牛Basigin基因變異體的克隆與鑒定
2022-12-29 07:32:10劉雪艷程林圓周國利
湖北農業科學 2022年20期
劉雪艷,程林圓,周國利
(聊城大學生命科學學院,山東 聊城 252059)
選擇性剪接(Alternative splicing,AS)是擴大真核細胞遺傳多樣性的一種普遍機制[1]。大約90%的人類基因通過這個精細調控的過程產生一個以上的轉錄本[2,3]。基因的前體mRNA通過不同的剪接方式產生不同的mRNA剪接變異體,通過剪接因子和相應轉錄組中順式作用元件之間的相互作用,進而精細調控組織和階段特異性剪接事件,可變剪接是調節基因表達和產生蛋白質組多樣性的重要機制,是導致真核生物基因和蛋白質種類多樣性和差異性的重要原因[4]。AS轉錄本的時空表達譜在很大程度上有助于細胞分化、特化和器官發生[5]。
選擇性多聚腺苷酸化(Alternative polyadenylation,APA)是真核生物細胞中普遍存在的一種基因調控機制,選擇不同的加尾信號可以使單個基因產生不同的轉錄本[6,7]。APA的結果是可以產生具有不同編碼序列的轉錄本,或者不同長度的3'端非翻譯區(3'untranslated region,3'UTR),而不同長度的3'UTR影響著mRNA的穩定性、定位和翻譯效率,因此,APA在細胞發育、分化和增殖過程中對基因表達具有重要的調控作用[8,9]。
牛的Basigin(BSG)基因,位于7號染色體上,長約9 kb,含有7個外顯子。BSG基因編碼一種廣泛表達的跨膜糖蛋白Basigin,也稱CD147,它參與機體腫瘤發生、胚胎發育、能量代謝、損傷修復等多種生理和病理過程[10]。人和小鼠的BSG基因剪接變異體已經被克隆[11,12],然而牛的BSG基因是否存在AS和APA變異體尚不清楚。因此,本研究的目的是通過3'RACE技術和測序技術去尋找牛的BSG基因是否存在AS或APA的生物學事件。研究結果將會補充牛的基因組數據,為進一步分析牛BSG基因不同變異體在不同的生物學過程中的功能奠定分子生物學基礎。
1 材料與方法
1.1 試劑
RNAeasy?Plus動物RNA抽提試劑盒、BeyoRT?cDNA第一鏈合成試劑盒購自上海碧云天生物技術有限公司;瓊脂糖、凝膠DNA回收試劑盒均購自天根生化科技公司;Taq酶、dNTPs、T載體PCR產物克隆試劑盒購自生工生物工程(上海)有限公司;其他試劑均為國產普通分析純。
1.2 動物組織樣品及總RNA提取
魯西黃牛的組織樣品采自山東省聊城市張爐集鎮屠宰場。在屠宰后立即采集牛的里脊、睪丸、腎臟、肝臟等組織,并迅速投到液氮中帶回實驗室,利用RNA試劑盒提取總RNA,用濃度為1%~1.5%的瓊脂糖凝膠電泳檢測總RNA的完整性。
1.3 BSG基因變異體的克隆與測序
根據NCBI中牛BSG基因的mRNA序列(Gen-Bank登錄號:NM_001075371.2),利用Oligo 6.0軟件設計用于牛BSG基因變異體克隆的2條上游特異性引物,第一個引物在第二個引物的下游。經BLAST程序比對,各條引物在牛的基因組上均沒有互補序列。
利用表1中所設計的用于3'RACE的cDNA反轉錄通用引物,用cDNA第一鏈合成試劑盒反轉錄合成相應組織的cDNA,然后以獲得的cDNA為模板進行PCR。PCR產物通過凝膠回收試劑盒純化回收,利用連接試劑盒將回收的PCR產物克隆到T載體上,陽性的克隆進行雙向測序,再用DNAStar 5.0軟件、NCBI和UCSC網站的BLAST程序作序列比對分析。
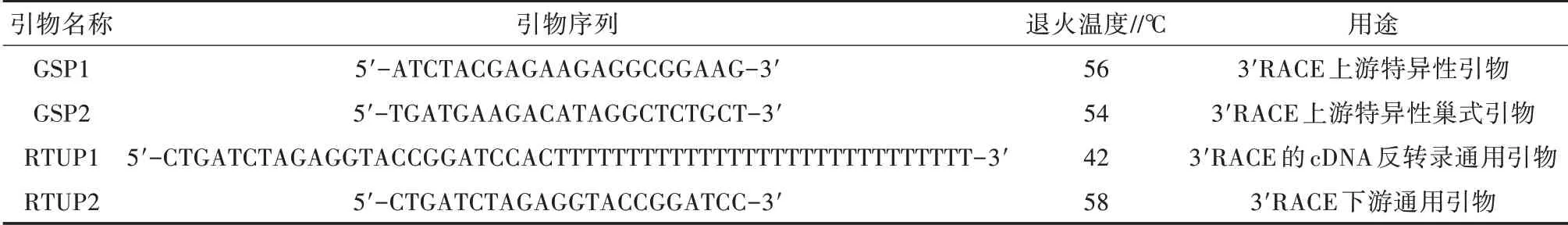
表1 BSG基因3'RACE相關引物信息
2 結果與分析
2.1 組織樣品總RNA電泳檢測
分別從肝、睪丸、腎、里脊組織中提取總RNA,通過瓊脂糖凝膠電泳檢測總RNA的完整性。電泳結果顯示,提取總RNA的28S rRNA與18S rRNA的條帶比較明顯,5S條帶比較暗,說明提取的RNA完整性較好,沒有明顯的降解(圖1),可以用于后續的相關試驗。
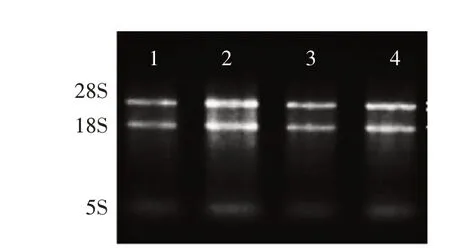
圖1 各組織樣品總RNA電泳檢測
2.2 3'RACE產物的電泳及測序
2.2.1 3'RACE產物電泳 通過濃度為1.5%的瓊脂糖凝膠電泳結果(圖2)可以看出,擴增出的目標片段至少為3條,可能是由于基因選擇性表達,它們并沒有來源于同一個組織。在腎組織的PCR擴增產物中,有2條比較明顯條帶,700 bp左右的條帶亮度明顯大于300 bp,并且在100 bp附近也能觀察到1條與里脊中大小相似的條帶,但是條帶的亮度比較弱。在里脊組織中有1條亮度明顯的約100 bp的條帶,但該條帶較寬,不能確定該條帶存在幾條目的片段,可以通過后續的克隆和測序驗證。
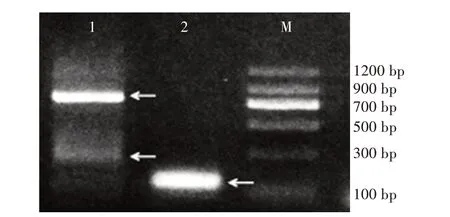
圖2 3'RACE產物電泳
2.2.2 3'RACE產物片段測序 通過利用上游GSP1、GSP2特異性引物與下游RTUP2通用引物進行巢式PCR,PCR產物經回收、克隆和測序后發現有4條目的片段,根據片段的大小將它們分別命名為BSG-v1、BSG-v2、BSG-v3和BSG-v4,各片段的部分序列及峰圖見圖3。
BSG-v1片段的組織來源為牛的腎臟組織,上下游引物之間長767 bp,在poly(A)尾上游17~23 bp處存在1個經典的加尾信號“AATAAA”(圖3a)。利用NCBI中的BLAST程序對該序列與牛的基因組進行比對,發現與牛BSG基因的序列完全一致,并且所獲得的BSG-v1就是NCBI中已經公布的序列(Gen-Bank登錄號:NM_001075371.2)。BSG-v2片段的組織來源為牛的腎臟組織,上下游引物之間長245 bp,在poly(A)尾上游12~18 bp處存在1個經典的加尾信號“AATAAA”,該加尾信號序列與BSG-v1是同一個加尾信號(圖3b)。該序列與牛的基因組比對,發現與牛BSG基因的序列高度同源。BSG-v3和BSGv4片段來源于牛的里脊組織,上下游引物之間分別為168 bp和129 bp,具有明顯的poly(A)尾結構,但無經典的加尾信號(圖3c,3d)。該序列與牛的基因組比對,發現BSG-v3和BSG-v4序列與牛BSG基因序列高度同源。
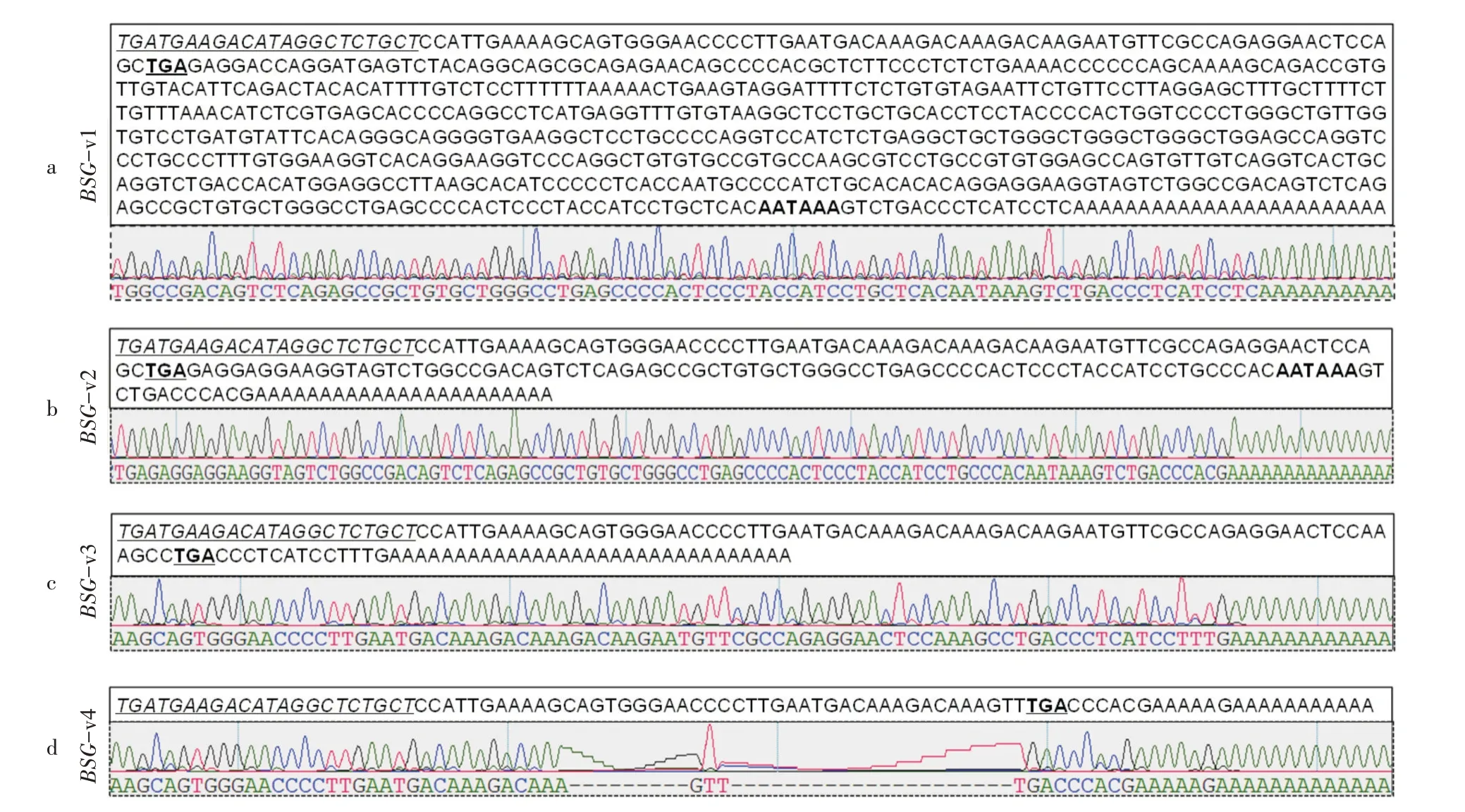
圖3 牛BSG基因各轉錄本變異體的部分序列與測序峰
通過使用NCBI比對發現,只有BSG-v1序列與已經報道的完全一致,BSG-v2,BSG-v3和BSG-v4是新發現的可變剪接變異體。
2.2.3 牛BSG基因的可變剪接變異體的結構分析利用NCBI網站上的BLAST程序和UCSC網站(http://genome.ucsc.edu/)上的BLAT程序聯合分析發現,與BSG-v1相比,BSG-v2、BSG-v3和BSG-v4是BSG-v1不同形式的變異體(圖4)。BSG-v2在第7外顯子處發生了A3SS,導致BSG-v2的外顯子7比BSG-v1的短了515 bp,相當于BSG-v2的內含子6在原來的基礎上延伸了515 bp,但分析發現BSG-v2的內含子6的邊界仍然符合“GT-AG”規則(圖4a)。此外,通過對來源于里脊組織的2個可變剪接變異體BSG-v3和BSG-v4的分析發現,二者都發生了APA現象,它們都沒有外顯子7,導致產生了更短的3'UTR(圖4b,4c)。進一步地分析發現,與BSG-v1和BSG-v2相比,雖然終止密碼子的位置發生了變動,但并沒有造成原有開放閱讀框(ORF)移碼突變,而是密碼子堿基以3的倍數形式缺失,造成羧基端變化。BSG-v3終止密碼子前2個密碼子與BSG-v1和BSG-v2不同,并且BSG-v3多了1個編碼氨基酸的密碼子(圖4b)。而BSG-v4在3'端缺失了9個編碼氨酸的密碼子,使BSG的羧基端變短(圖4c)。而且BSG-v1和BSG-v2的3'UTR區都非常短,無經典的加尾信號,具有明顯的poly(A)尾巴。
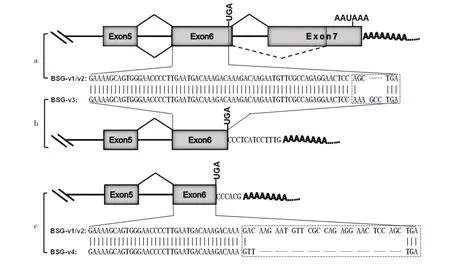
圖4 牛BSG基因各轉錄本變異體的部分結構與序列
3 小結與討論
在真核生物中,AS和APA的現象是普遍存在的[9,13],本研究通過3'RACE技術成功地克隆出了牛BSG基因的4條轉錄本的3'端片段。經序列對比和分析后發現,在這4條轉錄本中,最長的1條BSG-v1已經在NCBI中公布。BSG-v2則在外顯子7中(也是3'UTR)存在明顯的A3SS剪接現象。較短的2條BSG-v3和BSG-v4都分別存在1個新的poly(A)位點,但這4條不同的轉錄本并沒有來源于牛的同一個組織,這可能是基因的選擇性表達造成的。同時這也是AS和APA在真核細胞中的遺傳多樣性精細調控的一種體現[13]。
在本研究中,BSG-v2的外顯子7有515 bp的序列被剪接,與BSG-v1的內含子6一起作為新的內含子6被剪接掉,因為發生剪接的是外顯子7的5'端,并且屬于3'UTR,所以BSG-v2變異體的加尾信號并沒有改變(圖4a)。推測BSG-v1和BSG-v2可能會存 在 不 同 的 轉 錄 后 調 控 機 制[13,14]。MicroRNAs(miRNAs)是在真核生物中發現的一類內源性的具有調控基因表達功能的非編碼RNA,其大小長約20~25個核苷酸。成熟的miRNAs由較長的初級轉錄物經過一系列核酸酶的剪切加工而產生,隨后組裝進RNA誘導的沉默復合體,通過堿基互補配對的方式識別靶mRNA,并根據互補程度的不同指導沉默復合體降解靶mRNA或者阻遏靶mRNA的翻譯。成熟的miRNA結合到與其互補的mRNA的位點來調控基因表達,其中一些靶位點就存在于3'UTR[15]。研究發現,與一般3'UTR相比,一些基因通過內含子保留形成的3'UTR區域比其他區域有更大的可能含有更多miRNA的靶標位點,因此,保留內含子能顯著增加該基因被miRNA調控的可能[16]。一些研究已經報道,基因的AS或APA的事件會導致該基因逃避某些miRNAs或RNA結合蛋白(RNA binding proteins,RBPs)的調控[15-17]。所以由于BSG-v2變異體3'UTR的A3SS剪接事件存在,可能會使該變異體逃避miRNAs的調控,從而導致牛BSG-v1和BSG-v2的差異表達調控;此外,基因的3'UTR除受miRNAs的轉錄后調控外,還會有一些RBPs的結合,從而促進或抑制mRNA的穩定性或翻譯效率。與BSG-v1相比,BSG-v2的3'UTR長度短了515 bp,因此可能會缺失RBPs的結合位點,從而導致mRNA或蛋白水平上的差異。但具體逃避了哪些能夠與BSG-v2變異體的3'UTR發生相互作用的miRNAs或RBPs,還需通過相應的試驗進一步證實。
此外,有趣的是本次克隆BSG基因mRNA的3'端片段表明,來源于牛里脊組織的2條較短轉錄本具有明顯的新poly(A)位點,有非常短的3'UTR序列,且都不包含外顯子7。BSG-v3比BSG-v1的ORF多一 個密碼 子,BSG-v4比BSG-v1的ORF缺 失9個密碼子,它們都是羧基端發生了變化,這種變化是否影響了BSG蛋白的功能,尚需進行功能驗證。但有報道指出,APA如果發生在編碼區,可能會引起終止密碼子前移,提前終止編碼蛋白[6]。此外,在BSG-v4的poly(A)尾中還有一個“G”存在,這也是近年來研究發現的在poly(A)尾中存在非“A”堿基的一種新事件,也已經成為近年來關于轉錄后調控研究的熱點[18]。AS同樣也可能會引起終止密碼子位置的改變,根據當前的研究發現,人的BSG基因通過AS選擇性編碼出4種變異體蛋白,分別為basigin-1、basigin-2、basigin-3和basigin-4[11]。但目前獲得的BSG-v3和BSG-v4變異體只是包括了外顯子5和外顯子6,至于其前面的外顯子是否發生了AS事件,還需要進一步證實。但本研究成功克隆了牛BSG基因幾種不同形式的變異體,為進一步探索BSG基因在牛的各種相關生物學過程中的表達調控及生物學功能奠定了一定的分子生物學基礎。
