肺癌組織MTA1、VEGF?C表達變化及與患者臨床病理特征、預后關系
2022-02-22 06:35:02趙麗霞任成波馬峰李占林趙峻峰
廣東藥科大學學報 2022年1期
關鍵詞:肺癌
趙麗霞,任成波,馬峰,李占林,趙峻峰
(河北北方學院附屬第一醫院1.腫瘤內科;2.放療科;3.中醫科,河北 張家口 075000)
肺癌是世界范圍內發病率、死亡率最高的惡性腫瘤,且受空氣污染、煙草流行、人口老齡化等因素影響,其發病率、死亡率呈上升趨勢[1]。盡管隨著當前基礎醫學及相關學科發展,肺癌診療手段取得一定進步,但肺癌患者總體預后仍不理想,探究肺癌發生機制及與肺癌發展、預后相關的生物學標記物是臨床研究重要方向[2]。轉移相關基因1(metasta‐sis?associated gene 1,MTA1)蛋白分布于人體多種組織器官,研究認為,MTA1為脫乙酰基復合物的亞單位之一,可通過介導組蛋白去乙酰化、核小體重構過程參與基因表達調控,可能與腫瘤發生和轉移有關[3]。血管內皮生長因子C(vascular endothelial growth factor C,VEGF?C)與血管內皮生長因子同源,可誘導淋巴管內皮細胞增殖、遷移并影響其通透性,還能促進微血管形成,被認為與腫瘤淋巴管、病理性血管生成關聯密切[4]。國內外已有研究指出[5?6],在多種惡性腫瘤如肝癌、乳腺癌、胃癌、鼻咽癌等組織中可檢出MTA1、VEGF?C過表達,二者可能與腫瘤浸潤轉移有關。不過MTA1、VEGF?C在肺癌中的研究報道尚不多見,其與肺癌病理類型、預后等關系并無定論。為此,本研究通過免疫組織化學法檢測肺癌組織MTA1、VEGF?C表達情況,并分析其與患者臨床病理特征、預后關系,旨在為肺癌臨床早期診療及預后評估提供新思路。現報道如下。
1 資料與方法
1.1 一般資料
收集河北北方學院附屬第一醫院2016年1月-2018年9月經病理確診的200例肺癌組織標本及50例癌旁正常(距腫瘤邊緣>5 cm)組織標本。納入標準:患者均經手術或病理活檢明確原發性肺癌診斷,且具有明確的腫瘤分期[7];腫瘤均為單發;組織標本保存完好;年齡35~75歲;均獲取完善臨床病史及隨訪信息。排除標準:復發腫瘤病例;合并其他部位原發性腫瘤;無足夠可用于免疫組織化學分析的標本;術前已接受放化療及其他抗腫瘤治療;存在不明原因異常生化指標;失訪。200例患者中男149例,女51例;年齡41~75歲,其中<60歲117例,≥60歲83例;病理類型:腺癌94例,鱗癌79例,小細胞癌27例;分化程度:低分化癌68例,中分化癌73例,高分化癌59例;TNM分期:Ⅰ期29例,Ⅱ期62例,Ⅲ期84例,Ⅳ期25例;淋巴結轉移78例,無淋巴結轉移122例。本研究經醫院倫理委員會批準,患者或家屬簽署知情同意。
1.2 方法
1.2.1 免疫組化染色步驟 10%(φ)中性福爾馬林溶液固定標本,石蠟包埋,以4μm厚度連續切片,烤片機烘烤(58~60℃)60 min;切片經二甲苯脫蠟10 min,于梯度酒精脫苯處理后,以蒸餾水沖洗5 min;切片滴加3%H2O2,于室溫下孵育10 min,以阻斷內源性過氧化物酶,然后蒸餾水沖洗3次,再于PBS液浸泡5 min;切片置于枸櫞酸鹽緩沖液(濃度0.01 mmol/L,pH=6),高溫高壓下修復10 min,冷卻后用PBS液沖洗3次;加入封閉用正常血清工作液,室溫下封閉15~20 min,傾去,勿洗;滴加稀釋(1∶100)的一抗(羊抗人MTA1多克隆抗體、羊抗人VEGF?C多克隆抗體,均購自美國SANTA CRUZ公司),4℃孵育過夜,PBS液沖洗3次;滴加二抗工作液(生物素標記羊抗兔ⅠgG),室溫孵育15~20 min,PBS液沖洗3次;滴加辣根酶標記鏈酶卵白素工作液,室溫孵育15~20 min,PBS液沖洗3次;以DAB顯示(顯色劑購自福州邁新公司),光鏡下控制顯色程度,行蘇木素復染;梯度酒精脫水,二甲苯透明,中性樹膠封片。陰性對照以PBS緩沖液代替一抗。
1.2.2 結果判定 染色結果由2位工作5年以上病理科醫師獨立判斷。MTA1蛋白以細胞核著色為主,部分細胞質著色,陽性顯色為棕黃色;VEGF?C蛋白以細胞質著色為主,呈棕黃色顆粒。每片隨機選取5個高倍視野,每視野≥100個細胞。①染色強度:根據染色程度,棕黃色、黃色、淺黃色、無著色分別計3分、2分、1分、0分;②陽性細胞百分比:陽性細胞數>75%、51%~75%、26%~50%、≤25%、陰性分別計4分、3分、2分、1分、0分;總積分取兩項乘積,乘積≥3分視為陽性表達。
1.2.3 隨訪 以患者治療后出院為隨訪起點,以腫瘤進展為隨訪終點,通過電話結合門診復診隨訪形式,參照實體瘤療效標準[8],統計患者無進展生存期(progression free survival,PFS)(即從治療結束到腫瘤進展時間)及3年無進展生存率。隨訪時間截至2021年9月,200例患者隨訪時長5~49個月,平均(18.66±10.32)個月,中位隨訪時間16個月。
1.3 統計學分析
應用SPSS 22.0軟件,計數數據以[n(%)]描述,采用χ2檢驗;相關性分析采用Spearman秩相關法;繪制Kaplan?Meier生存曲線,采用Log?rank檢驗比較預后。P<0.05表示差異有統計學意義。
2 結果
2.1 肺癌組織及癌旁組織MTA1、VEGF?C表達比較
MTA1在肺癌組織表達陽性率70.50%,顯著高于癌旁正常組織的26.00%(P<0.05);VEGF?C在肺癌組織表達陽性率66.00%,顯著高于癌旁正常組織的42.00%(P<0.05)。見表1。
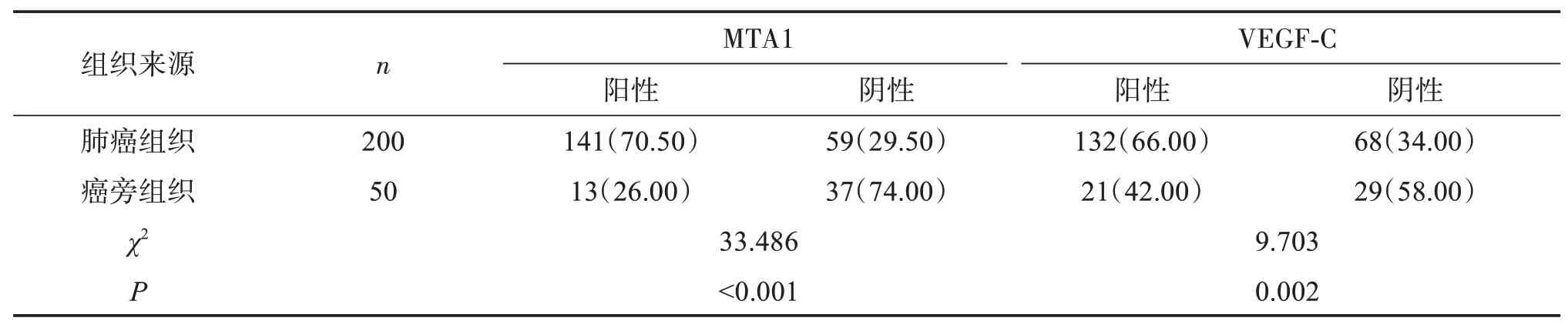
表1 肺癌組織及癌旁組織MTA1、VEGF?C表達比較Table 1 Comparison of MTA1 and VEGF?Cexpression in lung cancer tissues and adjacent tissues [n(%)]
2.2 不同病理類型肺癌組織MTA1、VEGF?C表達比較
不同病理類型肺癌組織中MTA1、VEGF?C表達差異均無統計學意義(P>0.05)。見表2。
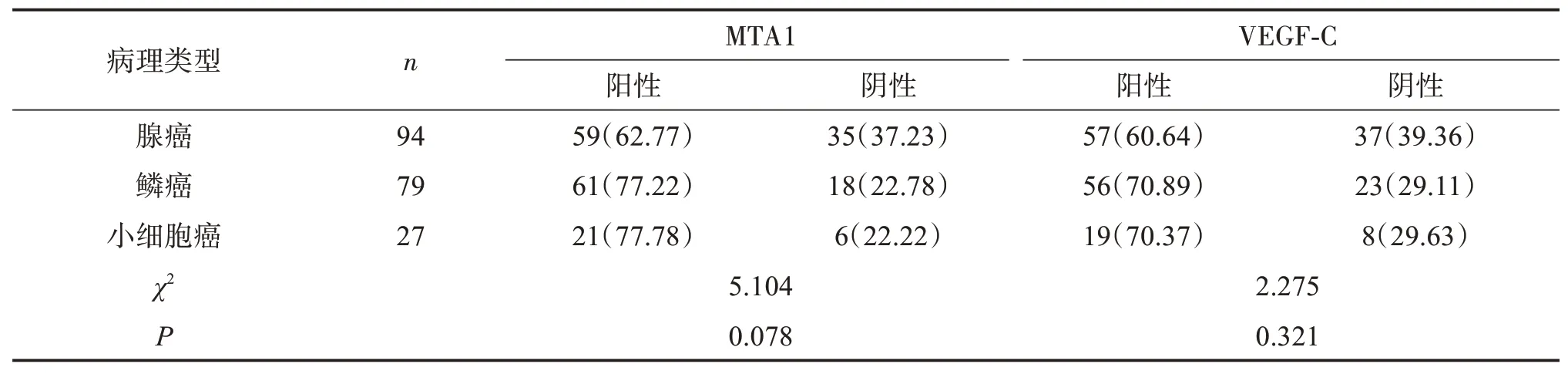
表2 不同病理類型肺癌組織MTA1、VEGF?C表達比較Table 2 Comparison of MTA1 and VEGF?Cexpression in different pathological types of lung cancer tissues [n(%)]
2.3 MTA1、VEGF?C表達與患者臨床病理特征關系
肺癌組織MTA1陽性表達與患者腫瘤分化程度低、TNM分期高、淋巴結轉移有關(均為P<0.05),與患者性別、年齡、腫瘤部位、腫瘤直徑、脈管浸潤無關(均為P>0.05);VEGF?C陽性表達與患者腫瘤分化程度低、TNM分期高、脈管浸潤、淋巴結轉移有關(均為P<0.05),與患者性別、年齡、腫瘤部位、腫瘤直徑無關(均為P>0.05)。見表3。
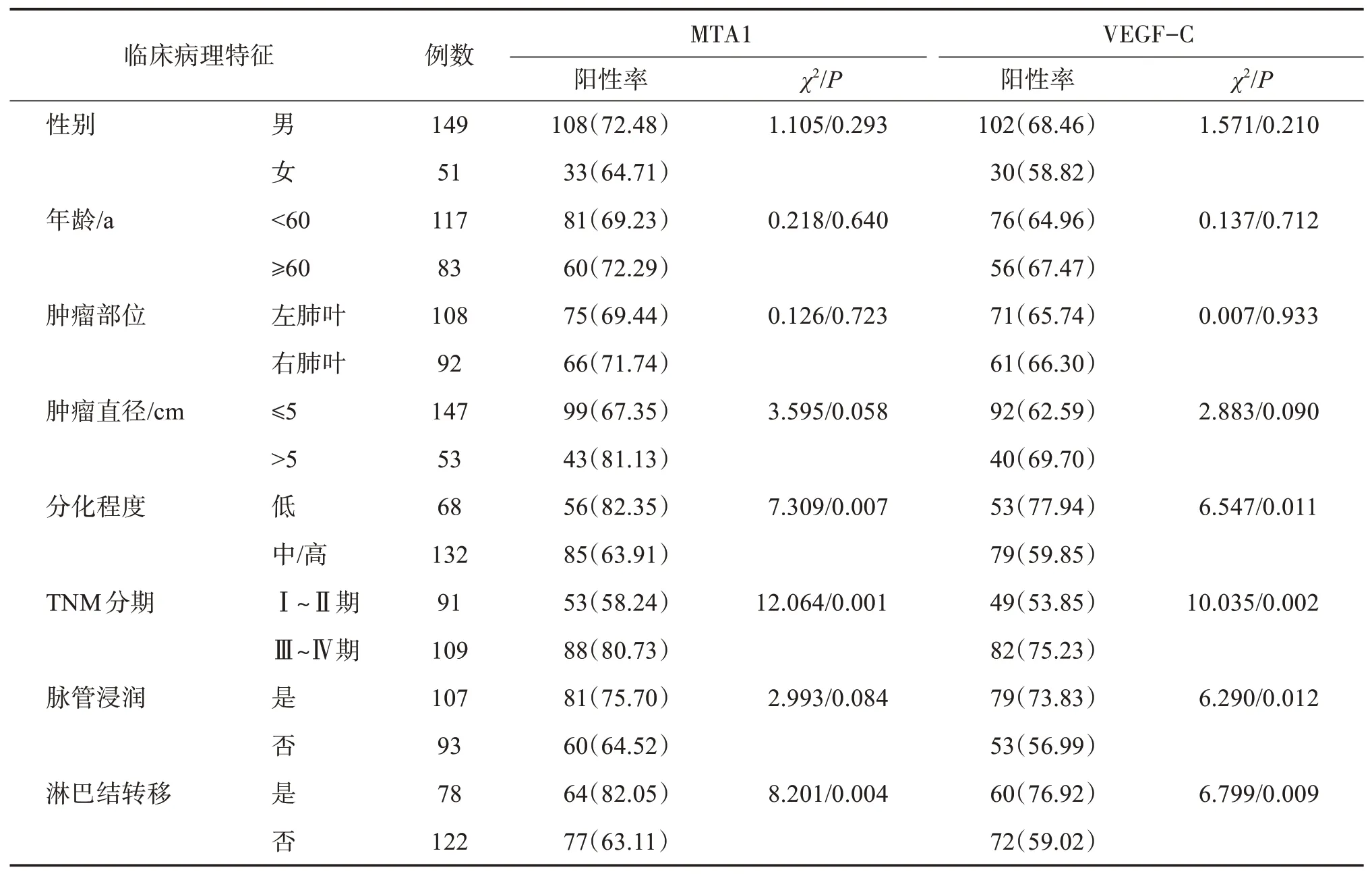
表3 MTA1、VEGF?C表達與患者臨床病理特征關系Table 3 Relationship between the expression of MTA1 and VEGF?Cand clinicopathological features of patients [n(%)]
2.4 肺癌組織MTA1、VEGF?C表達相關性
Spearman相關性分析顯示,肺癌組織MTA1與VEGF?C表達呈正相關(P<0.01)。見表4。

表4 肺癌組織MTA1、VEGF?C表達相關性Table 4 Correlation between the expression of MTA1 and VEGF?Cin lung cancer tissues
2.5 肺癌組織MTA1、VEGF?C表達與患者預后關系
隨訪時間截至2021年9月,肺癌組織MTA1陽性表達者PFS為(16.22±8.87)月,3年無進展生存率為5.67%,均低于陰性者的(24.49±11.25)月、23.73%(Log?rankχ2=22.522,P<0.001),見圖1;肺癌組織VEGF?C陽性表達者PFS為(16.99±9.58)月,3年無進展生存率為7.09%,均低于陰性者的(21.90±11.00)月、20.34%(Log?rankχ2=7.762,P=0.005),見圖2。
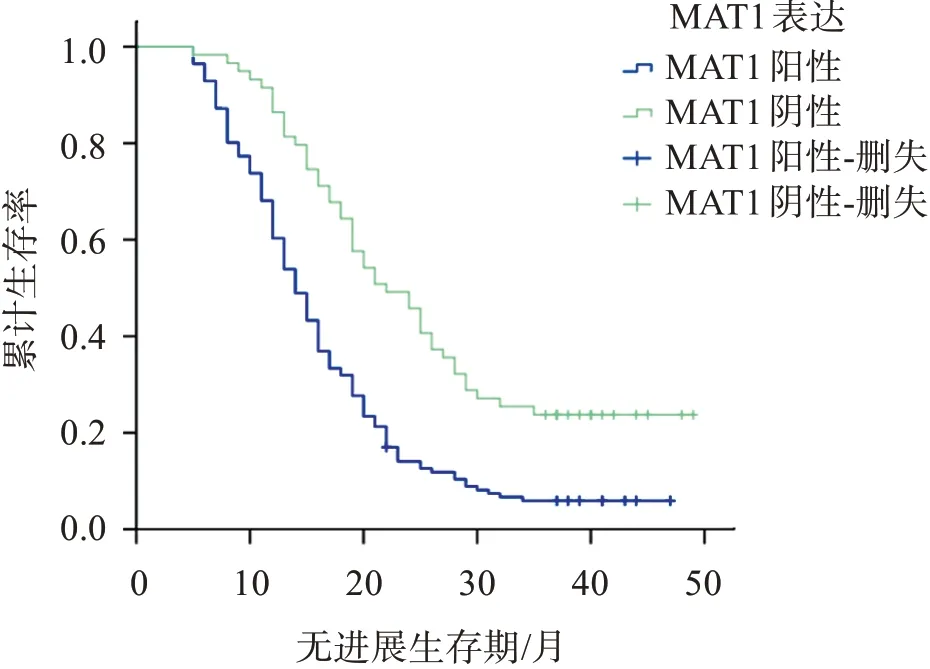
圖1 肺癌組織MTA1陽性表達與陰性表達的生存曲線Figure 1 Survival curve of positive and negative expression of MTA1 in lung cancer tissues
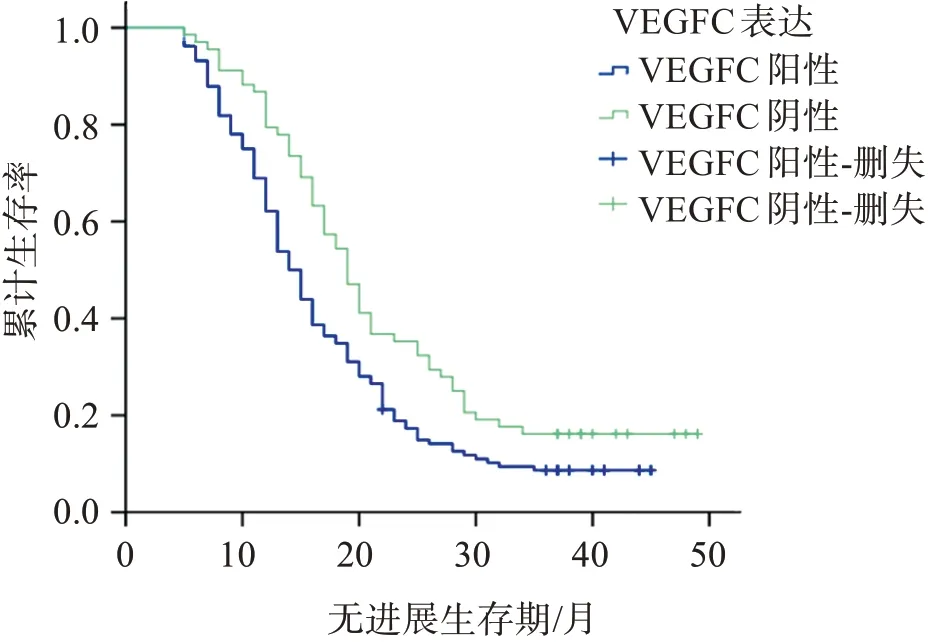
圖2 肺癌組織VEGF?C陽性表達與陰性表達的生存曲線Figure 2 Survival curve of positive and negative expression of VEGF?Cin lung cancer tissues
3 討論
腫瘤侵襲和轉移是肺癌患者死亡的主要原因之一,而惡性腫瘤的侵襲和轉移本身是一個極其復雜、高度選擇的非隨機過程,可受多基因、多分子調控,探究相關分子生物學機制是提高肺癌診斷及預防的重要前提[9]。一方面,細胞黏度、細胞外基質降解、病理性血管生成等過程是惡性腫瘤侵襲轉移的重要生物學基礎,另一方面,癌細胞的增殖、凋亡直接影響惡性腫瘤發生發展,因此圍繞相關過程探究敏感的生物標記物是肺癌研究的重要方向之一[10]。
MTA1是MTA家族重要成員,該家族能同組蛋白脫乙酰基酶1、組蛋白脫乙酰基酶2相互結合,參與核小體形成組蛋白去乙酰化復合物重塑,與染色質結構變化、核轉錄調控有關[11]。MTA1基因定位于人染色體14q32.3,編碼的MTA1蛋白羧基末端富含脯氨酸,其695~705殘基序列同SH3結構域完全配對,而后者可參與信號傳導通路中蛋白與蛋白間相互作用,與細胞內信號轉導、細胞運動、增殖等多種過程有關[12]。MTA1蛋白還具有2個酪氨酸激酶、7個酪蛋白激酶Ⅱ、9個蛋白激酶的磷酸化位點以及4個N?糖激化位點,可與信號傳導通路中維持細胞正常功能的蛋白發生作用[13]。此外,MTA1蛋白自251殘基起有一個亮氨酸反折區、5個SPXX域,自393殘基始 有 一個鋅指DNA結 合域Cys?X2?Cys,與DNA結合蛋白、基因調節蛋白有關;還有一個SANT結構域,同轉錄因子myb?相關蛋白的DNA結合域相似,可能參與細胞存活、增殖、分化等調節[14]。MTA1基因首次于老鼠的乳腺癌細胞株分離所得,其表達與乳腺腫瘤轉移呈正相關,而近些年來越來越多研究發現,MTA1蛋白高表達還與人類多種腫瘤如喉部鱗狀細胞癌、鼻咽癌等侵襲力有關[15?16]。國內有學者發現,抑制MTA1的表達,能抑制人食管癌Eca109細胞增殖,誘導細胞凋亡[17]。此外,MTA1表達并不局限于腫瘤,在正常組織器官如腦、肝、睪丸等中也可檢測出MTA1不同程度表達,其與各種癌腫的相關性報道也不盡相同[18]。本研究采用免疫組化法對肺癌組織中MTA1進行定性檢測,發現MTA1在細胞核、細胞質中均可表達,這可能與腫瘤發生發展中MTA1表達及定位紊亂有關。本文結果顯示,肺癌組織MTA1陽性表達率明顯高于癌旁正常組織,與不同病理類型無關,但與分化程度低、TNM分期高、淋巴結轉移有關,提示MTA1可能參與肺癌發生發展。
血管與淋巴管生成既是組織生長、傷口愈合的重要生理過程,也是包括腫瘤在內的多種疾病發生發展的重要病理基礎,其中血管內皮生長因子家族是血管生成主要的調控系統,也是參與淋巴管形成的重要因子[19]。VEGF?C作為血管內皮生長因子家族重要成員,是迄今發現的唯一特異性促淋巴管生長因子,一方面VEGF?C可通過與VEGFR?3結合,誘導淋巴管內皮細胞增殖、遷移,同時增加淋巴管通透性,與腫瘤淋巴管形成、癌細胞淋巴血管轉移關聯密切;另一方面VEGF?C還可通過與VEGFR?2結合,參與生理性或病理性血管生成過程啟動,與腫瘤血管生成有關。此外,VEGF?C在癌細胞脫離基質、到達轉移部位時抑制其凋亡,可促進腫瘤生長,因此被認為在腫瘤生長、轉移的不同階段均有重要作用[20?21]。在已研究的人類腫瘤中,有一半腫瘤細胞如胃癌、結腸癌、乳腺癌、前列腺癌等均可表達VEGF?C[22]。殷文娟等[23]研究指出,VEGF?C在喉鱗狀細胞癌中呈高表達,且與腫瘤淋巴管生成及淋巴結轉移有關。本文結果顯示,肺癌組織VEGF?C陽性表達率較癌旁正常組織高,與不同病理類型無關,與分化程度低、TNM分期高、脈管浸潤、淋巴結轉移有關,提示VEGF?C可能參與肺癌發生發展。本文結果還顯示,肺癌組織MTA1與VEGF?C表達呈正相關,提示二者可能在肺癌發生發展中有協同作用。有研究[24]指出,VEGF?C表達可受多個通路影響,其中包括MTA1的調控,其傳導通路為缺氧誘導因子?1α。此外,本文結果顯示,肺癌組織MTA1、VEGF?C陽性表達者PFS縮短、3年無進展生存率下降,說明MTA1、VEGF?C陽性表達與患者預后有關,二者在肺癌預后判斷中展現出一定應用價值,同時提示在臨床實踐中,對MTA1、VEGF?C陽性表達或許可通過相關靶向治療,提高抗腫瘤效果。
綜上所述,肺癌組織MTA1、VEGF?C呈異常高表達,可能與肺癌發展及預后有關,在肺癌預后評估及相關靶向治療中展現出一定應用前景。本研究不足之處在于缺乏MTA1、VEGF?C在肺癌作用中具體機制探究,且未對MTA1、VEGF?C表達進行定量檢測,可能結果不夠精準,因此尚需其他研究佐證,這也是筆者后期研究關注的方向。
猜你喜歡
保健醫苑(2023年2期)2023-03-15 09:03:04
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
中國藥學藥品知識倉庫(2022年1期)2022-03-23 04:16:57
昆明醫科大學學報(2021年4期)2021-07-23 01:21:44
天津醫科大學學報(2021年2期)2021-03-29 05:30:38
天津醫科大學學報(2019年6期)2019-08-13 07:04:26
癌變·畸變·突變(2016年3期)2016-02-27 06:15:34
醫學研究雜志(2015年12期)2015-06-10 06:57:46
中國當代醫藥(2015年7期)2015-03-01 02:01:19
鄭州大學學報(醫學版)(2015年1期)2015-02-27 14:50:26
