連翹酯苷A通過TLR4/NF-κB p65/NLRP3軸對宮內感染所致新生大鼠肺損傷及免疫功能的影響研究
2022-09-20 04:14:58趙亞麗郎明瑤尹紅亞
重慶醫學 2022年17期
關鍵詞:模型
王 云,趙亞麗,郎明瑤,尹紅亞
(1.河北省保定市第二醫院兒科 071000;2.河北省保定市第二中心醫院兒科 072750;3.河北省兒童醫院婦產科,石家莊 050031)
宮內感染主要是由于病原微生物侵入羊膜腔導致胎盤、胎膜發生炎性反應引起,炎性因子的“瀑布式”級聯反應常引發新生兒肺功能損傷,與胎兒發生支氣管肺發育不良密切相關。連翹酯苷A可抑制感染H9N2禽流感病毒小鼠的炎性反應[1]。此外,連翹酯苷A可增強氧自由基清除力和免疫功能,抑制潰瘍性結腸炎大鼠炎性反應[2]。Toll樣受體4(Toll-like receptor 4,TLR4)/核因子-κB(nuclear factor-kappa B,NF-κB)p65信號通路激活可促進炎性因子釋放。研究發現,芪藶強心膠囊可抑制該信號通路,減輕心力衰竭大鼠心肌組織炎性反應[3]。本研究通過建立宮內感染所致新生大鼠肺損傷模型,連翹酯苷A灌胃治療,探討連翹酯苷A對肺損傷和免疫功能的影響,以及對TLR4/NF-κB p65/NOD樣受體3(NOD-like receptor 3,NLRP3)信號通路的調節作用,現報道如下。
1 材料與方法
1.1 材料
1.1.1實驗動物
健康清潔級雌性SD大鼠50只,8周齡,體重180~200 g;雄性SD大鼠25只,8周齡,體重190~210 g,購自北京維通利華實驗動物技術有限公司,許可證號:SCXK(京)2018-0001。
1.1.2試劑與儀器
連翹酯苷A(純度>98%)購自北京國家藥品和生物制品控制研究所;脂多糖購自美國Sigma公司;大腸桿菌購自中國獸醫藥品檢察所;白細胞介素(interleukin,IL)-6、IL-8和IL-1β ELISA試劑盒購自美國R&D公司;兔抗鼠TLR4、NF-κB p65、p-p65、NLRP3及GAPDH抗體購自美國Abcam公司;CD4+和CD8+單克隆抗體購自北京同立海源生物科技有限公司;酶標儀購自美國Bioteck公司;小動物呼吸功能分析系統(AniRes2005)購自北京貝蘭博科技公司。
1.2 方法
1.2.1模型制備[4]
下午4:00后,雌雄大鼠2∶1比例合籠,次日上午用棉簽擦拭雌鼠陰道,對其分泌物做涂片檢查,顯微鏡下觀察,以精子布滿視野記為妊娠第0天。取40只雌鼠在孕第15天,陰道擴張器暴露孕鼠子宮頸,6號注射器將0.2 mL大腸桿菌稀釋液注射入子宮頸肌肉內,兩側宮頸均進行注射。造模第2天起,將40只孕鼠分為模型組、連翹酯苷A組、脂多糖組和脂多糖+連翹酯苷A組,每組10只,連翹酯苷A組和脂多糖+連翹酯苷A組孕鼠灌胃連翹酯苷A 50 mg/kg,每天1次,脂多糖組和脂多糖+連翹酯苷A組孕鼠尾靜脈注射脂多糖 5 mg/kg,每天1次,直至自然分娩,各組新生大鼠選取10只用于實驗。
1.2.2宮內感染判斷標準
孕鼠分娩后,留取胎盤組織,預冷磷酸鹽緩沖液(PBS)清洗干凈后,進行蘇木素-伊紅(HE)染色,顯微鏡下觀察,若出現血管水腫、充血,中性粒細胞增多則說明發生宮內感染。
1.2.3新生大鼠呼吸頻率測定
記錄孕鼠分娩情況,7 d后記錄新生大鼠存活情況。于出生后第8天將新生大鼠麻醉,切開氣管后進行插管,通過導管連接呼吸機和信號調理器,使用肺功能檢測軟件記錄新生大鼠呼吸頻率。
1.2.4ELISA檢測新生大鼠血清中IL-8、IL-6和IL-1β濃度
呼吸頻率記錄完成后,斷頭處死,收集3 mL主動脈血,離心,吸取上清液。嚴格按照ELISA試劑盒說明書進行操作,酶標儀上測定波長490 nm處的吸光度(A)值。
1.2.5流式細胞儀檢測血清中CD4+、CD8+、CD4+/CD8+淋巴細胞數量
呼吸頻率記錄完成后,取3 mL新生大鼠主動脈血于流式管中,嚴格按照試劑盒說明書操作,流式細胞儀分析CD4+、CD8+及CD4+/CD8+比值。
1.2.6HE染色觀察新生大鼠肺組織病理損傷
采血結束后,迅速取出右側肺組織,進行HE染色,倒置顯微鏡下觀察并拍照,每張切片選取5個視野。
1.2.7Western blot檢測肺組織TLR4、p-p65、NF-κB p65和NLRP3蛋白相對表達水平
左側肺組織液氮中研磨,裂解,離心,吸取上清液,BCA法測定蛋白濃度,煮沸。上樣、電泳、濕轉、洗膜、封閉,TLR4、p-p65、NF-κB p65、NLRP3、GAPDH一抗4 ℃孵育過夜,洗膜,二抗孵育1 h,洗膜,ECL發光液顯色,Image J軟件分析蛋白條帶灰度值,目的蛋白相對表達水平=目的蛋白條帶灰度值/GAPDH蛋白條帶灰度值。
1.3 統計學處理
2 結 果
2.1 胎盤組織HE染色
模型組和脂多糖組孕鼠胎盤組織輪廓不清晰,大量炎性細胞浸潤;連翹酯苷A組孕鼠胎盤輪廓尚清晰,少量中性粒細胞浸潤;脂多糖+連翹酯苷A組孕鼠胎盤組織病理情況較脂多糖組減輕,較連翹酯苷A組加重,見圖1。
2.2 孕鼠分娩情況及新生大鼠存活情況
孕鼠分娩情況:模型組、脂多糖組和脂多糖+連翹酯苷A組孕鼠出現早產,均在孕第18~20天分娩,對照組和連翹酯苷A組孕鼠在孕第22~23天分娩,孕鼠無死亡。
新生大鼠存活情況:對照組新生大鼠87只,死亡2只,死亡率2.30%;模型組新生大鼠共85只,死亡8只,死亡率9.41%;連翹酯苷A組新生大鼠82只,死亡3只,死亡率3.65%;脂多糖組新生大鼠82只,死亡8只,死亡率9.75%;脂多糖+連翹酯苷A組新生大鼠80只,死亡5只,死亡率6.25%。
2.3 新生大鼠呼吸頻率、肺指數測定
與模型組比較,連翹酯苷A組呼吸頻率減慢、肺指數升高,脂多糖組呼吸頻率加快、肺指數降低(P<0.05);與脂多糖+連翹酯苷A組比較,連翹酯苷A組呼吸頻率減慢、肺指數升高,而脂多糖組呼吸頻率加快、肺指數降低(P<0.05),見表1。
2.4 ELISA檢測結果
與模型組比較,連翹酯苷A組IL-8、IL-6和IL-1β水平降低,脂多糖組升高(P<0.05);與脂多糖+連翹酯苷A組比較,連翹酯苷A組IL-8、IL-6和IL-1β水平降低,脂多糖組升高(P<0.05),見表2。
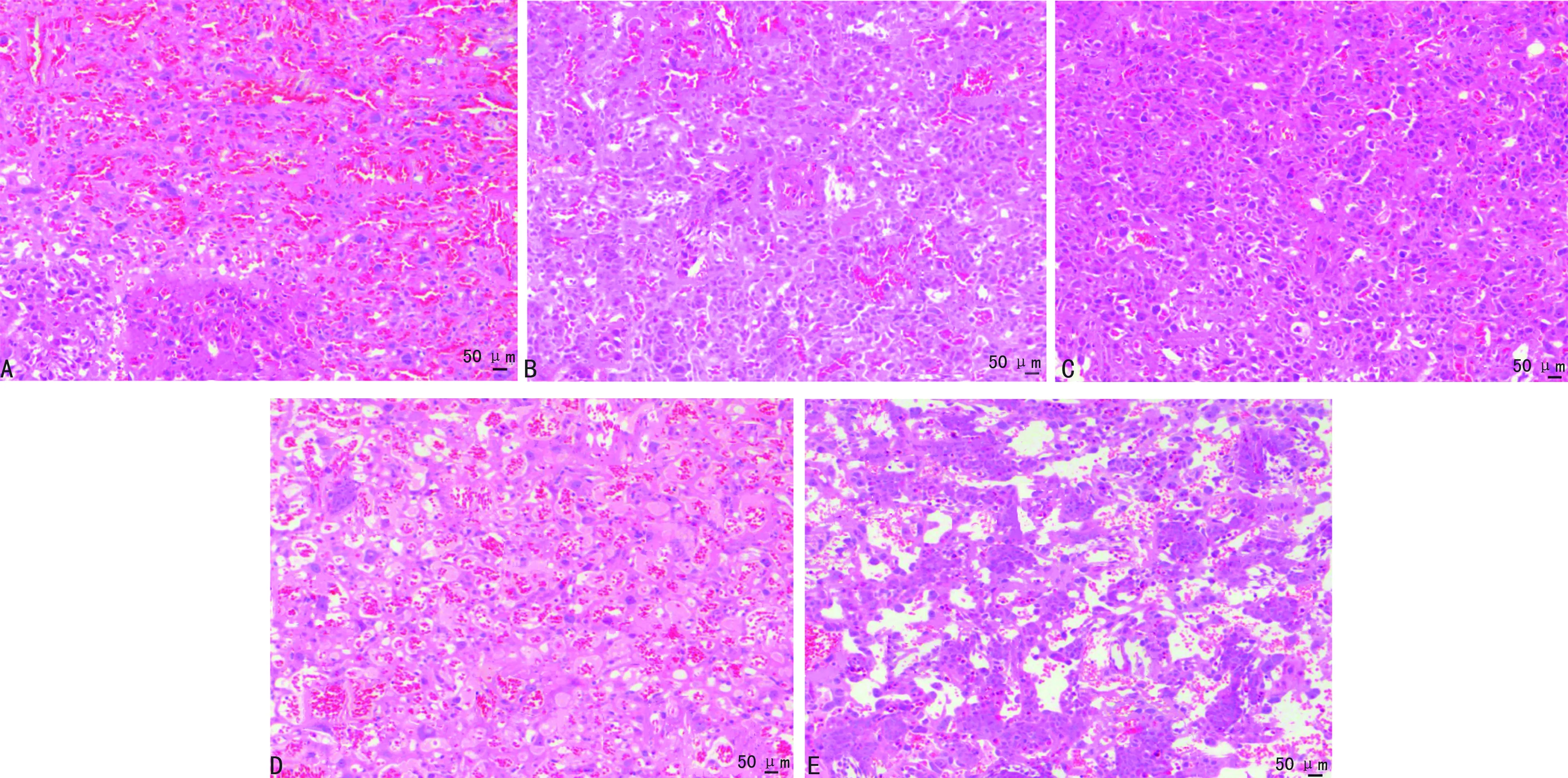
A:對照組;B:模型組;C:連翹酯苷A組;D:脂多糖組;E:脂多糖+連翹酯苷A組。
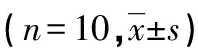
表1 新生大鼠呼吸頻率、肺指數比較
2.5 肺組織HE染色
模型組新生大鼠肺泡體積明顯增大,多數肺泡隔增厚,且可見大量炎性細胞浸潤;連翹酯苷A組肺泡稍擴張,少量肺泡隔增厚,少量炎性細胞浸潤;脂多糖組新生大鼠肺組織病理改變較模型組嚴重;脂多糖+連翹酯苷A新生大鼠肺組織病理變化較連翹酯苷A組加重,較脂多糖組減輕,見圖2。
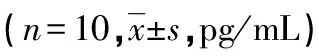
表2 血清IL-8、IL-6和IL-1β水平比較
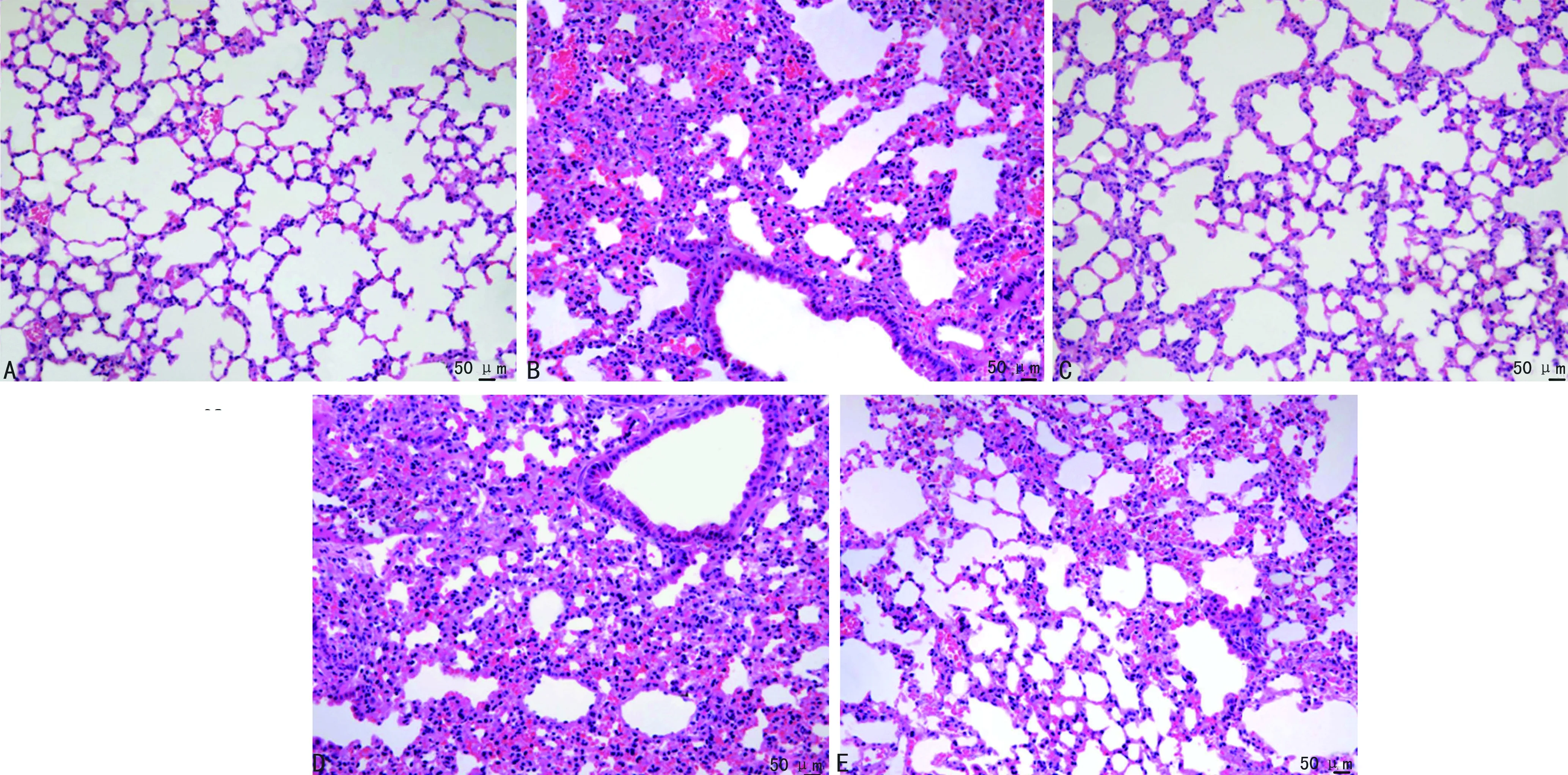
A:對照組;B:模型組;C:連翹酯苷A組;D:脂多糖組;E:脂多糖+連翹酯苷A組。
2.6 流式細胞儀檢測結果
連翹酯苷A組CD4+/CD8+(1.27±0.10)高于模型組(1.03±0.12),脂多糖組(0.82±0.17)低于模型組(P<0.05);脂多糖+連翹酯苷A組CD4+/CD8+(1.15±0.11)低于連翹酯苷A組,高于脂多糖組,差異有統計學意義(P<0.05)。
2.7 Western blot檢測結果
與模型組比較,連翹酯苷A組TLR4、p-p65和NLRP3蛋白相對表達水平降低,脂多糖組TLR4、p-p65和NLRP3蛋白相對表達水平升高(P<0.05);與脂多糖+連翹酯苷A組比較,連翹酯苷A組TLR4、p-p65和NLRP3蛋白相對表達水平降低,脂多糖組TLR4、p-p65和NLRP3蛋白相對表達水平升高(P<0.05),見表3、圖3。
表3 肺組織中TLR4、p-p65、NF-κB p65和NLRP3蛋白相對表達水平比較

A:對照組;B:模型組;C:連翹酯苷A組;D:脂多糖組;E:脂多糖+連翹酯苷A組。
3 討 論
宮內感染常通過胎盤垂直傳播造成胎兒先天性感染,胎齡越小,發生宮內感染的概率越高,是引起早產兒不良結局的重要原因[5]。支氣管肺發育不良是早產兒常發生的一種慢性肺部疾病,其主要的病理學改變為肺泡及肺微血管發育不良,而宮內感染引起羊水、胎盤和胎膜中炎性細胞積聚,并分泌大量炎性因子,被認為是支氣管肺發育不良的主要因素[6]。
ZHENG等[7]研究表明連翹酯苷A可調節免疫細胞平衡,降低甲型流感病毒FM1株在小鼠肺臟中引起的炎性反應。此外,連翹酯苷A可減輕脂多糖誘導的肺損傷[8]。T淋巴細胞是免疫反應的中心環節,CD4+和CD8+T淋巴細胞亞群之間比例失衡可導致免疫反應紊亂,從而引起炎性反應,造成組織損傷。IL-6是炎性反應中重要的炎性遞質,IL-8是啟動炎性反應的關鍵因子[9]。本研究結果顯示宮內感染造成新生大鼠呼吸頻率加快、肺指數降低,連翹酯苷A可減慢呼吸頻率、升高肺指數。宮內感染后新生大鼠肺泡數量明顯減少,且肺泡隔內可見大量炎性細胞浸潤,連翹酯苷A治療后少量炎性細胞浸潤;孕鼠宮內感染造成新生大鼠肺組織中CD4+/CD8+降低,炎性因子IL-6、IL-8和IL-1β水平升高,連翹酯苷A治療后CD4+/CD8+升高、炎性因子水平降低。此結果提示,連翹酯苷A可降低宮內感染新生大鼠炎性反應、改善免疫功能和肺損傷。
脂多糖作用于TRL4受體激活炎癥下游信號通路,NF-κB即為其重要的靶點之一[10]。NF-κB是多種炎性因子基因的靶點,通常以p65/p50組成的異源二聚體形式存在于細胞質中,一般情況下不表現其活性,當細胞受到刺激時,抑制因子IKB發生磷酸化降解,NF-κB解離并活化進入細胞核,調節下游因子表達[11]。NLRP3炎性小體是固有免疫信號通路中重要的多蛋白炎性體,細胞受到刺激時被活化,活化后的NLRP3炎性體切割含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)-1,使其成熟后促進炎性因子前體(pro-IL-1β、IL-18)成熟活化,發揮其生物功能,而炎性因子前體的產生依賴于NF-κB的激活[12]。研究發現,在脂多糖誘導的急性肺損傷小鼠模型中,肺組織p65、TLR4、NLRP3及caspase-1蛋白表達增加,肺組織損傷嚴重,而喘可治可減輕小鼠肺損傷,降低p65、TLR4、NLRP3及caspase-1蛋白在肺組織中的表達水平[13]。在脂多糖誘導的肺損傷小鼠模型中,脂多糖抑制TLR4/NF-κB p65信號通路激活及NLRP3炎性小體表達,降低氧化應激和炎性反應,改善肺功能障礙[14]。急性膿毒癥大鼠肺組織中p65和TLR4蛋白表達升高,肺水腫明顯,炎性因子水平升高,蘿卜硫素可降低p65和TLR4表達,降低肺水腫程度[15]。本研究通過脂多糖激活TRL4受體,探討連翹酯苷A是否通過該信號通路改善宮內感染新生大鼠肺損傷和免疫功能。結果顯示,脂多糖上調宮內感染新生大鼠肺組織中TLR4、p-p65和NLRP3蛋白表達,而連翹酯苷A治療后可抑制脂多糖的激活作用。此結果提示,連翹酯苷A可抑制TLR4/NF-κB p65/NLRP3信號通路激活。
綜上所述,連翹酯苷A可改善宮內感染新生大鼠肺損傷及免疫功能,其可能是通過抑制TLR4/NF-κB p65/NLRP3信號通路激活發揮作用,為臨床治療肺支氣管發育不良等疾病提供理論依據。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
