中培養(yǎng)。
1.2.2流式細(xì)胞儀檢測
取分離培養(yǎng)的EPC,去除培養(yǎng)基,加入PBS洗1~2次,去除PBS后加入1 mL胰酶消化1~3 min,加入3 mL完全培養(yǎng)基中和胰酶終止消化,將消化的細(xì)胞轉(zhuǎn)移至15 mL離心管中,1 000 r/min離心5 min。倒掉上清液后,加入3 mL PBS重懸細(xì)胞。加入PBS-1%牛血清白蛋白(BSA)重懸細(xì)胞,加入CD34抗體按照1∶50的比例稀釋,空白對照細(xì)胞加入同型對照抗體,冰上孵育30 min。細(xì)胞中加入400 μL PBS混勻,1 000 r/min離心,去除上清液,加入PBS洗3次,加入500 μL PBS緩沖液重懸細(xì)胞。CD34是EPC的特征性分子標(biāo)志物,采用流式細(xì)胞儀檢測分離細(xì)胞中的CD34陽性細(xì)胞群,見圖1。

圖1 EPC流式鑒定
1.2.3HG、TNF-α損傷處理
將EPC培養(yǎng)于培養(yǎng)皿中,待細(xì)胞貼壁后分別進(jìn)行正常培養(yǎng)、高糖(high glucose,HG)培養(yǎng)、加入TNF-α重組蛋白培養(yǎng)24 h后收集上清液并提取MPs,此為HG、TNF-α損傷的EPC來源MPs。將1×106個(gè)正常EPC接種于6孔板中,待細(xì)胞貼壁后,將50 μg/mL正常培養(yǎng)EPC來源MPs(control-EPC-MPs組)、50 μg/mL HG培養(yǎng)EPC來源MPs(HG-EPC-MPs組)、50 μg/mL TNF-α培養(yǎng)EPC來源MPs(TNF-α-EPC-MPs組)分別加入正常培養(yǎng)的EPC后,收集細(xì)胞進(jìn)行檢測。
1.2.4透射電鏡檢測
3組細(xì)胞用胰酶消化后,收集到離心管中,2 000 r/min離心5 min,去除培養(yǎng)基。向收集的細(xì)胞中加入戊二醛稀釋液固定備用;取5~10 μL細(xì)胞懸液加到Formvar-carbon載樣銅網(wǎng)上;取100 μL PBS加到封口膜上,用鑷子將銅網(wǎng)(Formvar膜面朝下)放在PBS液滴上清洗。在所有步驟中,保持Formvar膜面濕潤而另一面干燥;銅網(wǎng)放在50 μL 1%戊二醛液滴上5 min;銅網(wǎng)放在100 μL ddH2O洗2 min(洗8次);銅網(wǎng)放在50 μL草酸雙氧鈾液滴上(pH 7.0)5 min;銅網(wǎng)放在50 μL甲基纖維素液滴上10 min,冰上操作;銅網(wǎng)放到樣品臺頂端的不銹鋼環(huán)上,在濾紙上吸去多余液體。空氣干燥5~10 min;銅網(wǎng)放在樣品盒里,拍攝透射電鏡照片。
1.2.5MTT檢測
3組細(xì)胞培養(yǎng)24 h后,加入10 μL MTT(5 mg/mL),在細(xì)胞培養(yǎng)箱中繼續(xù)培養(yǎng)3 h。去除培養(yǎng)基,每孔加入100 μL Formazan溶解液,適當(dāng)混勻,在細(xì)胞培養(yǎng)箱內(nèi)繼續(xù)孵育,直至普通光學(xué)顯微鏡下觀察發(fā)現(xiàn)Formazan全部溶解。37 ℃孵育4 h,紫色結(jié)晶全部溶解。如果紫色結(jié)晶較小、較少,溶解時(shí)間較短;如果紫色結(jié)晶較大、較多,溶解時(shí)間延長,可適當(dāng)振搖數(shù)次。在570 nm測定吸光度(A)值;如無570 nm濾光片,可使用560~600 nm濾光片。
1.2.6細(xì)胞劃痕實(shí)驗(yàn)
將5×106個(gè)EPC接種于6孔板中,搖晃均勻后放入培養(yǎng)箱中培養(yǎng)。待細(xì)胞長滿后,去除培養(yǎng)基,用PBS洗1次;200 μL槍頭在6孔板中劃3道直線,PBS洗2次,去掉劃出的細(xì)胞;用PBS洗2次,將殘余的細(xì)胞洗干凈;在顯微鏡下拍照,加入無血清培養(yǎng)基,再分別加入control-EPC-MPs組、HG-EPC-MPs組、TNF-α-EPC-MPs組細(xì)胞,搖晃均勻后放入培養(yǎng)箱中常規(guī)培養(yǎng)12、24 h后拍照。
1.2.7細(xì)胞管腔形成實(shí)驗(yàn)
3組細(xì)胞用胰酶消化,1 000 r/min離心,去除上清液,加入完全培養(yǎng)基重懸細(xì)胞,將細(xì)胞分別種在24孔板中,每孔種1×105個(gè)細(xì)胞。培養(yǎng)6 h后,觀察EC管腔形成情況并拍照。拍照后,使用Image J軟件Angiogenesis Analyzer Plugin插件對管腔形成情況進(jìn)行分析。
1.2.8實(shí)時(shí)熒光定量PCR(qPCR)檢測
3組細(xì)胞去除培養(yǎng)基,加入PBS洗1次,加入1 mL TRIzol裂解細(xì)胞,將裂解液轉(zhuǎn)移至EP管中。加入氯仿200 μL,用力振搖15 s,室溫靜置15 min,4 ℃ 12 000 r/min離心15 min。離心后液體分為3層,小心吸取上層無色液體移入1個(gè)新的EP管中。加入等體積異丙醇,上下顛倒混勻,室溫靜置5 min,4 ℃ 12 000 r/min離心10 min。去除上清液,沉淀加入75%乙醇1 mL,輕微振蕩15 s,4 ℃ 7 500 r/min離心5 min。小心去除上清液,管內(nèi)沉淀在超凈臺中鼓風(fēng)靜置干燥3~5 min。加入50 μL DEPC水溶解,nanodrop檢測濃度水平,-80 ℃冰箱保存。RNA純度根據(jù)nanodrop檢測結(jié)果和230、260、280 nmA值決定。內(nèi)皮型一氧化氮合成酶(endothelial nitric oxide synthase,eNOS)、沉默信息調(diào)節(jié)因子1(silent information regulator 1,SIRT1)、ERK、大鼠肉瘤(rat sarcoma,RAS)引物情況見表1。

表1 PCR檢測引物序列
1.2.9Western blot
3組細(xì)胞加入200 μL RIPA裂解液,冰上裂解30 min。裂解后將裂解產(chǎn)物轉(zhuǎn)移到預(yù)冷的1.5 mL離心管(冰上)并在4 ℃下以12 000 r/min離心10 min。將離心后的上清液轉(zhuǎn)移至1.5 mL離心管中,-20 ℃保存。采用BCA法檢測總蛋白水平,十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate,polyacrylamide gel electrophoresis,SDS-PAGE)分離蛋白。濃縮膠黏劑層電壓為80 V,電泳時(shí)間為30 min。樣品進(jìn)入分離膠黏劑后,在120 V下電泳90 min,然后將蛋白轉(zhuǎn)移到硝酸纖維素膜上。在脫脂乳中封閉后,將膜與一抗在4 ℃孵育過夜。根據(jù)一抗,將膜與二抗在25 ℃孵育2 h。使用化學(xué)發(fā)光成像分析儀進(jìn)行掃描成像。
1.3 統(tǒng)計(jì)學(xué)處理
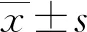
2 結(jié) 果
2.1 3組EPC電鏡下細(xì)胞器變化
control-EPC-MPs組細(xì)胞微管結(jié)構(gòu)完整,整體較長,高爾基體結(jié)構(gòu)完整;HG-EPC-MPs組細(xì)胞微管結(jié)構(gòu)完整,長度無明顯差異,高爾基體結(jié)構(gòu)相對完整;TNF-α-EPC-MPs微管結(jié)構(gòu)完整,長度有所縮短,高爾基體結(jié)構(gòu)相對完整,部分高爾基體出現(xiàn)囊泡狀散在分布在細(xì)胞質(zhì)中,見圖2。

綠色箭頭:微管;藍(lán)色箭頭:高爾基體。
2.2 3組EPC細(xì)胞增殖情況比較
與control-EPC-MPs組比較,HG-EPC-MPs組、TNF-α-EPC-MPs組細(xì)胞增殖率明顯下調(diào),差異有統(tǒng)計(jì)學(xué)意義(P<0.01),見圖3。

①:control-EPC-MPs組;②:HG-EPC-MPs組;③:TNF-α-EPC-MPs組;a:P<0.01,與control-EPC-MPs組比較。
2.3 3組EPC細(xì)胞遷移情況比較
與control-EPC-MPs組比較,HG-EPC-MPs組、TNF-α-EPC-MPs組0 h細(xì)胞遷移能力無明顯變化(P>0.05),12、24 h細(xì)胞遷移能力明顯降低,差異有統(tǒng)計(jì)學(xué)意義(P<0.01),見圖4。

A:細(xì)胞劃痕圖(100×);B:3組EPC遷移距離比較;a:P<0.01,與control-EPC-MPs組比較。
2.4 3組EPC管腔形成情況比較
與control-EPC-MPs組比較,HG-EPC-MPs組、TNF-α-EPC-MPs組成管明顯減少,差異有統(tǒng)計(jì)學(xué)意義(P<0.01),見圖5。

A:管腔形成圖(100×);B:成管分析結(jié)果;①:control-EPC-MPs組;②:HG-EPC-MPs組;③:TNF-α-EPC-MPs組;a:P<0.01,與control-EPC-MPs組比較。
2.5 3組EPC的mRNA和蛋白表達(dá)水平比較
與control-EPC-MPs組比較,HG-EPC-MPs組、TNF-α-EPC-MPs組eNOS、SIRT1、RAS和ERK的mRNA和蛋白表達(dá)水平下調(diào),差異有統(tǒng)計(jì)學(xué)意義(P<0.05),見圖6~7。

A:3組eNOS mRNA表達(dá)比較;B:3組SIRT1 mRNA表達(dá)比較;C:3組RAS mRNA表達(dá)比較;D:3組ERK mRNA表達(dá)比較;①:control-EPC-MPs組;②:HG-EPC-MPs組;③:TNF-α-EPC-MPs組;a:P<0.05,與control-EPC-MPs組比較。

A:3組eNOS、RAS Western blot圖;B:3組SIRT1、ERK Western blot圖;C:3組eNOS蛋白表達(dá)分析;D:3組SIRT1蛋白表達(dá)分析;E:3組RAS蛋白表達(dá)分析;F:3組ERK蛋白表達(dá)分析;①:control-EPC-MPs組;②:HG-EPC-MPs組;③:TNF-α-EPC-MPs組;a:P<0.05,與control-EPC-MPs組比較。
3 討 論
從骨髓動員進(jìn)入循環(huán)的EPC對血管修復(fù)、管壁組織重塑和血管生成[11]具有重要意義。前期研究發(fā)現(xiàn),MPs在CVD的發(fā)生、發(fā)展中發(fā)揮極其重要的作用[12]。研究人員已經(jīng)證實(shí),EPC來源MPs可以修復(fù)CVD中的組織損傷[9-10]。本研究應(yīng)用HG和TNF-α介導(dǎo)EPC損傷來源MPs,探索EPC功能障礙的機(jī)制,這可能為將來阻斷EPC損傷來源MPs、減少EPC損傷提供理論依據(jù)。
本研究發(fā)現(xiàn),HG和TNF-α介導(dǎo)EPC損傷來源MPs下調(diào)了EPC細(xì)胞增殖,降低了細(xì)胞遷移能力,細(xì)胞成管明顯減少;HG-EPC-MPs組和TNF-α-EPC-MPs組EPC損傷加重,并引起EPC微管和高爾基體等細(xì)胞器輕微改變;HG-EPC-MPs組和TNF-α-EPC-MPs組EPC變化可能與SIRT1/ERK1信號通路有關(guān)。有研究強(qiáng)調(diào)了線粒體和內(nèi)質(zhì)網(wǎng)在維持內(nèi)皮功能中的核心作用[13]。然而,關(guān)于微管和高爾基體在內(nèi)皮功能中的作用鮮有報(bào)道。本研究發(fā)現(xiàn),HG-EPC-MPs組、TNF-α-EPC-MPs組的EPC微管結(jié)構(gòu)完整,長度有所縮短,高爾基體結(jié)構(gòu)相對完整,部分高爾基體出現(xiàn)囊泡狀散在分布在細(xì)胞質(zhì)中,提示HG和TNF-α介導(dǎo)EPC損傷來源的MPs可能會導(dǎo)致微管和高爾基體結(jié)構(gòu)改變,從而加重應(yīng)激損傷反應(yīng)。因此,高爾基體損傷可能在EPC的損傷中發(fā)揮重要作用。
SIRT1是哺乳動物NAD依賴的sirtuin脫乙酰基酶組成部分,影響炎癥、線粒體生物發(fā)生、細(xì)胞衰老和凋亡等多個(gè)過程[14-15]。CAO等[16]證明了原兒茶酚醛通過調(diào)節(jié)SIRT1誘導(dǎo)自噬和抑制凋亡來挽救氧糖剝奪/再灌注(oxygen-glucose deprivation/reperfusion,OGD/R)誘導(dǎo)的EC損傷。此外,LI等[17]提出miR-126過表達(dá)通過促進(jìn)SIRT1/Nrf2信號通路來減輕氧化應(yīng)激和炎癥反應(yīng),從而減輕OGD/R誘導(dǎo)的人臍靜脈EC(human umbilical vein endothelial cells,HUVECs)的神經(jīng)毒性。本研究中,與control-EPC-MPs組比較,HG-EPC-MPs組、TNF-α-EPC-MPs組SIRT1的mRNA和蛋白表達(dá)水平下調(diào)。此外, HG-EPC-MPs組和TNF-α-EPC-MPs組的EPC增殖率、細(xì)胞遷移能力降低,細(xì)胞成管明顯減少。BI等[18]證明,血管生成素-1通過Tie2受體/ERK1/2-p38MAPK通路減輕了腎小球EC中內(nèi)質(zhì)網(wǎng)應(yīng)激誘導(dǎo)的細(xì)胞功能障礙和凋亡。WANG等[19]研究表明,堿性成纖維細(xì)胞生長因子參與了缺血氧化損傷模型中內(nèi)質(zhì)網(wǎng)應(yīng)激和線粒體功能障礙的抑制,這是PI3K/Akt和ERK1/2信號通路激活的基礎(chǔ)。與control-EPC-MPs組比較,HG-EPC-MPs組、TNF-α-EPC-MPs組ERK1的mRNA及其蛋白表達(dá)下調(diào),表明HG和TNF-α介導(dǎo)EPC損傷來源的MPs可以通過影響SIRT1/ERK1的表達(dá)和改變高爾基體結(jié)構(gòu)來抑制EPC的生物學(xué)功能。
eNOS是一種產(chǎn)生NO的酶,在心肌缺血再灌注損傷(myocardial ischemia reperfusion injury,MIRI)中具有重要意義。CAPE-oNO2通過SIRT1/eNOS通路抑制氧化應(yīng)激、炎癥反應(yīng)和細(xì)胞壞死,從而減輕MIRI[20]。PENG等[21]證明CC基元配體2促進(jìn)MAPK/ERK1/2/MMP-9、PI3K/AKT和Wnt/β-catenin信號通路相關(guān)蛋白的表達(dá),并以濃度依賴方式促進(jìn)HUVECs的增殖、遷移和管形成。醛糖還原酶通過SIRT1/AMPK-α1/mTOR通路抑制HG誘導(dǎo)的eNOS和血管細(xì)胞黏附因子1表達(dá),調(diào)控單核細(xì)胞白血病單核細(xì)胞與HUVECs的黏附,從而調(diào)控HG誘導(dǎo)的HUVECs死亡[22]。本研究中,與control-EPC-MPs組比較,HG-EPC-MPs組、TNF-α-EPC-MPs組SIRT1的mRNA和蛋白表達(dá)水平下調(diào)。RAS的激活在MIRI的發(fā)生、發(fā)展中發(fā)揮重要作用[23]。當(dāng)MIRI發(fā)生時(shí),腎素-血管緊張素-醛固酮系統(tǒng)的最終代謝產(chǎn)物血管緊張素Ⅱ與血管緊張素Ⅰ型受體結(jié)合,刺激線粒體氧化應(yīng)激損傷反應(yīng),導(dǎo)致活性氧過度產(chǎn)生[24]。活性氧過度產(chǎn)生可能誘發(fā)強(qiáng)烈的炎癥反應(yīng),增加心肌梗死面積,導(dǎo)致心功能異常。LI等[25]研究表明,高爾基體參與了鈣超載的形成和緩解,并對神經(jīng)細(xì)胞的缺血和再灌注有強(qiáng)烈的反應(yīng)。因此,本研究證實(shí)HG和TNF-α介導(dǎo)EPC損傷來源MPs可能通過影響EPC中eNOS、RAS的表達(dá),促進(jìn)氧化應(yīng)激、炎癥反應(yīng)和細(xì)胞壞死,從而加重EPCs的損傷。
綜上所述,HG和TNF-α介導(dǎo)EPC損傷來源MPs可能通過調(diào)控SIRT1/ERK1通路蛋白的表達(dá),使EPC內(nèi)含細(xì)胞器的結(jié)構(gòu)改變,從而影響EPC的生物學(xué)功能。
猜你喜歡
中國設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
