lncRNA SNHG14靶向miR-149-5p調控MIA誘導的骨關節炎模型細胞損傷
2022-03-22 12:07:54黃煒賀媛張衛濤
河北醫藥 2022年5期
黃煒 賀媛 張衛濤
骨關節炎(osteoarthritis,OA)是最常見的慢性退行性關節疾病,是由關節軟骨破壞、軟骨下骨改變或滑膜積液引起的,引起關節疼痛和僵硬[1,2]。骨關節炎的流行病學非常復雜,其分子機制仍未完全了解。軟骨細胞是維持軟骨結構和功能的重要細胞結構,其死亡會破壞胞外基質合成與降解的平衡,從而進一步加劇OA[3]。長鏈非編碼RNA(lncRNAs)是指長度超過200個核苷酸且不具備蛋白質編碼能力的RNA分子,已被證實在炎癥相關疾病的發生發展中發揮重要作用[4]。研究發現,在MPP誘導的帕金森細胞模型中,lncRNA SNHG14表達上調,下調lncRNA SNHG14通過miR-214-3p/KLF4軸增強細胞活力、抑制細胞凋亡和促炎因子的產生來減輕細胞損傷[5]。在OGD/R誘導的HT22小鼠海馬神經元細胞損傷中,lncRNA SNHG14通過miR-182-5p/BINP3軸誘導HT22細胞過度自噬,從而加劇神經元損傷[6]。然而lncRNA SNHG14在骨關節炎中的作用尚不清楚。本實驗通過StarBase預測發現lncRNA SNHG14與miR-149-5p具有結合位點。研究報道,下調miR-149逆轉了抑制PVT1表達對IL-1誘導的軟骨細胞異常代謝功能障礙和炎癥的影響[7]。盡管已有研究確定miR-149在骨關節炎中的作用,但lncRNA SNHG14在骨關節炎中的作用及其機制是否與miR-149-5p有關目前還尚未可知。因此,本實驗旨在研究lncRNA SNHG14是否通過靶向調控miR-149-5p表達來影響膿毒癥炎性反應。
1 材料與方法
1.1 材料與試劑 雄性SD大鼠,購自上海斯萊克實驗動物有限公司;碘乙酸納(MIA)購自美國Sigma公司;DMEM培養液購自上海鈺森生物技術有限公司;胎牛血清購自杭州亦博生物科技有限公司;Annexin V-FITC/PI凋亡檢測試劑盒購自上海晶抗生物工程有限公司;RIPA蛋白裂解液、BCA試劑盒購自上海研謹生物科技有限公司;MDA、SOD、CAT檢測試劑盒購自南京建城生物工程研究所;Trizol試劑、反轉錄試劑盒、熒光定量試劑盒購自日本Takara公司;Lipofectamine2000購自上海信帆生物科技有限公司。
1.2 大鼠軟骨細胞制備 無菌環境下,小心刮取大鼠雙側膝關節脛骨坪及髕骨內側透明軟骨,充分漂洗數次,剪碎至1 mm3,依次用0.25%胰蛋白酶和0.2%膠原酶消化,得到單細胞懸液,用DMEM/F-12培養基重懸細胞,培養于37℃ 和 5% CO2培養箱中,取2~3代對數生長期細胞用于后續實驗。
1.3 MIA處理與分組 將分離培養的大鼠軟骨細胞記為對照組,將經4 μmol/L MIA 處理 24 h 的大鼠軟骨細胞記為MIA組,將si-NC、si-SNHG14分別轉染至MIA組中,記為MIA+si-NC組和MIA+si-SNHG14組;將miR-NC、miR-149-5p分別轉染至MIA組中,記為MIA+miR-NC組和MIA+miR-149-5p組;將si-SNHG14分別與anti-miR-NC、anti-miR-149-5p共轉染至MIA組中,記為si-SNHG14+anti-miR-NC組和si-SNHG14+anti-miR-149-5p組。
1.4 實時熒光定量PCR(RT-qPCR) 提取各組細胞總RNA,反轉錄成cDNA,按試劑盒說明進行PCR,檢測各組lncRNA SNHG14和miR-149-5p水平以驗證轉染效果。lncRNA SNHG14上游引物序列:5’-GGGTGTTTACGTAGACCAGAACC-3’,下游引物序列:5’-CTTCCAAAAGCCTTCTGCCTTAG-3’;miR-149-5p上游引物序列:5’-CCCTCATTCTGT GCCACACTCCAGCTGGG-3’,下游引物序列:5’-TGGTGTCGTGGAGTCG-3’;GAPDH上游引物序列:5’-AATTCCATGGCACCGTCAAG-3’,下游引物序列:5’-TGGACTCCACGACGTACTC-3’;U6上游引物序列:5’-GCTTCGGCAGCACATATACTAAAAT-3’,下游引物序列:5’-CGCTTCACGAATTTGCGTGTCAT-3’;引物由上海生工生物工程公司合成。
1.5 Annexin V檢測細胞凋亡 按照Annexin V-FITC/PI試劑盒說明檢測SW480細胞凋亡,收集細胞并用PBS漂洗2次,結合緩沖液重懸后,加入Annexin V-FITC、碘化丙啶(PI)各5 μl,37℃孵育15 min后,用流式細胞儀檢測細胞凋亡。
1.6 蛋白質印跡(Western blot)法檢測蛋白表達 提取各組細胞總蛋白,用BCA試劑盒進行定量。各組蛋白上樣量60 μg,進行SDS-PAGE后轉移至PVDF上,用5%脫脂牛奶室溫封閉,然后加入一抗4℃孵育過夜,PBS洗滌3次,加入二抗室溫孵育2 h,PBS洗滌3次,暗室中曝光顯影,定影,用Quantity One軟件檢測各組蛋白條帶灰度值,以目的條帶和GAPDH條帶的比值作為蛋白表達水平。
1.7 MDA水平與SOD和CAT活性測定 按照MDA、SOD、CAT檢測試劑盒說明書檢測各組軟骨細胞中MDA水平與SOD和CAT活性。
1.8 雙熒光素酶報告實驗檢測lncRNA SNHG14和miR-149-5p的靶向關系 構建SNHG14野生型和突變型熒光素酶表達載體WT-SNHG14和MUT-SNHG14,將其分別與miR-NC和miR-149-5p共轉染至MIA組軟骨細胞中,按照說明書檢測熒光素酶活性。

2 結果
2.1 lncRNA SNHG14和miR-149-5p在MIA誘導軟骨細胞損傷中的表達 與對照組比較,經MIA誘導的軟骨細胞中,SNHG14的表達水平升高,miR-149-5p的表達水平降低(P<0.05)。見表1。

表1 lncRNA SNHG14和miR-149-5p在MIA誘導軟骨細胞損傷中的表達
2.2 抑制lncRNA SNHG14表達對MIA誘導軟骨細胞損傷的影響 與對照組比較,MIA組SNHG14的表達水平升高,細胞凋亡率升高,Bcl-2表達水平降低,Bax表達水平升高,MDA表達水平升高,SOD和CAT活性降低(P<0.05);與MIA+si-NC組比較,MIA+si-SNHG14組SNHG14的表達水平降低,細胞凋亡率降低,Bcl-2表達水平升高,Bax表達水平降低,MDA表達水平降低,SOD和CAT活性升高(P<0.05)。見圖1,表2。
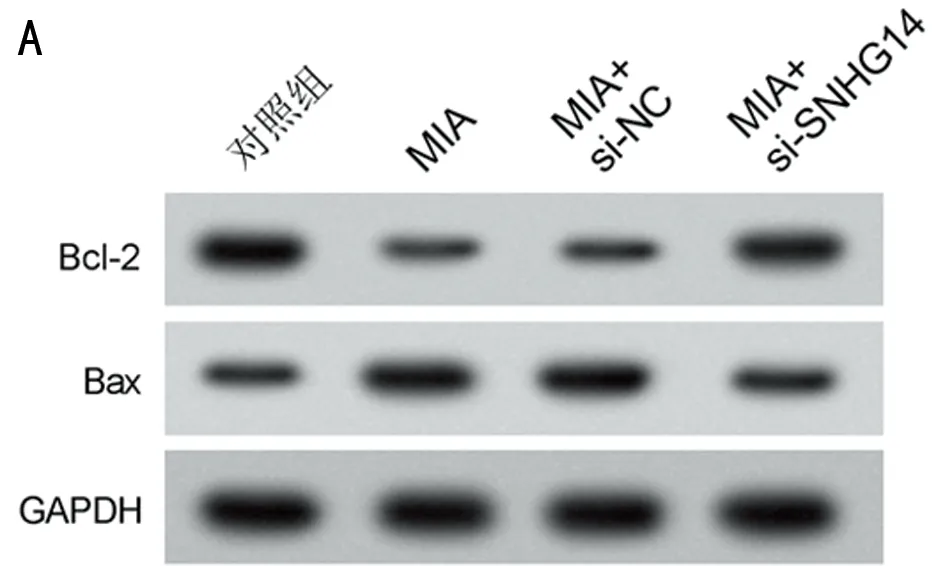
圖1 抑制lncRNA SNHG14表達對MIA誘導軟骨細胞凋亡的影響;A 細胞凋亡流式圖;B 凋亡相關蛋白表達
表2 抑制lncRNA SNHG14表達對MIA誘導軟骨細胞損傷的影響
2.3 lncRNA SNHG14靶向調控miR-149-5p的表達(LncBase Predicted v.2) StarBase預測顯示lncRNA SNHG14的序列中含有與miR-149-5p互補的核苷酸序列。雙熒光素酶實驗顯示,與轉染miR-NC的WT-SNHG14比較,轉染miR-149-5p的熒光素酶活性降低(P<0.05),而轉染miR-NC和miR-149-5p的MUT-SNHG14中,熒光素酶活性無差異(P>0.05)。與pcDNA組比較,pcDNA-SNHG14組的miR-149-5p表達水平降低(P<0.05);與si-NC組比較,si-SNHG14組miR-149-5p表達水平升高(P<0.05)。見圖2,表3、4。
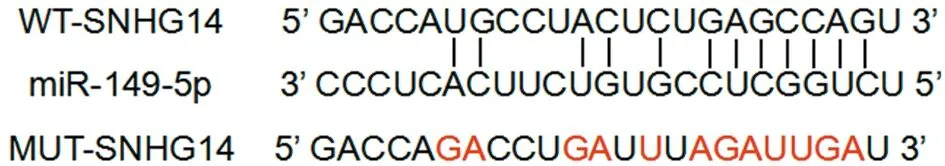
圖2 lncRNA SNHG14的序列中含有與miR-149-5p互補的核苷酸序列
表3 雙熒光素酶報告實驗
表4 lncRNA SNHG14調控miR-149-5p的表達
2.4 過表達miR-149-5p對MIA誘導軟骨細胞損傷的影響 與MIA+miR-NC組比較,MIA+miR-149-5p組miR-149-5p的表達水平升高,細胞凋亡率降低,Bcl-2表達水平升高,Bax表達水平降低,MDA表達水平降低,SOD和CAT活性升高(P<0.05)。見圖3,表5。
表5 過表達miR-149-5p對MIA誘導軟骨細胞損傷的影響
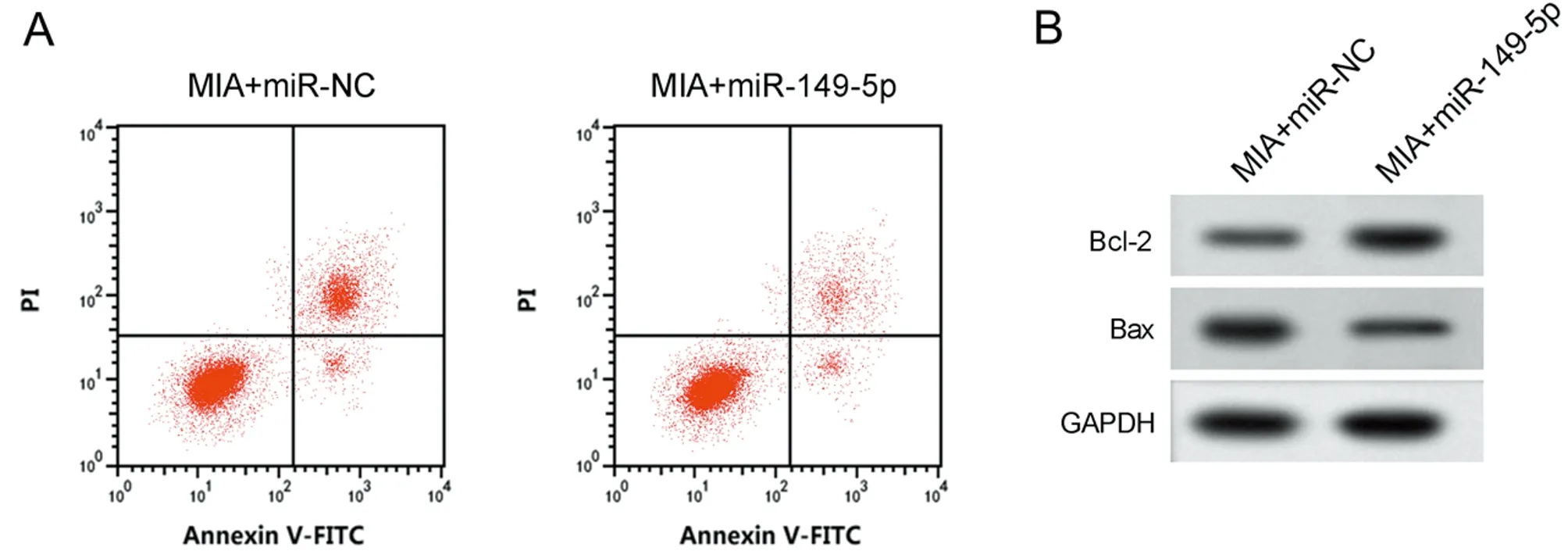
圖3 過表達miR-149-5p對MIA誘導軟骨細胞損傷的影響;A 凋亡相關蛋白表達;B 細胞凋亡流式圖
2.5 干擾miR-149-5p表達逆轉了抑制lncRNA SNHG14表達對MIA誘導軟骨細胞損傷的作用 與MIA+si-SNHG14+anti-miR-NC組比較, MIA+si-SNHG14+anti-miR-149-5p組miR-149-5p的表達水平降低,細胞凋亡率升高,Bcl-2表達水平降低,Bax表達水平升高,MDA表達水平升高,SOD和CAT活性降低(P<0.05)。見圖4,表6。
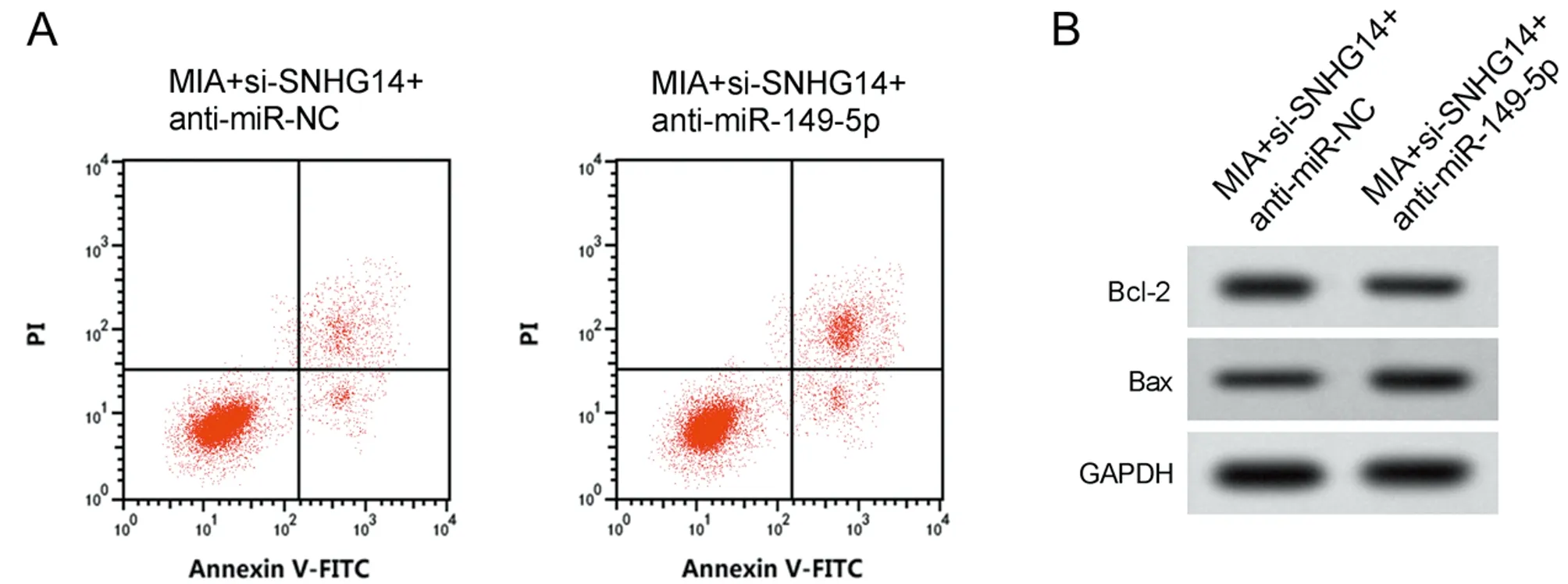
圖4 干擾miR-149-5p表達逆轉了抑制lncRNA SNHG14表達對MIA誘導軟骨細胞凋亡的作用;A 凋亡相關蛋白表達;B 細胞凋亡流式圖
表6 干擾miR-149-5p表達逆轉了抑制lncRNA SNHG14表達對MIA誘導軟骨細胞損傷的作用
3 討論
研究發現,lncRNA的異常表達參與了許多炎癥相關疾病。在帕金森病小鼠腦組織中,lncRNA SNHG14表達上調,在魚藤酮誘導的MN9D細胞中,lncRNA SNHG14過表達,干擾lncRNA SNHG14通過調控miR-133b/α-synuclein通路減輕魚藤酮誘導的神經元損傷[8]。在急性肺損傷中,lncRNA SNHG14和WISP1表達升高,miR-34c-3p表達降低,下調lncRNA SNHG14和過表達miR-34c-3p均降低IL-18、IL-1β、TNF-α和IL-6的表達水平,下調lncRNA SNHG14通過miR-34c-3p/WISP1軸緩解LPS誘導的急性肺損傷[9]。在腦缺血再灌注誘導的PC12細胞損傷中,lncRNA SNHG14通過miR-136-5p/ROCK1軸加重神經損傷和炎性反應[10]。在小鼠大腦中動脈閉塞模型和小膠質細胞缺氧缺糖模型中,lncRNA SNHG14過表達,敲減lncRNA SNHG14分別上調和下調miR-145-5p和PLA2G4A的表達,從而逆轉缺氧缺糖對BV-2細胞的激活作用[11]。在低密度脂蛋白誘導的動脈粥樣硬化細胞模型中,lncRNA SNHG14表達上調,lncRNA SNHG14通過miR-186-5p/WIPF2軸調節細胞增殖和凋亡[12]。在AngⅡ誘導心肌細胞肥大中,lncRNA SNHG14和PCDH17均可逆轉敲除SP1基因對心肌細胞肥大的抑制作用[13]。在小鼠小膠質細胞缺血-再灌注損傷中,lncRNA SNHG14表達上調,右美托咪定可抑制小鼠小膠質細胞的活化,并下調lncRNA SNHG14表達,過表達lncRNA SNHG14通過上調HMGB1表達逆轉了右美托咪定對小鼠小膠質細胞缺血-再灌注損傷的治療作用[14]。與上述結果相似,本實驗結果顯示,在MIA誘導的軟骨細胞中,lncRNA SNHG14表達水平升高,細胞凋亡率升高,Bcl-2表達水平與SOD和CAT活性降低,Bax和MDA表達水平升高;抑制lncRNA SNHG14表達降低了細胞凋亡率,提高了Bcl-2表達水平與SOD和CAT活性,降低了Bax和MDA表達水平。提示抑制lncRNA SNHG14表達可抑制細胞凋亡和氧化應激反應,從而減輕MIA誘導的軟骨細胞損傷。
研究表明lncRNA SNHG14可作為競爭性內源RNA負調控炎癥相關miRNA表達來促進相關疾病進展[5-6,8-13]。本實驗的StarBase預測顯示,lncRNA SNHG14的序列中含有與miR-149-5p互補的核苷酸序列,雙熒光素酶報告實驗結果顯示,在WT-SNHG14中,miR-149-5p組的雙熒光素酶活性顯著降低,且過表達和抑制lncRNA SNHG14分別降低和提高miR-149-5p的表達水平,即lncRNA SNHG14靶向負調控miR-149-5p表達。研究表明,miR-149參與了炎癥相關疾病的發生發展,在ox-LDL處理的人血管內皮細胞中,miR-149-5p表達降低,Circ 0124644通過miR-149-5p/PAPP-A軸加重內皮細胞[15]。褪黑素通過上調miR-149表達抑制缺血誘導的成肌細胞纖維化[16]。在缺血再灌注(I/R)誘導的腦損傷中,miR-149-5p表達下調,miR-149-5p通過Sirt1/p53軸在神經元細胞死亡中發揮調節作用[17]。在LPS誘導的肺損傷中,miR-149表達下調,lncRNA MALAT1通過miR-149/MyD88軸促進急性肺損傷的炎性反應[18]。與前人研究結果一致,在MIA誘導的軟骨細胞損傷中,miR-149-5p表達水平降低,過表達miR-149-5p降低了細胞凋亡率,提高了Bcl-2表達水平與SOD和CAT活性,降低了Bax和MDA表達水平。提示過表達miR-149-5p可抑制細胞凋亡和氧化應激反應,以減輕MIA誘導的軟骨細胞損傷。且下調miR-149-5p表達逆轉了抑制lncRNA SNHG14表達對軟骨細胞凋亡和氧化應激反應的影響,提高了細胞凋亡率,降低了Bcl-2表達水平與SOD和CAT活性,提高了Bax和MDA表達水平。提示lncRNA SNHG14可能通過調控miR-149-5p表達影響MIA誘導的軟骨細胞凋亡和氧化應激反應。
綜上所述,lncRNA SNHG14在MIA誘導的軟骨細胞中上調,抑制lncRNA SNHG14表達通過靶向上調miR-149-5p表達調控MIA誘導的軟骨細胞凋亡和氧化應激反應。這意味著lncRNA SNHG14可能成為治療OA的新靶點,但僅限于體外實驗,lncRNA SNHG14在體內的作用以及調控機制還有待進一步研究。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產業(2016年3期)2016-05-17 04:32:12
學苑創造·A版(2015年11期)2016-01-14 09:03:27
