不同月份不同樹齡香榧葉片光合特性的變化
2022-03-01 07:12:22陸呂佳胡淵淵戴文圣索金偉宋麗麗吳家勝喻衛武
浙江農林大學學報 2022年1期
關鍵詞:研究
劉 璉,陸呂佳,胡淵淵,2,戴文圣,2,3,索金偉,2,3,宋麗麗,2,3,吳家勝,2,3,喻衛武,2,3,樓 君,4
(1. 浙江農林大學 省部共建亞熱帶森林培育國家重點實驗室,浙江 杭州 311300;2. 浙江農林大學 國家林業草原香榧工程技術研究中心,浙江 杭州 311300;3. 浙江農林大學 香榧產業國家創新聯盟,浙江 杭州 311300;4. 浙江省杭州市富陽區農業技術推廣中心 林業站,浙江 杭州 311400)
隨季節變化,植物生長環境中的光照強度、氣溫等均會發生改變。一般認為,植物光合作用的季節性變化是其對氣溫的適應性響應[1]。光能和二氧化碳(CO2)分別是光合作用的動力和基本原料,研究光合作用對光強和CO2的響應特征有助于闡明植物在環境變化中的生理生態適應性[2?3]。因此,研究植物光合作用的季節性變化對預測樹種響應氣候變化引起的氣溫改變具有重要的意義[4]。研究不同季節植物葉片的氣體交換參數(Jmax和Vcmax)、葉綠素含量、葉氮含量及氮分配的變化是揭示植物葉片光合作用季節性變化的生理機制的一種方法[1,5?12]。香榧Torreya grandis ‘Merrillii’種籽經炒制后食用,風味香醇,營養價值高,是中國珍稀的上等干果。隨著市場需求的增大,從2008年開始,浙江香榧的種植面積迅速增大。截至2019年,浙江省香榧種植面積達4.8萬hm2,已成為中國南方山區重要的經濟樹種[13]。在生產上,通常采用2年生實生榧樹Torreya grandis作為砧木嫁接香榧,4~5 a開始掛果,15 a后達盛果期。葉片或樹體的生理年齡被認為是評估其對逆境響應能力時需協同考慮的重要因子,因為不同生理年齡的葉片或樹體生長對碳源的需求、酶活性、激素活性、水、冠層微環境和葉片的形態結構等均會發生變化[14]。隨樹齡的增加,樹體會由于光合作用/呼吸作用比率降低發生碳饑餓,樹體老化[15]。橡樹Quercus cerris葉片的光合速率隨樹齡的增加呈降低趨勢[16]。近年來,對于香榧的研究主要集中在藥用價值和幼苗與生長環境的關系上[17?19],對香榧不同樹齡不同季節葉片光合特性方面的研究國內尚未見報道。本研究以6年生嫁接苗(初掛果期)和16年嫁接苗(盛果期)香榧葉片為研究對象,從表觀光合特性和光合內部機構系統地探討不同樹齡香榧葉片光合作用及對氣溫的響應機制,為制定香榧高效栽培措施提供理論依據。
1 材料與方法
1.1 材料
試驗區位于浙江省杭州市臨安區浙江農林大學香榧基地,29°56′~30°23′N,118°51′~119°52′E,屬于亞熱帶季風氣候區,溫暖濕潤,光照充足,雨量充沛。土壤類型為黃壤土,林地肥料以復合肥為主,配施農家肥。供試材料為16年生和6年生香榧嫁接苗。其中,16年生香榧嫁接苗為2000年采用嫁接苗造林,林分密度為450株·hm?2,砧木為2年生實生榧樹,接穗為1年生香榧,平均地徑為11.5 cm,平均樹高為2.6 m,記為16-a;6年生香榧嫁接苗為2010年采用嫁接苗造林,砧木為2年生實生榧樹,接穗為1年生香榧,平均地徑為8.3 cm,平均樹高為1.8 m,記為6-a。香榧于每年4月底萌發新葉,約50 d葉片生長成為成熟葉[20]。本研究分別以2017年春季(5月)的未成熟葉片、夏季(8月)的成熟葉和秋季(11月)的成熟葉的當年生葉片為材料。
1.2 方法
1.2.1 凈光合速率和暗呼吸速率的測定 氣象數據取自杭州市臨安區氣象局。選擇晴朗無風的天氣,采用Li-6400便攜式光合儀(Licor-6400,Licor,美國),使用可控光簇狀葉室(22 L)進行香榧當年生葉片的光響應測定。測定時,葉室內CO2摩爾分數設置為400 μmol·mol?1,由便攜式CO2小鋼瓶控制CO2摩爾分數;根據氣溫(5月的最低和最高氣溫分別為16.2和27.2 ℃;8月分別為21.0和32.5 ℃;11月分別為13.3和24.7 ℃),將5月、8月及11月葉室測定溫度分別控制在25、30和20 ℃,葉室的空氣相對濕度約50%;采用RGB-18白光光源,設定氣體流動速度為 500 μmol·s?1。正式記錄前,先用800 μmol·m?2·s?1光強對葉片進行光誘導20 min,待葉片活化穩定后,再記錄測定數據,記為凈光合速率;再將光強調至0 μmol·m?2·s?1,并用黑色布將測定葉室罩住,待穩定后再記錄測定數據,記為暗呼吸速率。重復3~5次,取平均值作為測定結果,以上測定均在9:00—11:00完成。
1.2.2 CO2響應曲線 成熟香榧葉片的光飽和點為 (779.0±71.9) μmol·m?2·s?1[21],因此,對葉片進行CO2響應曲線測定時,光照強度設為800 μmol·m?2·s?1。同時,為模擬生長環境的溫度,5月、8月及11月葉室溫度分別控制為25、30和20 ℃。在開始CO2響應曲線測定前,先將葉片在CO2摩爾分數為400 μmol·mol?1和光強為 800 μmol·m?2·s?1下持續 30 min,再依次按不同的 CO2摩爾分數 (400、300、200、100、50、400、800、1 200、1 600、1 800、2 000 μmol·mol?1)進行凈光合速率 (Pn)的測定,再將數據通過FARQUHAR等[5]的模型擬合CO2響應曲線。當RuBP充足時,Rubisco是光合速率(Ac)的限制因子,公式如下:

式(1)中:Vcmax是最大RuBP羧化速率;Kc和Ko是Rubisco羧化作用和加氧作用的Michaelis常數;Ci和O分別為細胞間隙中的CO2和氧氣(O2)摩爾分數;Rd是光下的呼吸速率;Г*是不含暗呼吸的CO2補償點。本研究采用von CAEMMERER等[22]模型中的Rubisco動力學參數進行數據擬合。
當光合速率(Aj)受RuBP更新的限制,可表示為:

式(2)中:Jmax是光飽和時用于RuBP再生的光合電子傳遞速率。通過非線性最小二乘法得到擬合A—Ci曲線,分析得出 Vcmax和 Jmax。葉片的 Vcmax和 Rd可根據 A—Ci曲線 (Ci<250 μmol·mol?1)得出;Jmax則根據 A—Ci曲線 (Ci>600 μmol·mol?1)獲得。
CO2響應曲線的測定在9:00—11:30和14:00—16:00完成。每株樹為1個重復,每個處理為5~6個重復。
1.2.3 比葉重(SLW)測定 在氣體交換測定完成后,收集葉片并帶回實驗室,將測定枝上的披針葉全部剪下,用透明膠帶把葉片粘貼在復印紙上,然后把復印出的針葉剪下稱量,葉面積根據單位復印紙的質量計算。隨后將葉片放置烘箱,保持85 ℃烘干至恒量,稱干質量。比葉重(g·m?2)=干質量/葉面積。
1.2.4 葉綠素 (Chl)測定 采香榧新鮮葉片約 0.02 g,剪碎浸入盛有 8 mL 提取液 (體積分數為 95% 乙醇)的離心管中,密封避光低溫浸提至無色[20]。根據用紫外分光光度計(UV 2500,島津)檢測波長在649和664 nm處的吸光度。記錄所測葉片的鮮質量和面積,計算葉綠素含量(mg·dm?2)。
1.2.5 葉片氮含量、光合氮素利用效率(PNUE)及氮素在光合機構中的分配 采香榧葉片,用凱式定氮法[20]測定單位面積上的葉氮含量(NA,g·m?2)。以飽和最大光合速率(Amax)與NA的比值表示光合氮素利用效率 (photosynthetic nitrogen utilization efficiency,PNUE)[20]。植物葉氮在光合機構中的分配[12],即分配到Rubisco的氮素(Vcmax/NA)、RuBP再生作用的氮素(Jmax/NA)和捕光組分的氮素(Chl/NA)。
1.3 數據處理與分析
用Excel處理數據,通過SigmaPlot 12.5作圖,使用Origin 8.0繪制光合—光響應曲線和光合—CO2響應曲線,用SPSS檢驗顯著性差異。
2 結果與分析
2.1 不同月份不同樹齡香榧葉片比葉重的變化
從圖1可以看出:無論樹齡大小,8月和11月香榧葉片的比葉重顯著大于5月香榧葉片的比葉重(P<0.05),8月和11月香榧葉片比葉重之間無顯著差異(P>0.05)。5月,16-a的比葉重顯著高于6-a(P<0.05);8月和11月,樹齡之間的比葉重無顯著差異 (P>0.05)。
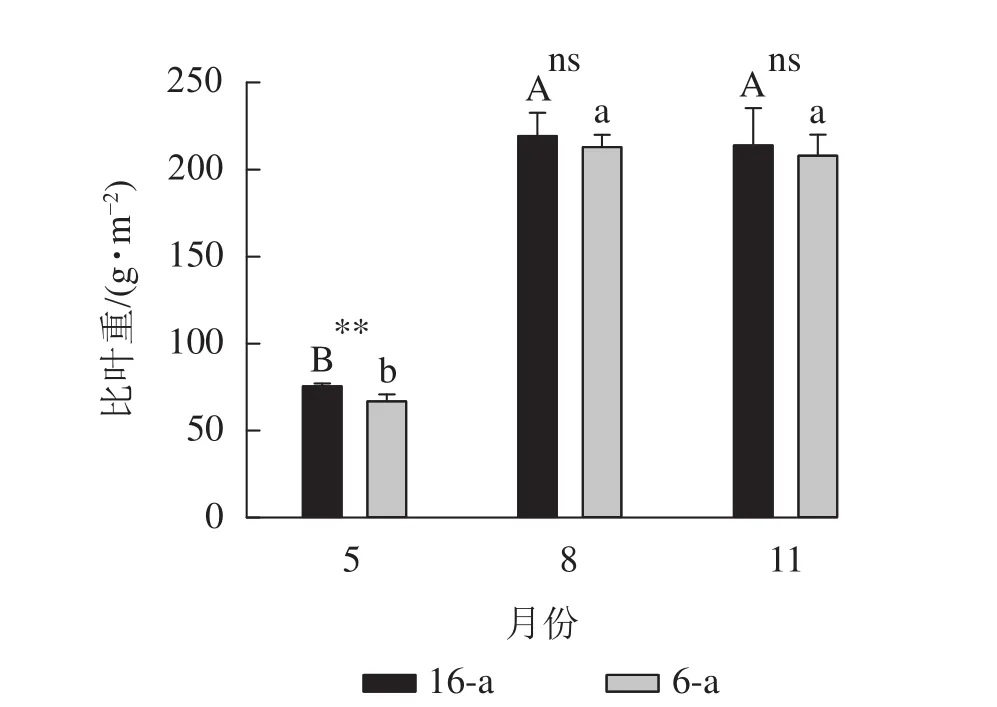
圖1 不同月份不同樹齡香榧葉片比葉重的變化Figure 1 Changes in specific leaf weight (SLW) of T. grandis‘Merrillii’ at different tree ages among different months
2.2 不同月份不同樹齡香榧葉片凈光合速率和暗呼吸速率的變化
無論樹齡大小,香榧葉片的Pn在5月較低,8月快速上升達到峰值,且保持穩定至11月(圖2)。5—11月,6-a和16-a香榧葉片的Pn均無顯著差異(P>0.05)。5月6-a香榧葉片的Rd顯著高于16-a(P<0.05),但8—11月兩者之間無顯著差異(P>0.05)。
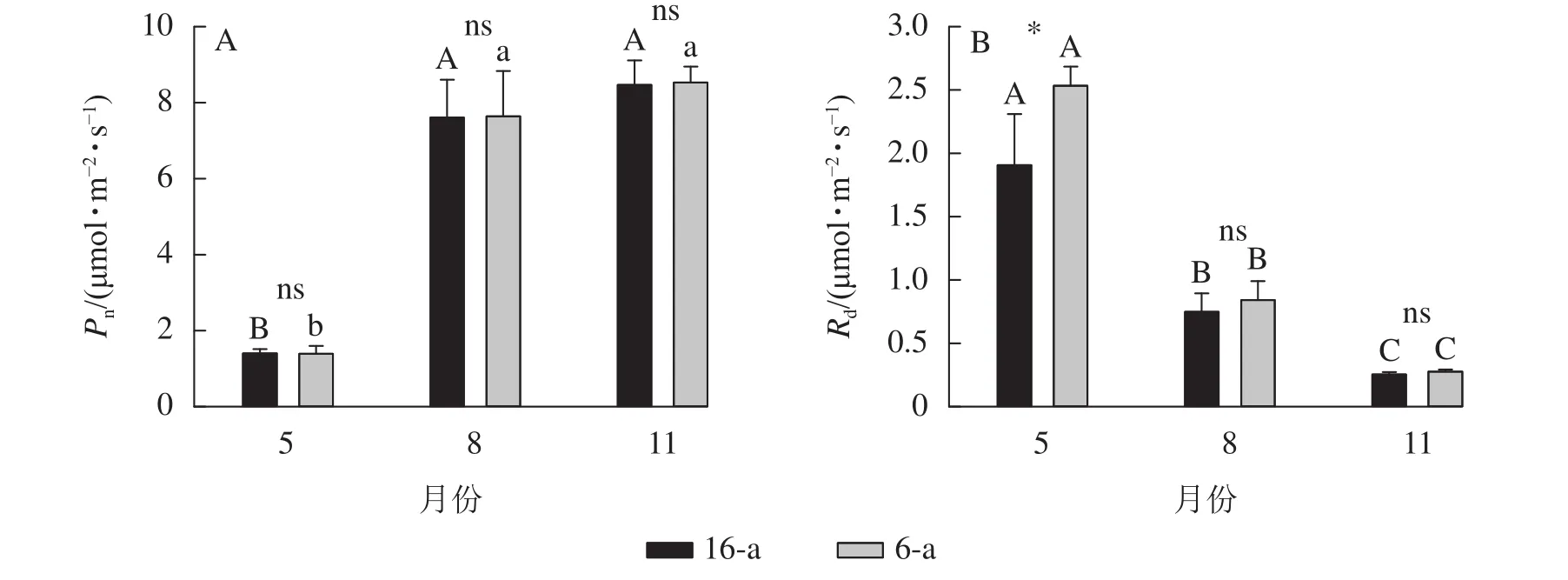
圖2 不同月份不同樹齡香榧葉片凈光合速率(Pn)和暗呼吸速率(Rd)的變化Figure 2 Changes in net photosynthetic rate (Pn) and dark respiration rate (Rd) of T. grandis ‘Merrillii’ at different tree ages among different months
2.3 不同月份不同樹齡香榧葉片CO2響應曲線的變化
由圖3可知:無論樹齡大小,11月香榧葉片的CO2飽和點顯著高于8月(P<0.05);8月時,16-a和 6-a 香榧葉片的 CO2飽和點分別為 725和 550 μmol·mol?1,11月時分別為 740和 1075 μmol·mol?1。由圖4可知:無論樹齡大小,5月香榧葉片的Vcmax和Jmax均顯著低于8月和11月(P<0.05)。與8月相比,11月16-a和6-a的Vcmax分別降低了54.4%和64.6%,而Jmax分別降低了22.1%和25.8%。
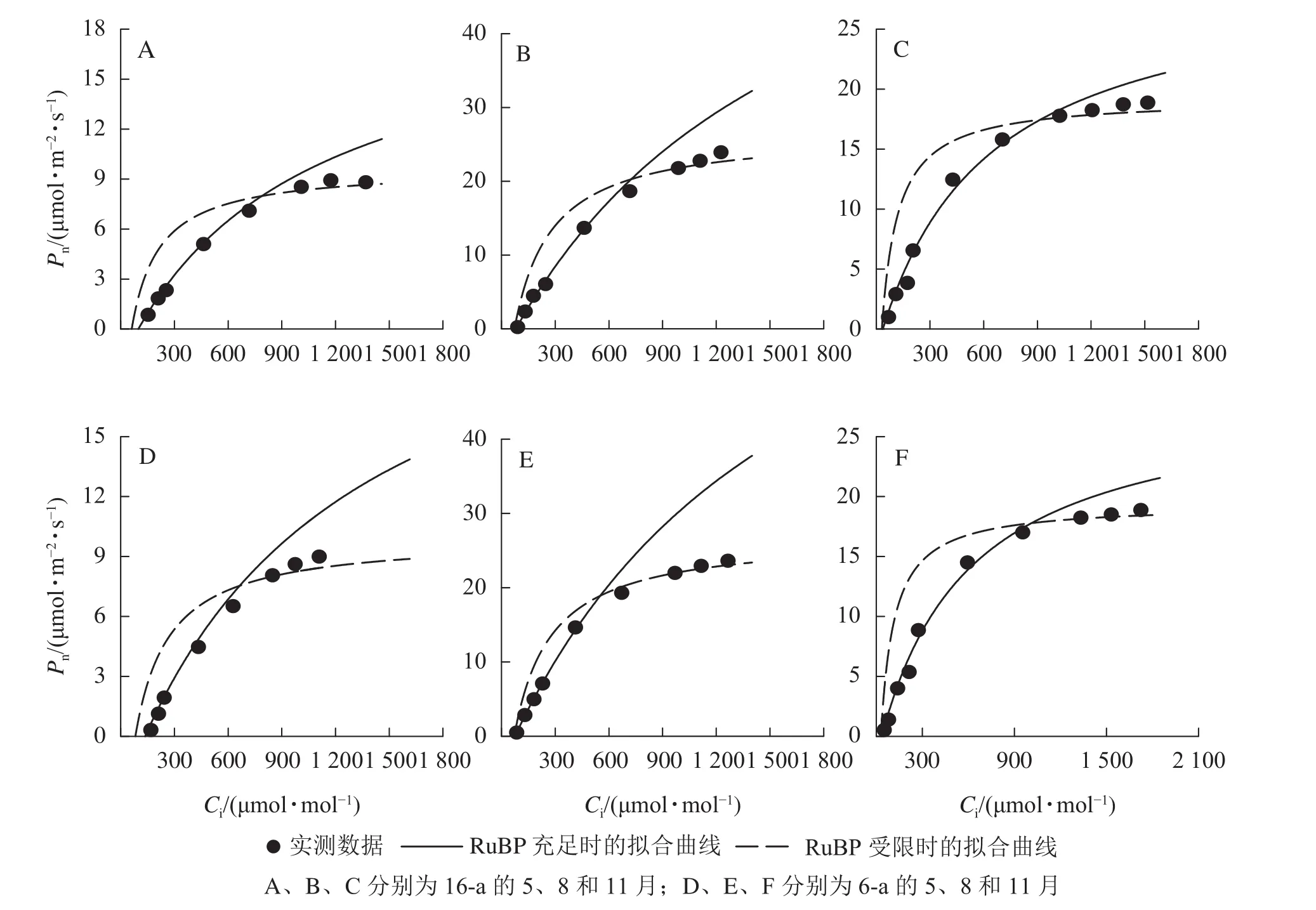
圖3 不同月份不同樹齡香榧葉片CO2光合響應曲線的變化Figure 3 Changes in CO2 response curves of photosynthesis in leaves of T. grandis ‘Merrillii’ at different tree ages among different months
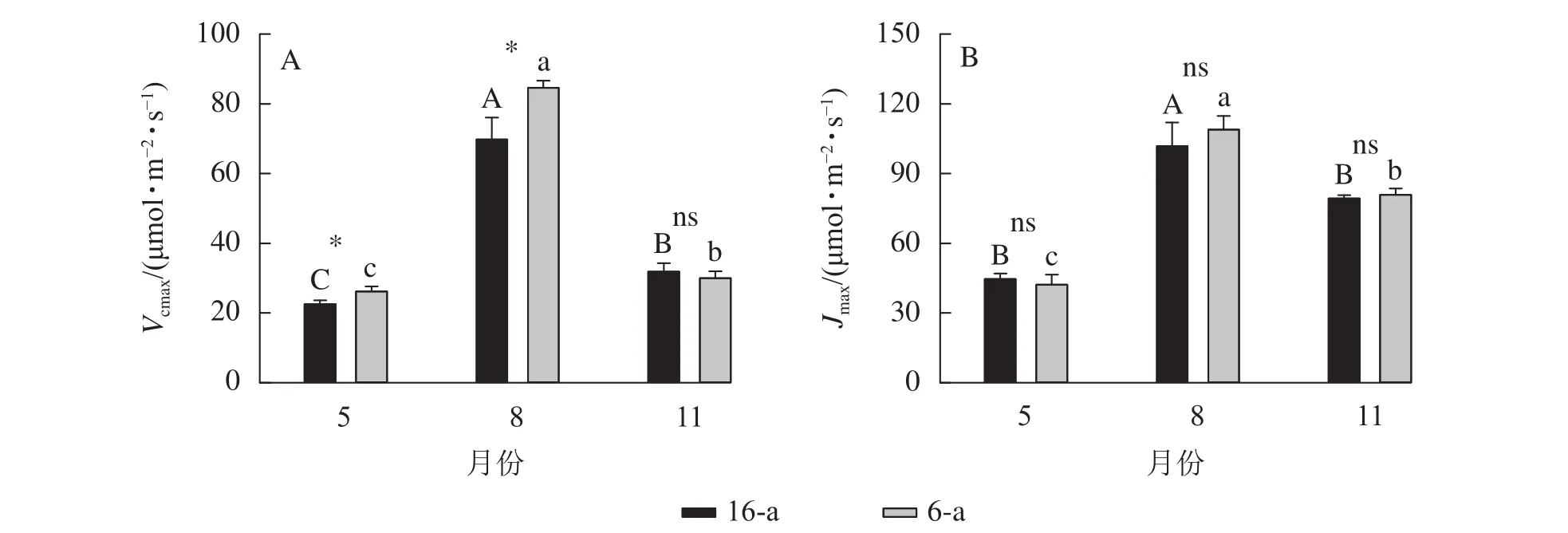
圖4 不同月份不同樹齡香榧葉片的 Vcmax 和 Jmax 的變化Figure 4 Changes in Vcmax and Jmax in leaves of T. grandis ‘Merrillii’ at different tree ages among different months
2.4 不同月份不同樹齡香榧葉片葉色素含量的變化
圖5顯示:無論樹齡大小,5月香榧葉片的葉綠素含量顯著高于8和11月(P<0.05),且16-a香榧葉片的葉綠素含量均極顯著高于6-a香榧葉片的葉綠素含量(P<0.001)。
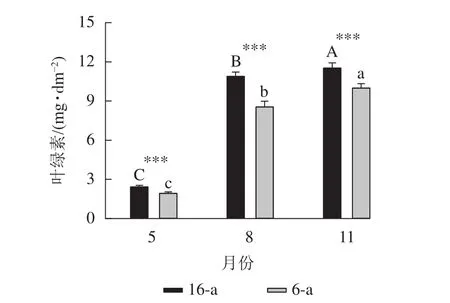
圖5 不同月份不同樹齡香榧葉片葉綠素含量的變化Figure 5 Changes in chlorophyll content in leaves of T. grandis‘Merrillii’ at different tree ages among different months
2.5 不同月份不同樹齡香榧葉片氮含量、光合氮素利用效率(PNUE)及氮素在光合機構中的分配
由圖6可見:無論樹齡大小,與5月相比,8月16-a和6-a香榧葉片的氮含量分別下降了23.8%和17.6%,差異顯著(P<0.05);與8月相比,11月16-a和6-a香榧葉片的氮含量均增加了11.0%,差異顯著(P<0.05);且6-a香榧葉片的PNUE均極顯著高于16-a (P<0.001);其中8月和11月時,6-a香榧葉片的PNUE分別為16-a香榧葉片的1.14和1.20倍。
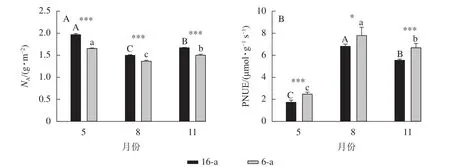
圖6 不同月份不同樹齡單位面積上的香榧葉片氮含量(NA)和光合氮素利用效率(PNUE)的變化Figure 6 Changes in nitrogen content per area (NA) and photosynthetic nitrogen utilization efficiency (PNUE) in leaves of T. grandis‘Merrillii’ at different tree ages among different months
由表1可見:無論樹齡大小,與5月相比,8月香榧葉片的Jmax/Vcmax顯著降低(P<0.05);而與8月相比,11月香榧葉片的Jmax/Vcmax顯著增加(P<0.05)。5月和8月時,16-a香榧葉片的Jmax/Vcmax顯著高于6-a (P<0.05)。無論樹齡大小,與5月相比,8月香榧葉片的Jmax/NA、Vcmax/NA和Chl/NA均顯著增加(P<0.05);與8月相比,11月的16-a和6-a的Jmax/NA分別降低了30.1%和29.4%,而Vcmax/NA分別降低了60.9%和67.8%,而Chl/NA無顯著變化(P>0.05)。

表1 不同月份不同樹齡香榧葉片 Jmax/Vcmax、Jmax/NA、Vcmax/NA 和 Chl/NA 的變化Table 1 Changes in Jmax/Vcmax, Jmax/NA, Vcmax/NA and Chl/NA in leaves of T. grandis ‘Merrillii’ at different tree ages among different months
3 討論與結論
植物干物質的90%~95%來源于光合作用。本研究結果顯示:5月時,16-a香榧葉片的比葉重顯著高于6-a葉片的,而此時16-a與6-a香榧葉片的Pn之間無顯著差異,但6-a香榧葉片的Rd則明顯大于16-a香榧葉片,這可能是由于呼吸消耗相對較低的16-a香榧葉片更有利用干物質的積累,可為盛果期的豐產提供較多的物質基礎。5和8月時,16-a和6-a香榧葉片的SLW均增至最大,這與其快速增長的Pn、太陽總輻射和日照長度的增長密切相關。杭州地區8月的太陽總幅射和日照時數均明顯多于5月[23]。
本研究結果顯示:與5月相比,無論樹齡大小,8月香榧葉片的Pn和SLW均明顯增大;但8月香榧葉片的NA均顯著降低,這可能是由于葉片光合作用增強,碳水化合物快速積累,而使其葉片中的氮含量被稀釋了[9?10]。有研究表明:隨著山胡椒屬Lindera植物Lindera umbellate落葉植物葉片的衰老,葉片中可代謝的氮素(如光合相關蛋白質)會降解成可再利用的氮素[24],再轉移至其他發育的器官中[25]。無論樹齡大小,與8月相比,11月香榧葉片的Pn略有上升,且NA顯著增加,表明當年生的香榧葉片并未啟動衰老過程,其所增加的葉片氮含量可能來源于其他多年生衰老的葉片。這與趙雨馨[26]的研究結果相一致,12月香榧葉片光合速率顯著高于7和10月。無論哪個月份,16-a香榧葉片的NA均顯著高于6-a香榧葉片的,但其PNUE卻均顯著低于6-a香榧葉片的,表明在生產上須注意氮肥的施用,尤其是像16-a這種進入盛果期的香榧。通常,葉片中50%以上的氮素分配到光合作用相關的蛋白質中,如Rubisco、捕光蛋白等[8]。有研究表明:Vcmax的降低通常是由于Rubisco含量的減少[27]或Rubisco活化位點降低[28]。矮垂頭菊Daphniphyllum humile當年生葉片的氮含量、Rubisco含量隨季節變化(從春季到秋季)呈逐漸增加的趨勢,但其葉片的飽和光合速率穩定不變[9]。本研究結果顯示:無論樹齡大小,與8月相比,11月香榧葉片SLW、Pn無顯著變化,而其NA均顯著增加,但Vcmax均顯著降低,表明當年生的香榧葉片Vcmax的降低可能是Rubisco活化位點的減少引起的。8月時,6-a香榧葉片的Vcmax顯著高于16-a香榧葉片的,但兩者之間的Amax無顯著差異,這可能是由于其Rubisco未完全激活,僅起著儲存氮素的作用[29]。已有研究表明:相比較易招引昆蟲的氨基酸形式的氮素形態,Rubisco是更好的氮素儲存形式[30]。
與20世紀相比,大氣CO2摩爾分數和溫度均明顯升高,且未來還將會持續升高[31?32]。本研究結果顯示:無論樹齡大小,香榧葉片的CO2飽和點均大于500 μmol·mol?1,表明香榧葉片的光合作用在當今大氣的CO2摩爾分數下仍未達到飽和,對高摩爾分數CO2有一定的適應性,這可能也是香榧的結果期可達百年甚至千年的原因之一。研究表明:Jmax/Vcmax可以指示RuBP再生作用和羧化作用2個過程的相關蛋白質的分配情況[33?34]。本研究結果顯示:無論樹齡大小,隨溫度的降低(8—11月),香榧葉片的Jmax/Vcmax均明顯增大,這與前人的研究結果一致[7, 35]。當光合速率處于受RuBP羧化作用限制的階段(Ac)時,碳同化對溫度的變化不太敏感,由于低溫下光呼吸也受抑制,有利于促進CO2的羧化固定,降低的光呼吸消耗從一定程度上彌補了低溫引發減弱的RuBP羧化作用[36],而當光合速率受RuBP再生作用限制的階段(Aj),其碳同化較易受到低溫的影響[36?37],因此,在高CO2摩爾分數下,溫度對其光合碳同化的影響遠大于其在低CO2摩爾分數下的影響。由此可知,較低溫下(與8月相比,11月香榧苗木生長的環境溫度約下降了10 ℃以上),香榧葉片通過Jmax的相對增加(下降幅度相對較小)來緩減低溫對RuBP再生作用的影響,即植物通過改變Jmax/Vcmax來調整Ac和Aj之間的失衡狀態[37?38]。
此外,調整氮素在光合機構的分配常被認為是植物適應溫度變化的重要機制[11]。本研究結果顯示:隨著氣溫的降低(8—11月),香榧葉片分配到Rubisco的氮素(Vcmax/NA)顯著減少,其下降幅度明顯大于分配到其他光合蛋白的氮素(Jmax/NA和Chl/NA)。這可能是由于不同光合蛋白的理化性質引起的,即Rubisco是一種可溶性蛋白,而反應中心和捕光蛋白(如LHCII)是類囊體膜蛋白,因此,Rubisco中氮素再利用效率明顯高于其他捕光蛋白(如LHCII)中的氮素。有研究表明:在高CO2摩爾分數下,如果將氮素相對較少地分配到Rubisco,且相對較多地分配到RuBP再生作用相關的光合蛋白上,則會大大提高氮素的光合利用效率[37, 39]。本研究結果顯示:隨著氣溫的降低(8—11月),6-a香榧葉片Vcmax/NA的下降幅度明顯大于16-a葉片的,而6-a與16-a香榧葉片Jmax/NA的下降幅度相差不多,因此,低溫下(11月),6-a香榧葉片分配到RuBP羧化作用的氮素較少和分配到RuBP再生作用的氮素較多是其葉片PNUE相對較高的原因。此外,香榧是一種長壽的經濟樹種,是否能通過調控氮素在光合組分中的分配來適應逐漸變暖、CO2摩爾分數持續上升的全球氣候變化?因此,未來應該重點研究香榧古樹與新種植的香榧樹對溫度和CO2摩爾分數倍增變化的適應性是否存在差異。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
