肉雞骨骼發(fā)育的調(diào)控機(jī)制
2022-08-05 06:09:22王翠月李明睿高玉時(shí)陳大偉
畜牧獸醫(yī)學(xué)報(bào) 2022年7期
關(guān)鍵詞:生長(zhǎng)
王翠月,李明睿,高玉時(shí),陳大偉*
(1. 江蘇省家禽科學(xué)研究所,揚(yáng)州 225125; 2. 山東農(nóng)業(yè)大學(xué),泰安 271018; 3. 山西農(nóng)業(yè)大學(xué),太谷 030801)
骨骼發(fā)育主要是通過(guò)生長(zhǎng)板處進(jìn)行的軟骨內(nèi)骨化完成的,該過(guò)程由多種調(diào)控因子組成的復(fù)雜調(diào)控網(wǎng)絡(luò)精準(zhǔn)調(diào)控以確保骨骼正常的生長(zhǎng)發(fā)育。骨骼起保護(hù)臟器、支撐體重和維持運(yùn)動(dòng)的作用,例如脛骨是肉雞主要的負(fù)重骨骼,強(qiáng)健的脛骨保證了肉雞正常的生命活動(dòng)。商品肉雞體重增長(zhǎng)迅速,骨骼生長(zhǎng)速度與體重增長(zhǎng)速度不匹配,腿部骨骼不足以支撐快速增長(zhǎng)的體重,尤其是胸肌的快速生長(zhǎng)使肉雞重心前傾,進(jìn)一步壓迫脛骨并損傷生長(zhǎng)板,使其更易患腿病。常見(jiàn)的肉雞腿病包括脛骨軟骨發(fā)育不良(TD)、外翻-內(nèi)翻畸形(VVD)和脛骨短粗等,病雞表現(xiàn)為行走困難、癱瘓、料肉比高、死淘率高等,嚴(yán)重阻礙了家禽養(yǎng)殖業(yè)的發(fā)展并引發(fā)了動(dòng)物福利問(wèn)題。然而,目前對(duì)于肉雞腿病發(fā)病的分子機(jī)制的研究較少且治療困難,綜述骨形成過(guò)程及參與骨骼發(fā)育的調(diào)控因子對(duì)于從骨骼發(fā)育角度探究肉雞腿病的發(fā)病機(jī)制以及制定合理治療方案至關(guān)重要。
1 骨形成過(guò)程
骨形成是一個(gè)嚴(yán)格的調(diào)控過(guò)程,包括膜內(nèi)成骨和軟骨內(nèi)成骨。膜內(nèi)成骨發(fā)生于胚胎時(shí)期,間充質(zhì)細(xì)胞濃縮凝聚并轉(zhuǎn)分化為成骨細(xì)胞,主要形成顱骨、鎖骨等。為保證軀體大多數(shù)骨骼的形成,特別是長(zhǎng)骨的發(fā)育,間充質(zhì)干細(xì)胞分化為軟骨細(xì)胞,軟骨細(xì)胞凝集形成軟骨基質(zhì)模板,即在骨骺處形成生長(zhǎng)板結(jié)構(gòu),通過(guò)軟骨內(nèi)骨化來(lái)調(diào)控骨骼發(fā)育。軟骨內(nèi)骨化是長(zhǎng)骨發(fā)育的第二階段,肉雞脛骨和股骨的發(fā)育主要是通過(guò)該過(guò)程驅(qū)動(dòng)的,這一過(guò)程一直持續(xù)至骨成熟。生長(zhǎng)板是機(jī)體進(jìn)行軟骨內(nèi)骨化的場(chǎng)所,主要包括五個(gè)區(qū)域,從上到下依次是靜止區(qū)、增殖區(qū)、前肥大區(qū)、肥大區(qū)和成骨區(qū)。在軟骨內(nèi)骨化的進(jìn)程中,靜止區(qū)是軟骨細(xì)胞的“補(bǔ)充池”,其具體功能存在爭(zhēng)議,但可實(shí)現(xiàn)自我更新并進(jìn)入增殖區(qū)形成排列規(guī)則、緊密且平行于骨長(zhǎng)軸的柱狀結(jié)構(gòu)的增殖型軟骨細(xì)胞,增殖型軟骨細(xì)胞進(jìn)行快速增殖以保證足夠數(shù)量的軟骨細(xì)胞進(jìn)行肥大性分化。在增殖區(qū)柱狀結(jié)構(gòu)的最下端,軟骨細(xì)胞停止增殖,進(jìn)入肥大區(qū)進(jìn)行肥大性分化,實(shí)現(xiàn)細(xì)胞體積的迅速擴(kuò)大并逐漸形成終末肥大軟骨細(xì)胞。隨后,終末肥大軟骨細(xì)胞凋亡或轉(zhuǎn)分化為成骨細(xì)胞,并形成骨嶺和骨化中心,軟骨基質(zhì)隨著血管、破骨細(xì)胞的侵入逐漸礦化,最終實(shí)現(xiàn)成骨取代軟骨基質(zhì),完成長(zhǎng)骨的縱向延伸。研究表明,在福美雙誘導(dǎo)肉雞TD和錳缺乏導(dǎo)致肉雞腿病時(shí),生長(zhǎng)板病變,生長(zhǎng)板增殖型和肥大型軟骨細(xì)胞嚴(yán)重受損。生長(zhǎng)板軟骨細(xì)胞受損會(huì)破壞軟骨內(nèi)成骨的進(jìn)程,嚴(yán)重抑制骨骼發(fā)育。軟骨細(xì)胞增殖和肥大是軟骨內(nèi)骨化的重要步驟,該過(guò)程異常會(huì)導(dǎo)致骨骼發(fā)育異常。軟骨內(nèi)骨化是由多種因子協(xié)作調(diào)控的,這些因子構(gòu)成了復(fù)雜精密的調(diào)控網(wǎng)絡(luò)確保骨骼正常的生長(zhǎng)發(fā)育。
2 骨骼發(fā)育的調(diào)控因子
在骨骼發(fā)育的過(guò)程中,多種信號(hào)通路和轉(zhuǎn)錄因子共同調(diào)控生長(zhǎng)板軟骨細(xì)胞的增殖、分化、轉(zhuǎn)分化和成骨細(xì)胞分化等。調(diào)控骨骼發(fā)育的轉(zhuǎn)錄因子主要包括性別決定基因盒9(SOX9)、Runt相關(guān)轉(zhuǎn)錄因子2(RUNX2)和肌細(xì)胞增強(qiáng)因子2C(MEF2C)等,信號(hào)分子主要包括轉(zhuǎn)化生長(zhǎng)因子β(TGF-β)、骨形態(tài)發(fā)生蛋白(主要是BMP2)、成纖維細(xì)胞生長(zhǎng)因子(FGFs)、血管內(nèi)皮生長(zhǎng)因子(VEGF)、甲狀旁腺激素相關(guān)肽(PTHrP)和印度刺猬因子(IHH)等。生長(zhǎng)板軟骨細(xì)胞外基質(zhì)(ECM)主要包括Ⅱ型膠原(COLⅡ)、X型膠原(COLX)和蛋白聚糖(ACAN)等。這些調(diào)控因子在生長(zhǎng)板各區(qū)內(nèi)特異性表達(dá)或不同區(qū)內(nèi)發(fā)揮特定調(diào)控作用(圖1),例如SOX9特異性表達(dá)于軟骨生成早期、RUNX2和MEF2C主要表達(dá)于前肥大和肥大區(qū)、PTHrP與前肥大和肥大區(qū)的受體Pth1r結(jié)合后發(fā)揮調(diào)控作用等。研究表明,福美雙誘導(dǎo)肉雞發(fā)生TD時(shí),BMP2、RUNX2、VEGF、IHH和PTHrP等多種調(diào)控因子表達(dá)異常,錳缺乏誘導(dǎo)肉雞發(fā)生TD時(shí),TGF-β、MEF2C、COLⅡ和COLX等表達(dá)異常,肉雞患細(xì)菌性軟骨壞死合并骨髓炎時(shí),SOX9、IHH和ACAN等表達(dá)異常。
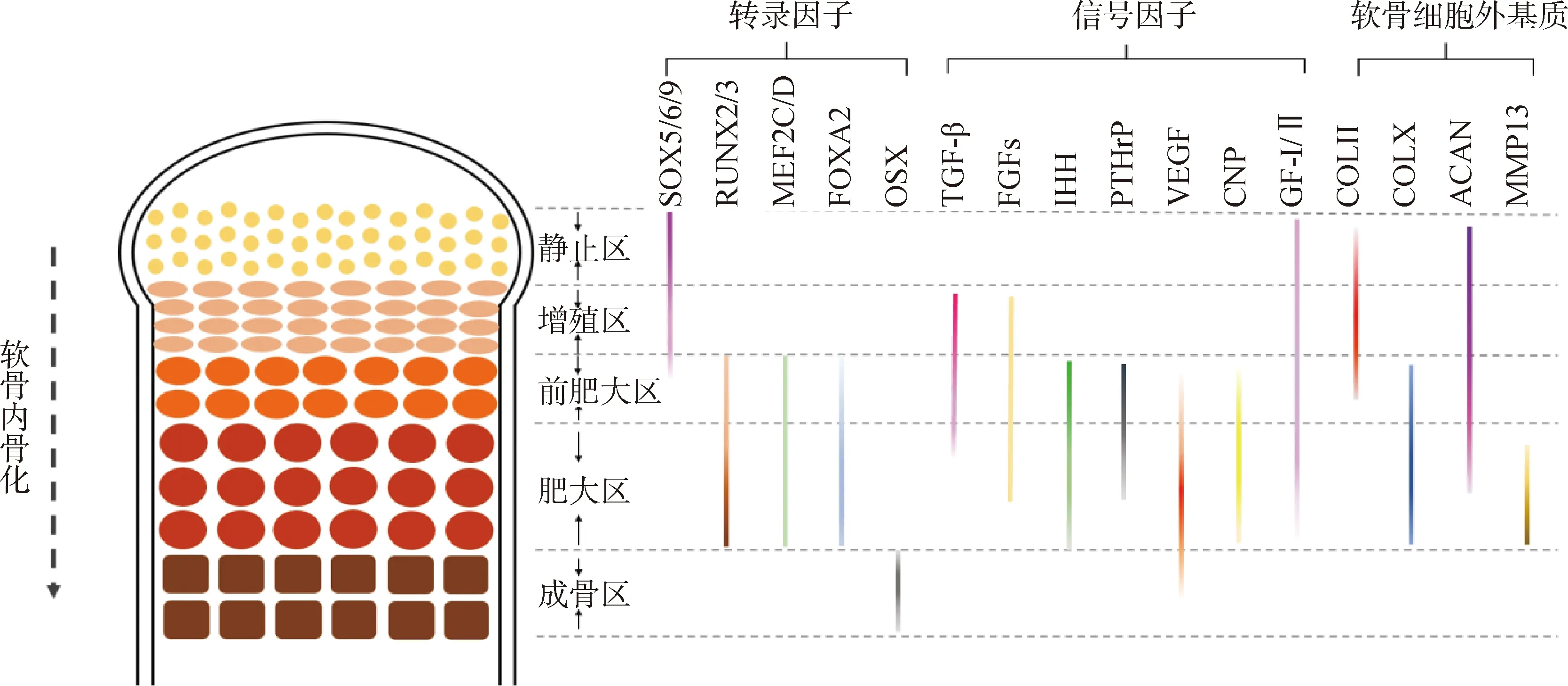
虛線箭頭表示從上到下為軟骨內(nèi)骨化的進(jìn)程(下圖同);帶顏色實(shí)線表示該因子在此區(qū)域內(nèi)表達(dá)或發(fā)揮主要調(diào)控作用,顏色越深表示在此區(qū)域表達(dá)越高或在此區(qū)域內(nèi)發(fā)揮作用越強(qiáng)Dotted arrow indicates the process of endochondral ossification from top to bottom (same as below). The colorful solid lines indicate that the factor is expressed or plays a major regulatory role in this zone. The darker the color, the higher the expression or the stronger the function of the factor in this zone圖1 各調(diào)控因子在生長(zhǎng)板各區(qū)的表達(dá)情況或在各區(qū)內(nèi)發(fā)揮主要調(diào)控作用示意圖Fig.1 Diagram of the regulatory factors are expressed in different zone or play a major role in certain zones of the growth plate
2.1 骨骼發(fā)育相關(guān)轉(zhuǎn)錄因子
2.1.1 SOX轉(zhuǎn)錄因子家族 SOX9正向調(diào)控軟骨細(xì)胞增殖,促進(jìn)早期軟骨細(xì)胞形成的標(biāo)志物(COLⅡ和ACAN)表達(dá)。SOX9是調(diào)控早期軟骨形成的主要轉(zhuǎn)錄因子,能夠調(diào)控SOX5/6表達(dá)并與其協(xié)同調(diào)控軟骨細(xì)胞增殖。9基因缺失使細(xì)胞增殖受阻,導(dǎo)致生長(zhǎng)板增殖區(qū)變窄。此外,SOX9能夠抑制RUNX2的轉(zhuǎn)錄活性,降低RUNX2泛素化并驅(qū)動(dòng)RUNX2被溶酶體降解,從而抑制軟骨細(xì)胞從增殖到前肥大的進(jìn)程,防止軟骨細(xì)胞過(guò)早肥大。SOX9還可以通過(guò)抑制β-catenin信號(hào)通路抑制成骨細(xì)胞表型的獲得,并協(xié)同MEF2C激活COLX的表達(dá)。因此,SOX9在骨形成的早期階段和軟骨細(xì)胞肥大過(guò)程中發(fā)揮復(fù)雜的調(diào)控作用,有利于維持骨骼發(fā)育及穩(wěn)態(tài)。
2.1.2 RUNX轉(zhuǎn)錄因子家族 RUNX2和RUNX3表達(dá)于前肥大和肥大型軟骨中,是調(diào)控軟骨細(xì)胞肥大及成熟的主要轉(zhuǎn)錄因子,對(duì)終末肥大軟骨細(xì)胞轉(zhuǎn)分化為成骨細(xì)胞至關(guān)重要。RUNX2調(diào)控肥大軟骨細(xì)胞中X型膠原基因(101)以及終末肥大軟骨細(xì)胞中MMP13和VEGF表達(dá)。2基因缺失抑制COLX合成、血管侵襲、軟骨細(xì)胞肥大和成熟,導(dǎo)致生長(zhǎng)板結(jié)構(gòu)破壞和骨化過(guò)程完全失敗。RUNX2誘導(dǎo)軟骨細(xì)胞表達(dá)IHH,促進(jìn)軟骨細(xì)胞的增殖,IHH也能夠誘導(dǎo)軟骨膜中RUNX2表達(dá),RUNX2還能抑制SOX9的轉(zhuǎn)錄活性。因此,RUNX2與多種調(diào)控因子協(xié)同合作共同確保骨骼正常發(fā)育。研究表明,錳缺乏可能通過(guò)抑制RUNX2和SOX9的表達(dá)抑制軟骨內(nèi)骨化的進(jìn)程進(jìn)而誘發(fā)肉雞TD。
2.1.3 MEF2轉(zhuǎn)錄因子家族 MEF2C和MEF2D表達(dá)于前肥大和肥大軟骨細(xì)胞,協(xié)同控制軟骨細(xì)胞肥大,其中MEF2C起主要調(diào)控作用,MEF2C在RUNX2的上游發(fā)揮作用,是誘導(dǎo)或維持肥大軟骨細(xì)胞表達(dá)RUNX2所必需的。在軟骨內(nèi)骨化的進(jìn)程中,MEF2C通過(guò)激活軟骨細(xì)胞肥大的基因程序來(lái)控制骨骼發(fā)育,2基因缺失抑制軟骨細(xì)胞肥大和血管侵入,導(dǎo)致骨骼發(fā)育受阻。相反,MEF2C過(guò)表達(dá)導(dǎo)致軟骨細(xì)胞過(guò)早肥大和生長(zhǎng)板過(guò)早骨化,引起侏儒癥。此外,MEF2C是101轉(zhuǎn)錄的直接調(diào)控因子。
2.1.4 FOXA轉(zhuǎn)錄因子家族 FOXA家族成員能夠調(diào)控軟骨細(xì)胞分化,與SOX9競(jìng)爭(zhēng),與RUNX和MEF2家族共同調(diào)控COLX和其他肥大軟骨標(biāo)記物的表達(dá)。小鼠軟骨細(xì)胞中缺失FOXA2和FOXA3,軟骨細(xì)胞肥大被抑制,堿性磷酸酶(ALP)表達(dá)下調(diào)和胸骨礦化缺陷,表現(xiàn)為出生后侏儒癥,生長(zhǎng)板中COLX和MMP13表達(dá)水平顯著下調(diào)。此外,敲除FOXA2能夠降低的表達(dá),而編碼,因此,F(xiàn)OXA2通過(guò)PTHrP通路調(diào)控骨骼發(fā)育。
2.1.5 成骨細(xì)胞特異性轉(zhuǎn)錄因子Osterix(OSX) OSX是成骨細(xì)胞分化和骨形成所必需的。OSX作用于RUNX2下游,是軟骨內(nèi)骨化的重要轉(zhuǎn)錄因子,在軟骨內(nèi)骨化晚期(如鈣化)發(fā)揮調(diào)控作用,OSX缺陷導(dǎo)致肥大期軟骨細(xì)胞停止軟骨內(nèi)骨化且軟骨基質(zhì)鈣化受損。此外,OSX在2基因敲除鼠中不表達(dá),且基因缺失小鼠不會(huì)發(fā)生骨形成。OSX靶向誘導(dǎo)MMP13表達(dá),OSX和RUNX2的相互作用是誘導(dǎo)軟骨內(nèi)成骨過(guò)程中MMP13表達(dá)的必要條件。研究表明,褪黑素能夠通過(guò)增強(qiáng)OSX表達(dá)直接調(diào)控晚期成骨細(xì)胞分化,促進(jìn)堿性磷酸酶活性和骨礦化。
2.2 骨骼發(fā)育相關(guān)信號(hào)分子
2.2.1 轉(zhuǎn)化生長(zhǎng)因子β(TGF-β) TGF-β在軟骨形成、軟骨細(xì)胞增殖、細(xì)胞外基質(zhì)沉積和終末分化等階段均起作用。在軟骨內(nèi)骨化的早期,TGF-β主要起正向調(diào)控作用,在分化后期起抑制作用來(lái)維持肥大軟骨細(xì)胞表型。TGF-β能夠誘導(dǎo)COLⅡ表達(dá),促進(jìn)軟骨細(xì)胞增殖,增加S期細(xì)胞比例。在分化的后期,TGF-β通過(guò)激活Smad2/3通路下調(diào)RUNX2功能,抑制軟骨細(xì)胞肥大,還能抑制Wnt8c和β-catenin通路,進(jìn)而抑制軟骨細(xì)胞成熟和COLX啟動(dòng)子的激活。研究表明,分解素和金屬蛋白酶12(ADAM12)可通過(guò)調(diào)控TGFβ1誘導(dǎo)的IGF1和RUNX2的表達(dá)參與軟骨細(xì)胞分化。
2.2.2 骨形態(tài)發(fā)生蛋白(BMPs) BMPs屬于轉(zhuǎn)化生長(zhǎng)因子β家族成員,在成骨細(xì)胞分化和骨形成中起著關(guān)鍵作用。BMP信號(hào)通路通過(guò)其受體BMPR1 (BMPR1a和BMPR1b)和BMPR2發(fā)揮作用,BMP信號(hào)通路能夠促進(jìn)IHH表達(dá),抑制FGF信號(hào)通路的關(guān)鍵效應(yīng)因子STAT和ERK1/2 MAPK的激活。BMP2通過(guò)激活Smad1/5/8信號(hào)調(diào)控多種轉(zhuǎn)錄因子的表達(dá)和功能,進(jìn)而控制骨和軟骨的發(fā)育,例如BMP-Smad信號(hào)能夠通過(guò)RUNX2和OSX通路調(diào)控骨形成。研究表明,骨碎補(bǔ)總黃酮對(duì)肉雞TD模型BMP2和RUNX2表達(dá)的下調(diào)有緩解作用。
2.2.3 成纖維細(xì)胞生長(zhǎng)因子(FGF) FGF及其受體(FGFRs)在骨形成及發(fā)育中起重要作用,敲除FGFR1和FGFR2的小鼠出生后表現(xiàn)為縱向骨生長(zhǎng)受損。FGF18能夠抑制MIR-195對(duì)軟骨細(xì)胞增殖以及對(duì)COLⅡ和ACAN表達(dá)的抑制作用,F(xiàn)GF2能夠誘導(dǎo)MMP13表達(dá)。FGFR1和FGFR2失活使FGF9表達(dá)代償性增加,進(jìn)而異常激活FGFR3,抑制軟骨細(xì)胞增殖和肥大,而FGFR3信號(hào)通路的增強(qiáng)又進(jìn)一步促進(jìn)FGF9和FGF18表達(dá),抑制IHH和Pthlh表達(dá)。FGFR3表達(dá)和信號(hào)傳導(dǎo)的異常激活可損害生長(zhǎng)板軟骨細(xì)胞增殖、終末分化和細(xì)胞外基質(zhì)的合成。此外,F(xiàn)GF還通過(guò)調(diào)控軟骨細(xì)胞自噬來(lái)調(diào)節(jié)生長(zhǎng)板ECM的組成。
2.2.4 印度刺猬蛋白(IHH) IHH主要表達(dá)于前肥大區(qū)和肥大區(qū),是骨骼發(fā)育和穩(wěn)態(tài)的關(guān)鍵調(diào)控因子。IHH能夠促進(jìn)軟骨細(xì)胞增殖和成骨過(guò)程,也通過(guò)刺激PTHrP分泌調(diào)控肥大軟骨細(xì)胞的分化。IHH能夠調(diào)控軟骨膜中RUNX2的表達(dá),下調(diào)IHH導(dǎo)致ALP活性減低和軟骨基質(zhì)礦化減少。基因缺失小鼠表現(xiàn)為增殖型軟骨細(xì)胞顯著減少,軟骨細(xì)胞成熟異常以及缺乏成熟的成骨細(xì)胞。Deng等研究表明,敲除小鼠軟骨細(xì)胞基因可能通過(guò)影響TGF-β/Smad和OPG/RANKL信號(hào)通路抑制軟骨細(xì)胞生長(zhǎng)和分化。
2.2.5 甲狀旁腺激素相關(guān)肽(PTHrP) PTHrP在骨發(fā)育和成熟過(guò)程中發(fā)揮作用,通過(guò)與PTH1R受體結(jié)合來(lái)抑制軟骨細(xì)胞終末分化。敲除基因?qū)е萝浌莾?nèi)成骨嚴(yán)重異常,基因缺失仔鼠的四肢骨明顯縮短,生長(zhǎng)板顯著變窄。研究表明,通過(guò)添加環(huán)巴胺抑制PTHrP上游的IHH信號(hào)后,基因表達(dá)水平隨時(shí)間的延長(zhǎng)顯著降低。PTHrP的表達(dá)受到其上游信號(hào)因子IHH影響,而過(guò)表達(dá)PTHrP又能下調(diào)IHH表達(dá),形成IHH/PTHrP負(fù)反饋環(huán)調(diào)控軟骨細(xì)胞增殖和分化,保證軟骨內(nèi)骨化有條不紊地進(jìn)行。
2.2.6 C型利鈉肽(CNP) CNP是維持軟骨穩(wěn)態(tài)和軟骨內(nèi)成骨的重要調(diào)節(jié)因子,CNP與其主要受體利鈉肽受體B(NPRB)結(jié)合后,刺激軟骨細(xì)胞和軟骨基質(zhì)的合成。Fujii等用ZFN技術(shù)成功建立CNP敲除大鼠模型,CNP 敲除大鼠表現(xiàn)為骨骼生長(zhǎng)受損,長(zhǎng)骨較野生型大鼠短,生長(zhǎng)板較野生型大鼠窄,特別是肥大區(qū)明顯變窄。研究表明,CNP通過(guò)抑制絲裂原激活蛋白激酶(MAPK)通路來(lái)拮抗FGFR3下游信號(hào)通路。然而,有關(guān)CNP在肉雞發(fā)生腿病時(shí)表達(dá)的研究較少,需要進(jìn)一步探索。
2.2.7 WNTs信號(hào)因子 WNTs信號(hào)因子對(duì)于骨骼形成、維持骨穩(wěn)態(tài)和進(jìn)行骨修復(fù)至關(guān)重要。在早期骨骼發(fā)育的過(guò)程中,Wnt/β-catenin信號(hào)通路激活OSX表達(dá),誘導(dǎo)間充質(zhì)干細(xì)胞向成骨細(xì)胞分化。隨后,生長(zhǎng)板軟骨細(xì)胞中的WNTs信號(hào)因子促進(jìn)軟骨細(xì)胞存活并調(diào)控軟骨內(nèi)骨化進(jìn)程。WNTs信號(hào)因子能夠促進(jìn)BMP和IHH表達(dá),促進(jìn)軟骨細(xì)胞肥大和調(diào)控骨礦化。此外,WNT11能夠上調(diào)RUNX2表達(dá),并與TGF-β協(xié)同促進(jìn)軟骨細(xì)胞分化。研究表明,在福美雙誘導(dǎo)肉雞TD模型中,WNT4表達(dá)下調(diào),使用淫羊藿苷通過(guò)上調(diào)WNT4表達(dá)能夠改善TD的發(fā)生。
2.2.8 血管內(nèi)皮生長(zhǎng)因子(VEGF) 在軟骨內(nèi)骨化的進(jìn)程中,血管內(nèi)皮生長(zhǎng)因子A(VEGFA)由前肥大和肥大軟骨細(xì)胞分泌,對(duì)于維持軟骨內(nèi)骨化中血管生成和侵入以及成骨細(xì)胞分化至關(guān)重要。VEGFA能夠誘導(dǎo)內(nèi)皮細(xì)胞遷移和增殖,間接刺激成骨和血管生成,并通過(guò)旁分泌信號(hào)調(diào)節(jié)成骨生長(zhǎng)因子的分泌。VEGFA缺失導(dǎo)致胚胎骨形成受阻,基因敲除小鼠的生長(zhǎng)板肥大區(qū)變寬、無(wú)血管侵入并出現(xiàn)細(xì)胞的大面積死亡。VEGFA能夠刺激IHH的表達(dá)和活性,增加β-catenin表達(dá)。研究表明,在福美雙誘導(dǎo)的TD模型中,VEGFA表達(dá)顯著下調(diào),生長(zhǎng)板血管形成和侵入延遲。
2.2.9 缺氧誘導(dǎo)因子(HIF-1α) 生長(zhǎng)板軟骨細(xì)胞形成的早期階段是處于缺氧狀態(tài)的,局部激活HIF-1α有利于保證軟骨細(xì)胞生存并調(diào)控軟骨細(xì)胞膠原合成和修飾,延長(zhǎng)HIF-1α信號(hào)能夠干擾細(xì)胞生物能學(xué)和生物合成,進(jìn)而導(dǎo)致骨骼發(fā)育不良。在缺氧條件下,缺失HIF-1α的軟骨細(xì)胞表現(xiàn)為Ⅱ型膠原表達(dá)顯著下調(diào)、能量生成不足和軟骨細(xì)胞增殖遲緩。HIF-1α和BMP2協(xié)同促進(jìn)軟骨細(xì)胞的增殖、抑制軟骨細(xì)胞肥大并維持軟骨細(xì)胞表型。此外,HIF-1α能夠正向調(diào)控VEGF的表達(dá)。
2.2.10 胰島素樣生長(zhǎng)因子(IGF) 軟骨內(nèi)成骨受多種激素和分泌因子調(diào)節(jié),其中GH-IGF軸對(duì)于骨骼發(fā)育至關(guān)重要。IGF-I和IGF-Ⅱ是生長(zhǎng)和代謝的關(guān)鍵調(diào)控因子,IGF-I表達(dá)于生長(zhǎng)板各個(gè)區(qū)域,但表達(dá)程度低,而IGF-Ⅱ在生長(zhǎng)板中高度表達(dá),從靜止區(qū)到肥大區(qū)表達(dá)水平逐漸降低。研究表明,IGF-I可增強(qiáng)BMP在體外和體內(nèi)成骨作用,添加ICF-1有助于提高大鼠異位骨模型的骨礦化。然而,有關(guān)IGF-Ⅱ?qū)ιL(zhǎng)板維持正常生理機(jī)能的作用以及對(duì)骨骼發(fā)育的調(diào)控作用的研究報(bào)道較少,需要進(jìn)一步探究。
2.2.11 微RNA(miRNA) 多種miRNA均有保證骨骼發(fā)育的作用,能夠調(diào)控軟骨細(xì)胞增殖和維持軟骨表型。例如,miR-218靶向并下調(diào)了多種調(diào)控因子(RUNX2、MEF2C和COLⅩ)的表達(dá),進(jìn)而調(diào)控軟骨細(xì)胞的終末肥大。miR-21通過(guò)調(diào)控MEF2C、RUNX2和OSX等來(lái)調(diào)控成骨細(xì)胞發(fā)育和骨礦化。miR-195負(fù)向調(diào)控FGF18,通過(guò)ERK1/2途徑調(diào)控軟骨細(xì)胞增殖,通過(guò)SOX9途徑調(diào)控Ⅱ型膠原及蛋白聚糖的表達(dá),從而起到維持軟骨表型的作用,下調(diào)miR-195可通過(guò)靶向FGF-18途徑促進(jìn)軟骨細(xì)胞增殖和維持軟骨細(xì)胞表型。
2.3 細(xì)胞外基質(zhì)(ECM)
ECM起支持和保護(hù)軟骨細(xì)胞的作用,同時(shí)作為一種媒介使信號(hào)分子和生長(zhǎng)因子能夠通過(guò)無(wú)血管的軟骨向靶細(xì)胞擴(kuò)散。軟骨細(xì)胞ECM主要由Ⅱ型膠原(COLⅡ)、X型膠原(COLX)、蛋白聚糖(ACAN)組成,并富含基質(zhì)金屬蛋白酶(主要是MMP13)。內(nèi)骨化的早期階段,軟骨細(xì)胞特異性合成COLⅡ和ACAN,COLⅡ是早期軟骨生成的特異性標(biāo)志物。COLX主要表達(dá)于肥大區(qū),是軟骨細(xì)胞肥大性分化的特異性標(biāo)志物。研究表明,Ⅱ型膠原基因2a1缺失的仔豬表現(xiàn)為長(zhǎng)骨明顯縮短。此外,COLⅡ能夠促進(jìn)FOXA2表達(dá),ACAN與FOXA2表達(dá)呈正相關(guān)。MMP13主要由終末肥大軟骨細(xì)胞分泌,起降解軟骨基質(zhì)的作用,小鼠生長(zhǎng)板13基因缺失使間質(zhì)膠原大量聚集,導(dǎo)致肥大區(qū)明顯變寬,并延遲軟骨內(nèi)成骨進(jìn)程、初級(jí)骨化中心的形成和血管侵入。
上述調(diào)控因子構(gòu)成了復(fù)雜精密的網(wǎng)絡(luò)調(diào)控生長(zhǎng)板軟骨內(nèi)骨化(圖2、圖3),其中SOX9、RUNX2、IHH和PTHrP等主要調(diào)控生長(zhǎng)板軟骨細(xì)胞的增殖和分化, RUNX2、OSX和VEGF等主要調(diào)控骨形成后期階段的成骨細(xì)胞分化、骨礦化和血管侵入等,各調(diào)控因子獨(dú)自調(diào)控或不同調(diào)控因子間相互作用協(xié)同調(diào)控軟骨內(nèi)骨化的進(jìn)程以完成骨骼正常的生長(zhǎng)發(fā)育。
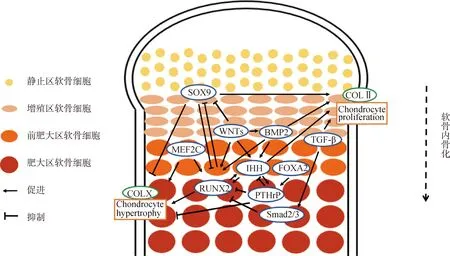
圖2 軟骨內(nèi)骨化過(guò)程中調(diào)控軟骨細(xì)胞增殖和分化的分子機(jī)制圖Fig.2 Diagram of molecular mechanism that regulate chondrocyte proliferation and differentiation during endochondral ossification
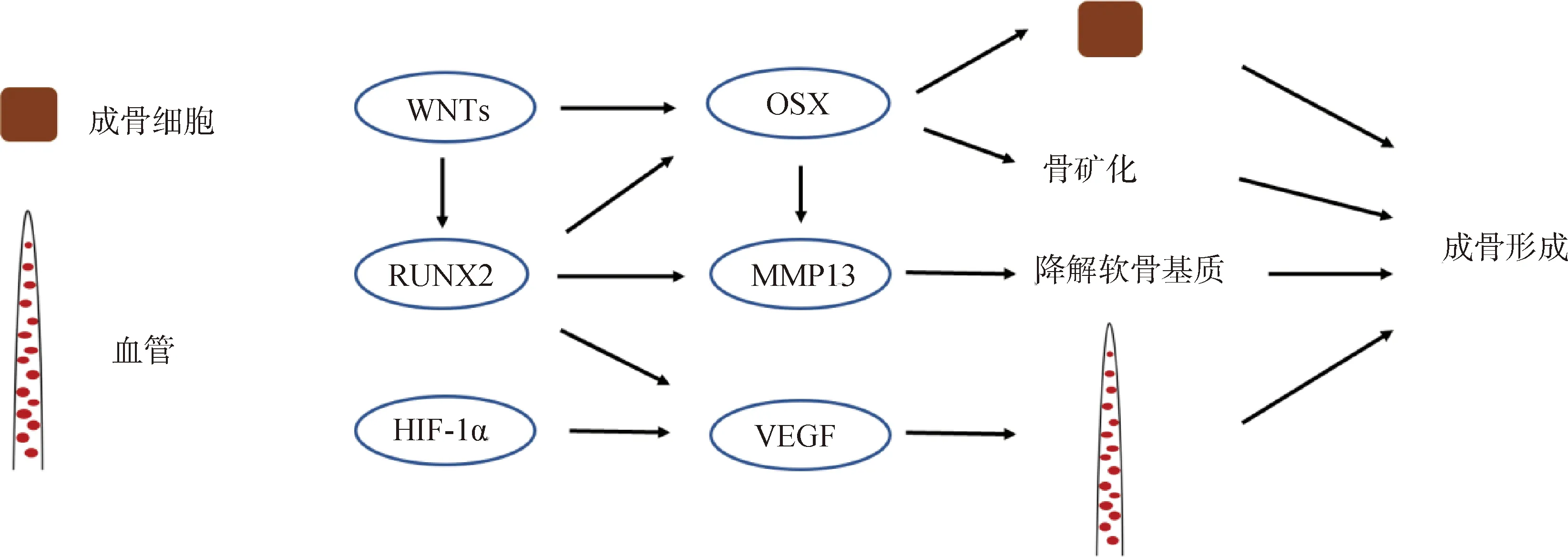
圖3 軟骨內(nèi)骨化過(guò)程中調(diào)控成骨形成的分子機(jī)制圖Fig.3 Diagram of molecular mechanism that regulate osteogenesis during endochondral ossification
3 小 結(jié)
骨骼發(fā)育主要是通過(guò)由SOX9、RUNX2、MEF2C、TGF-β、BMP2、FGFs、IHH和PTHrP等多種因子組成的復(fù)雜精確的調(diào)控網(wǎng)絡(luò)調(diào)控的軟骨內(nèi)骨化完成的。持續(xù)研究調(diào)控骨骼發(fā)育的分子機(jī)制,特別是進(jìn)一步探究各調(diào)控因子的局部調(diào)控作用及不同調(diào)控因子間的相互作用,對(duì)于充分理解機(jī)體如何精確調(diào)控骨形成和生長(zhǎng)板的軟骨內(nèi)骨化至關(guān)重要,有助于加快對(duì)肉雞腿病發(fā)病的分子機(jī)制的探究,為開(kāi)發(fā)新的治療靶點(diǎn)和方法提供可能,通過(guò)調(diào)控軟骨內(nèi)成骨來(lái)逆轉(zhuǎn)各種骨骼疾病。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
