基于生物信息學分析人微小RNA-142-5p的靶基因及其與心血管疾病的關聯
2022-05-14 08:03:38俞虹宇蔣智淵
廣西醫學 2022年4期
俞虹宇 周 爽 蔣智淵
(廣西醫科大學第一附屬醫院心血管內科,南寧市 530021,電子郵箱:1606188485@qq.com)
miRNA是一類長度約22個核苷酸的非編碼RNA,其主要作用是通過與靶基因mRNA互補堿基配對,RNA誘導沉默復合體(RNA-induced silencing complex,RISC)降解mRNA或阻礙其翻譯,從而在轉錄后水平負性調控靶基因的表達。真菌、植物和動物中的miRNA堿基序列在物種進化過程中相當保守,且表達具備時序性和組織特異性[1]。由此可見,miRNA在物種進化過程中起著不可替代的作用,決定著組織和細胞的功能特異性。研究表明,miRNA參與許多生物學過程,包括細胞的生長、增殖、分化、凋亡,器官的形成,生物個體的發育、繁殖,并與人類疾病息息相關[1]。
微小RNA-142(microRNA-142,miR-142)以兩種成熟異構體的形式存在,分別為miR-142-3p和miR-142-5p,它們是由完整miR-142雙鏈的正義鏈和反義鏈經核糖核酸酶代謝產生。目前,針對miR-142-5p在心血管疾病中的作用研究,主要集中于心肌凋亡方面[2-4],其在心血管疾病中的作用仍有待探索。本研究利用生物信息學技術預測人miR-142-5p(homo sapiens-miR-142-5p,hsa-miR-142-5p)的靶基因,并通過分析靶基因涉及的生物學過程和信號通路初步探討hsa-miR-142-5p在心血管疾病中的作用。
1 資料與方法
1.1 檢索hsa-miR-142-5p在各個組織器官中的表達情況 使用miRGator v3.0數據庫(http://genome.ewha.ac.kr/miRGator/miRGator.html)對hsa-miR-142-5p進行檢索,查看其在各個組織器官中的表達情況。
1.2 分析hsa-miR-142-5p的染色體定位及物種保守性 使用PubMed數據庫(http://www.ncbi.nlm.nih.gov/pubmed/)檢索hsa-miR-142-5p的相關文獻,并結合miRBase 22.1(http://www.mirtarbase.org)和UCSC Genome Browser(http://genome.ucsc.edu/)的檢索結果,獲取其染色體定位、堿基序列和物種保守性等基本信息。
1.3 預測hsa-miR-142-5p的靶基因 利用TargetScan 7.2(http://www.targetscan.org/vert_72)、DIANA TOOLS(http://www.microrna.gr/microT-CDS)、miRDB(http://www.mirdb.org/)和miRWalk 3.0(http://www.umm.uni-heidel-berg.de/apps/zmf/mirwalk/)預測hsa-miR-142-5p的靶基因,并利用Venny.2.1.0建立Venn圖,取它們的交集。通過miRTarBase(http://mirtarbase.mbc.nctu.edu.tw)獲取經過較強實驗證據驗證的hsa-miR-142-5p的靶基因,將其與上述靶基因的交集進行合并,組成新的基因集。
1.4 hsa-miR-142-5p靶基因集的生物功能和通路富集分析 使用基因本體論(Gene Ontology,GO)功能富集分析網站(http://geneontology.org/)和 DAVID 6.7數據庫(https://david.ncifcrf.gov/)對此基因集進行功能富集分析,并利用京都基因與基因組百科全書(Kyoto Encyclopedia of Genes and Genomes,KEGG)數據庫(http://www.genome.jp/kegg/)進行信號通路富集分析。選取高頻率注釋及有統計學意義的信號通路進行分析,其中以P<0.05為差異有統計學意義。
2 結 果
2.1 hsa-miR-142-5p的表達情況 hsa-miR-142-5p在淋巴細胞、淋巴樣細胞、外周血單核細胞、扁桃體中的表達豐度較高,其中心臟存在基礎表達量。見圖1。
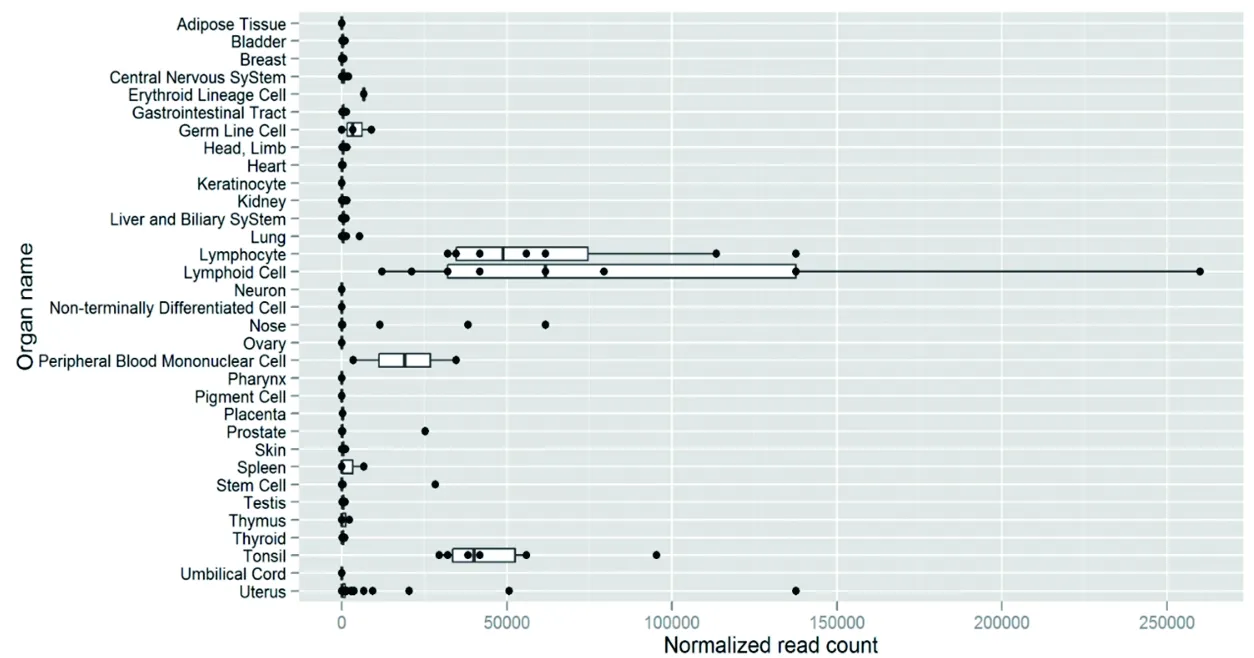
圖1 hsa-miR-142-5p在各組織器官中的表達情況
2.2 hsa-miR-142-5p的染色體定位及物種保守性 使用UCSC Genome Browser檢索,發現hsa-miR-142-5p定位于染色體17q22,共21個堿基序列。使用miRBase檢索到包括人類在內的共26個不同物種miR-142-5p的成熟序列,對其進行分析后發現,在這26個物種中幾乎都包含了miR-142-5p的“CAUAAAGUAGAAAGCACU”18個堿基序列,表明該堿基序列在這26個物種中高度保守,見表1。

表1 26個物種 miR-142-5p 的成熟序列
2.3 hsa-miR-142-5p的靶基因預測結果 TargetScan、DIANA TOOLS、miRDB和miRWalk軟件預測得到的 hsa-miR-142-5p靶基因分別為950個、1 934個、1 133個、956個,將4種預測方法所得的靶基因取交集后共獲得39個靶基因,分別為AFF4、AHR、AKAP11、ARMC8、ATP13A3、ATP1B1、BMPR1A、BTBD7、C21orf91、CHD9、CIT、CNTN1、EIF4E3、FAM91A1、FBXO21、FIGN、GAS7、GRSF1、HS2ST1、IL6ST、ITGAV、KITLG、MAN1A1、NFAT5、NR2C1、ONECUT2、OTUD4、PIK3CA、PUM2、RAP1A、RC3H1、SCN2A、SETX、SLC5A3、SPRY3、SPTBN1、TBC1D9、TRIM36、ZBTB37,見圖2。通過miRTarBase獲得12個經較強實驗證據驗證過的靶基因,分別是CLDN1、HIF1A、IGF2BP3、NFE2L2、PTEN、RAC1、SIRT1、SMAD3、SOCS1、TGFBR2、TP53INP1、ZEB1,合并后共獲得51個候選基因,組成1個靶基因集。

圖2 4種軟件對has-miR-142-5p靶基因預測結果的Venn圖
2.4 hsa-miR-142-5p靶基因集的GO功能富集分析和KEGG通路富集分析結果 將hsa-miR-142-5p靶基因集進行GO功能富集分析,分別獲得66項生物學過程、18項分子功能和13個細胞組分,其中有統計學差異的生物學過程有37項、分子功能9項、細胞組分7項。結果顯示,靶基因功能富集在心內膜墊融合、胚胎心管形態發生、心臟心室形態的發生和心肌肥厚的調控、心臟上皮向間質轉化的正調控等生物學過程,富集在轉錄因子β受體、蛋白激酶和特異性核酸序列等分子功能上,以及胞質溶膠、核染色質(體)和PML主體等細胞組分中(均P<0.05)。見表2。
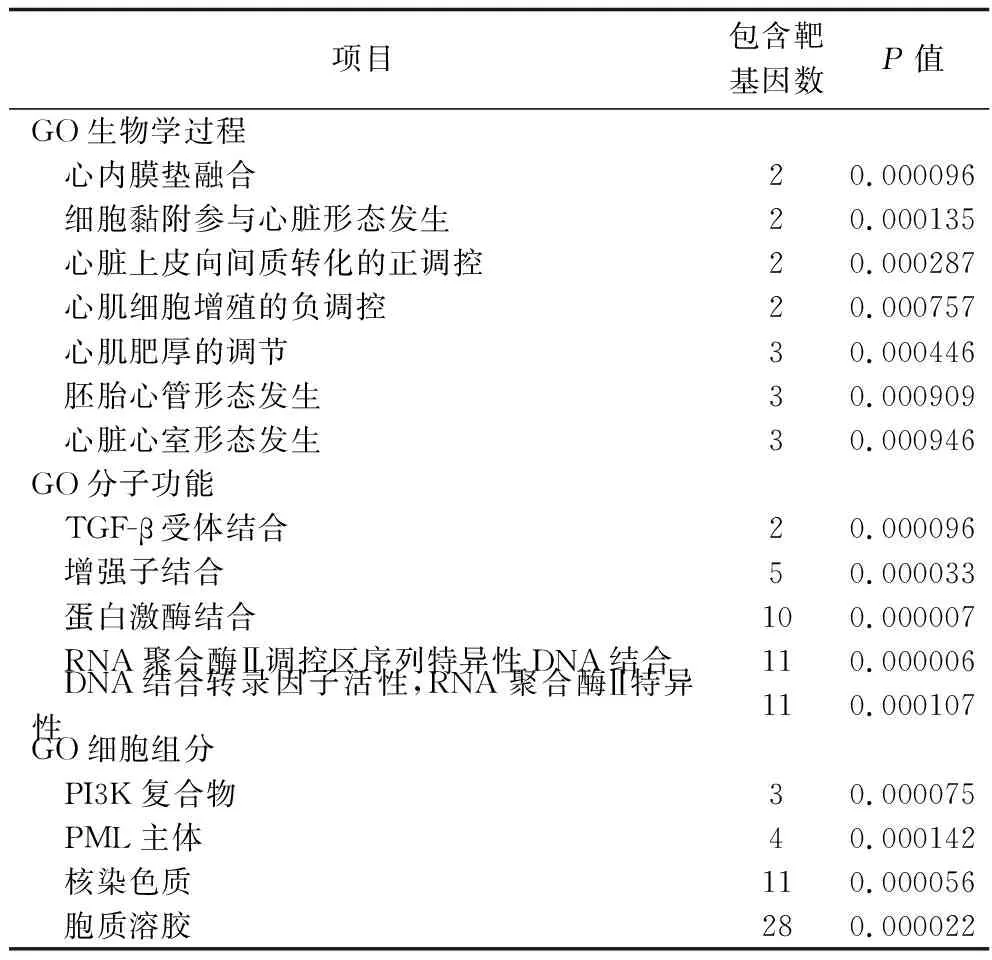
表2 hsa-miR-142-5p靶基因的 GO分析結果
通過DAVID數據庫對 hsa-miR-142-5p靶基因集進行基于KEGG的信號通路富集分析,共獲取24條信號通路(P<0.05),本部分僅選取其中8個信號通路進行分析,包括hsa-miR-142-5p靶基因富集于局灶性粘連、磷脂酰肌醇3-激酶(phosphoinositide 3-kinases,PI3K)/蛋白激酶B(protein kinase B,Akt)信號通路、叉頭框蛋白O(forkhead box O,FoxO)信號通路、白細胞跨內皮遷移、甲狀腺激素信號通路、Rap1信號通路、黏附連接、轉化生長因子β(transforming growth factor β,TGF-β)等信號通路中(均P<0.05),見表3。
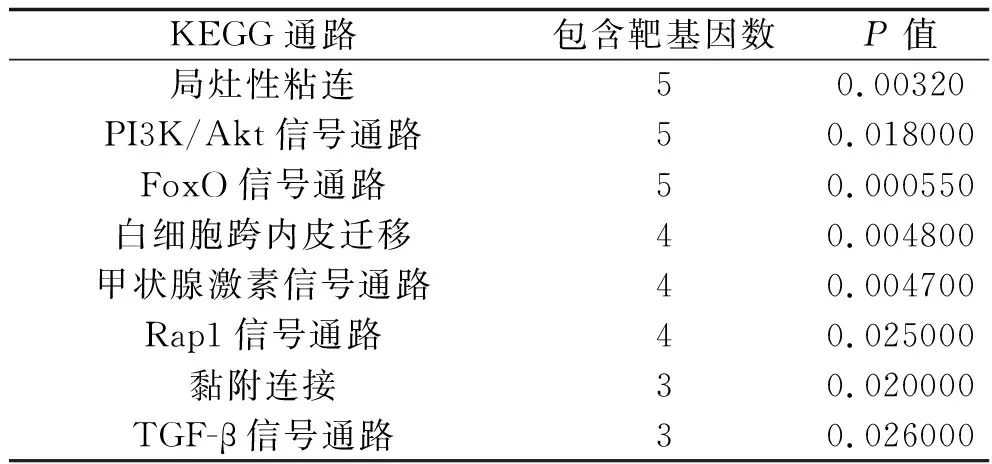
表3 hsa-miR-142-5p靶基因的KEGG通路分析結果
3 討 論
hsa-miR-142-5p序列在人類參考基因組中被發現,是miR-142的家族成員之一,經檢索發現其序列共有21個核苷酸,且在26個物種間高度保守。通過5個數據庫進行靶基因預測,共獲得包含51個候選基因的靶基因集;GO功能富集分析結果顯示,hsa-miR-142-5p的靶基因功能富集在心內膜墊融合、胚胎心管形態發生、心臟心室形態的發生和心肌肥厚的調控、心臟上皮向間質轉化的正調控等生物學過程,并主要集中于胞質溶膠、染色質(體)等細胞組分,調控轉錄生長因子β受體、蛋白激酶和特異性核酸序列等分子功能。由此可見,hsa-miR-142-5p與多個基因相互作用,調節生命形式所必需的基本功能。
研究表明,擴張型心肌病患兒的外周血miR-142-5p表達顯著上調,miR-142-5p有可能成為診斷擴張型心肌病和評估心力衰竭的潛在生物指標[5]。還有研究表明, miR-142-5p表達下調與非ST段抬高型的急性冠脈綜合征患者慢性心力衰竭的發生有關[6]。此外,母體血清中miR-142-5p表達下調與胎兒先天性心臟缺陷的發生相關[7]。Pan等[8]研究發現,miR-142-5p的循環水平在支架內在狹窄患者中顯著升高。上述研究提示miR-142-5p或可作為診斷心血管疾病的血清學標志物,但其在上述疾病發生中的具體作用仍有待探索。近期,已有學者對miR-142-5p在心血管疾病發生中的作用進行了探索。Du等[2]研究發現,長鏈非編碼RNA GAS5可作為競爭性內源RNA與miR-142-5p結合,沉默長鏈非編碼RNA GAS5表達可促進miR-142-5p表達,從而激活PI3K/Akt和絲裂原活化蛋白激酶激酶/細胞外調節蛋白激酶信號通路,減少缺氧造成的大鼠心肌凋亡。Wang等[9]的研究表明, miR-142-5p與miR-212-5p協同抑制c-Myc/腫瘤蛋白p53誘導核蛋白1信號通路,從而抑制小鼠心肌梗死后心臟成纖維細胞增生和膠原纖維分泌,改善心臟功能。Shrestha等[4]發現,miR-142-5p可以控制出生后心肌細胞的適應性生長,是調控心肌細胞適應性肥厚的關鍵因子;miR-142-5p表達上調可導致小鼠心肌細胞過度凋亡、心臟明顯擴張和心功能障礙。此外,還有研究表明,miR-142-5p還能通過下調B細胞易位基因3促進血管平滑肌細胞增殖[10]。
上述研究多集中在miR-142-5p心肌細胞凋亡中的作用,個別研究涉及心肌纖維化及平滑肌細胞增殖。我們利用生物信息學技術進行分析后發現,hsa-miR-142-5p與心內膜墊融合、胚胎心管形態發生、心臟心室形態發生和心肌肥厚的調節有關,提示hsa-miR-142-5p可能在先天性心臟病和心肌肥厚的發生中起著重要作用,此外,hsa-miR-142-5p還參與了心臟上皮向間質轉化的調控,這可能與心肌纖維化有關[11]。我們還發現,hsa-miR-142-5p參與PI3K/Akt信號通路、黏附連接、TGF-β信號通路、甲狀腺激素信號通路、Rap1信號通路和FoxO信號通路的調控。研究表明,PI3K/Akt信號通路與心肌纖維化和心肌肥厚有關[12-13],黏附連接信號通路對于維持血管內皮細胞屏障及正常功能起著重要作用[14],TGF-β信號通路與心肌纖維化密切相關[15],甲狀腺激素信號通路與心肌肥厚有關[16],Rap1信號通路則主要與血管平滑肌細胞及血管內皮細胞的調控有關[17-18],而FoxO信號通路則主要與心肌肥厚的調控有關[19]。上述結果提示,hsa-miR-142-5p在先天性心臟病、心肌肥厚、心肌纖維化、血管平滑肌細胞和血管內皮細胞調控中具有一定的作用,可作為今后進一步研究的方向,而PI3K/Akt信號通路、黏附連接、TGF-β信號通路、甲狀腺激素信號通路、Rap1信號通路和FoxO信號通路則為進一步探索hsa-miR-142-5p在上述病理生理過程中的具體機制提供了線索。
綜上所述,hsa-miR-142-5p的靶基因可能與先天性心臟病、心肌肥厚、心肌纖維化、血管平滑肌細胞和血管內皮細胞調控有關,而PI3K/Akt信號通路、黏附連接、TGF-β信號通路、甲狀腺激素信號通路、Rap1信號通路和FoxO信號通路可能參與上述病理生理過程。上述結果為進一步研究hsa-miR-142-5p在心血管疾病發生中的作用及機制提供了一定的線索。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
鴨綠江(2021年35期)2021-04-19 12:24:18
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
電子制作(2018年18期)2018-11-14 01:48:24
汽車觀察(2018年10期)2018-11-06 07:05:26
電子制作(2018年11期)2018-08-04 03:25:42
山東工業技術(2016年15期)2016-12-01 05:31:22
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
