云南高黎貢山保山茜天然更新的分布特征1)
2021-11-03 12:12:30趙欣鳳李貴祥柴勇和麗萍邵金平
東北林業大學學報 2021年9期
趙欣鳳 李貴祥 柴勇 和麗萍 邵金平
(云南省林業和草原科學院,昆明,650201)
保山茜(Fosbergiashweliensis)為茜草科(Rubiaceae)大果茜屬(Fosbergia)常綠小喬木[1-2],原名為瑞麗茜樹(Aidiashweliensis)[3]。保山茜為云南特有,高達10 m,花期為2—7月,當年生種子到翌年成熟,未受到干擾的保山茜種球不脫落。保山茜主要分布于云南高黎貢山一帶,生長于海拔1 800~2 000 m[3]。云南省極小種群物種拯救保護規劃綱要(2010—2020年)將其列為極小種群物種。
森林天然更新是森林資源再生產的一個重要的生態學過程,也是森林得以繁衍、持續不衰和森林生態系統穩定的基礎[4-5]。森林天然更新主要受到環境因子與林分因子的影響[6],林下光照是影響樹種更新及演替的重要因素,林冠層郁閉度是決定幼苗生長發育的關鍵[7]。郭連金[8]認為香果樹幼苗受喬木蓋度、灌木蓋度、光照強度以及大氣的溫濕度影響較大;韓文娟等[9]認為凋落物是影響種子萌發的主要因素;王晗生[10]認為水分因子對人工林天然更新起著重要作用。許寧等[11]研究發現林窗或林緣更適合于水青樹的自然更新;楊清培等[12]認為群落結構與物種組成是影響南方鐵杉更新能力的主要因素;李榮等[13]認為近自然經營間伐措施能夠促進遼東櫟次生林的林木更新;影響林分天然更新的因子眾多,森林類型或樹種不同,影響林分天然更新的關鍵因子也不同[6,14],對于相同的林分其立地的差異也會導致其更新不一致[15]。
對保山茜的研究主要集中在該物種的確認[1]、染色體核型[2]、快速繁殖和離體保存[16]、物種名錄[17]、微衛星位點標記[18]、干燥耐受力和發芽潛力[19]以及葉綠體基因組系列[20]等方面,有關保山茜天然更新的研究未見報道,本文通過對保山茜更新能力及空間格局、更新苗分布狀況、距母樹不同距離、不同方位的天然更新狀況的調查分析,從而為保山茜種群保護和擴繁提供技術支撐。
1 研究區概況
研究區位于以云南高黎貢山南段(地處云南省保山市隆陽區的赧亢村、騰沖市的小地方村和龍陵縣的小黑山交匯地段)保山茜較為集中的分布區域。地勢東高西低,為我國西南部亞熱帶高原季風氣候,海拔2 000~2 300 m。全年盛行西南風,干濕季分明,日氣溫溫差大,兼有大陸性和海洋性氣候特征。年均氣溫15 ℃,最冷月為1月,氣溫為7 ℃左右,年降水量1 200 mm左右。11月下旬至翌年4月為干季,日照充足,降水少,相對濕度50%~60%,5—10月為濕季,降水量占全年的87%,相對濕度比干季高20%左右。研究區土壤為黃棕壤、棕壤,主要植被類型為中山濕性常綠闊葉林。
2 研究方法
2.1 調查方法
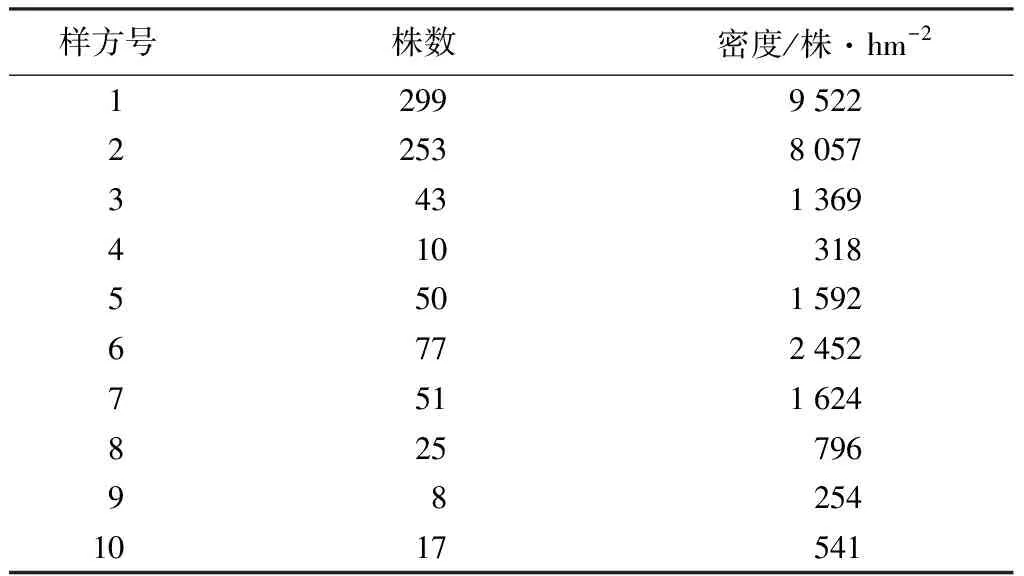
在保山茜分布的區域內,通過充分踏查,隨機選擇已結實的10株保山茜作為母樹,所選母樹基本情況見表1,分別記錄母樹的位置、海拔、郁閉度、胸徑、樹高和冠幅等。

表1 保山茜天然更新調查母樹基本情況
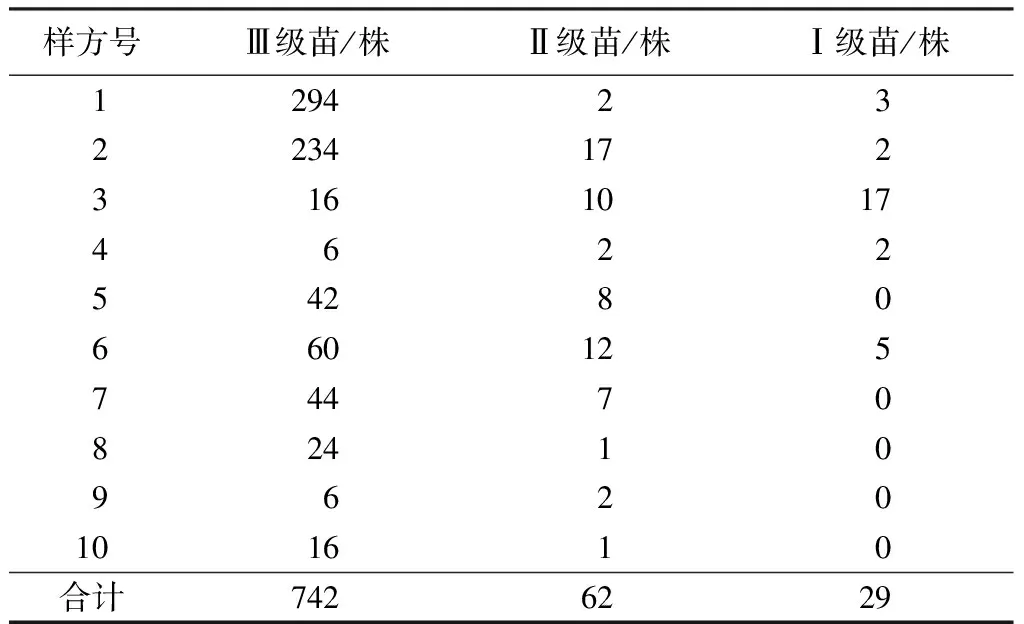
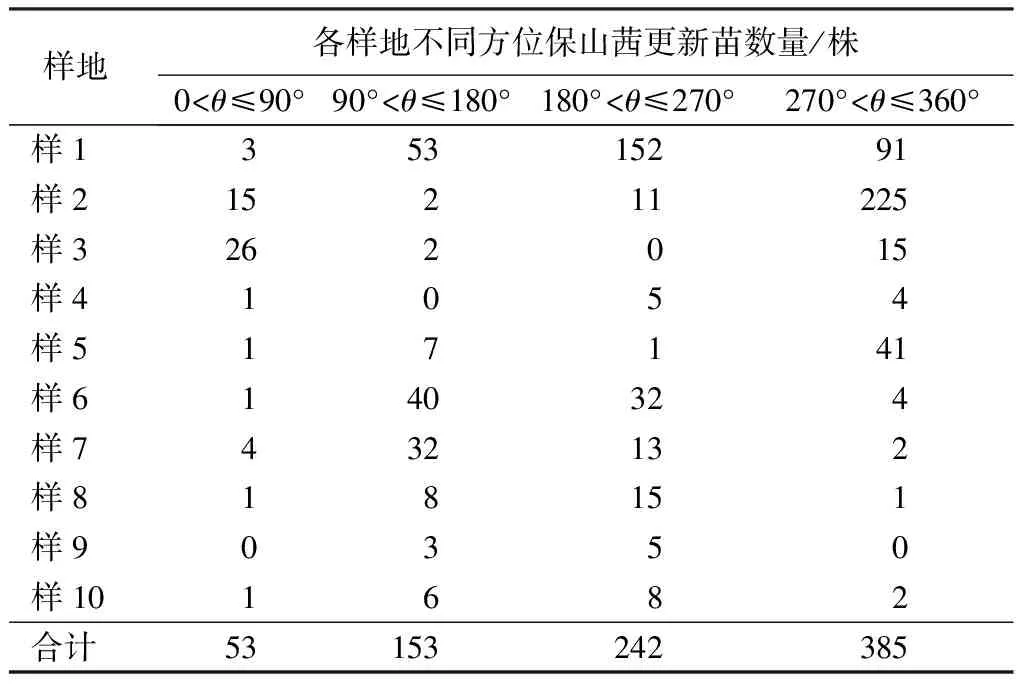
以母樹為中心向外輻射,按半徑(d)0 密度計算公式:D=x/s。式中x為樣圓或樣帶內保山茜的個體數;s為樣圓或樣帶面積;D為密度。 種群分布格局采用方差與平均數的比率確定,即S2/m[21-22],m=(∑fx)/n;S2=∑(x-m)2/(n-1)。式中x為樣方中某個種的個體數;m為各樣方中某個種個體平均數;n為取樣個體數;S2為分散度(方差)。 年齡結構是種群動態的重要特征[23],一般采用高度和徑級大小結構代替年齡結構的方法來分析其結構特征[8,11,23-26],但大小結構沒有統一的劃分標準,大多按植物特征和實際需要進行劃分。因此,保山茜苗木分級按天然更新的實際情況并參照袁士云等[25]的劃分方法,將保山茜天然更新苗高(H)劃為Ⅰ級苗(H≥50 cm)、Ⅱ級苗(30 cm≤H<50 cm)和Ⅲ級苗(H<30 cm),母樹作為大樹不計入更新分析。 采用Microsoft Excel 97-2003對保山茜更新苗木進行統計整理,并在不同方位角和不同距離進行方差分析。 由表2可知,通過對研究區保山茜10株母樹更新調查統計,保山茜的更新主要在以母樹為中心的10 m半徑范圍內,在314 m2的樣圓內更新株數為8~299株,10個樣圓內共有833株;保山茜種群密度為254~9 522株/hm2,平均密度為2 652株/hm2。 表2 保山茜在各樣圓中的更新數量 組成種群的個體在其生活空間中的位置狀態或布局分為均勻型、隨機型和集聚型[22],通過對保山茜種群內分布型的指標方差和平均數的比率(S2/m)計算,得到S2/m=130.69,即S2/m顯著大于1,說明保山茜在調查樣圓內為集聚型分布,表明保山茜種群個體分布不均勻,幼樹的分布為集聚型。 由表3可知,在10個樣圓分布的843株保山茜植株總數中,母樹10株,更新苗833株,其中Ⅲ級苗有742株,占更新株數的89.08%;Ⅱ級苗有62株,占更新株數的7.44%;Ⅰ級苗29株,占更新株數的3.48%。從Ⅰ級苗、Ⅱ級苗和Ⅲ級苗更新數量來看,隨著苗木齡級增長,大量的更新苗受種內競爭與環境因素的影響,數量急劇減少,而且所調查樣圓內保山茜更新的苗木樹高均小于1.3 m,說明保山茜雖然更新幼苗較多,但在母樹下沒有苗木長成小樹或大樹。 表3 保山茜不同齡級(苗高)的更新狀況 通過對保山茜不同齡級的更新分布株數進行方差分析,得到F=4.597 822,P=0.019 123<0.05,表明不同齡級的保山茜更新分布株數差異顯著。 由表4可知,保山茜在方位角0<θ≤90°、90°<θ≤180°、180°<θ≤270°、270°<θ≤360°的更新株數分別為53、153、242、385株,所占比例分別為6.36%、18.37%、29.05%和46.22%。從不同方位角的更新狀況來看,不同方位角范圍內,保山茜更新具有一定差異,但通過對保山茜不同方位角更新分布株數進行方差分析,得到F=1.037 789,P=0.387 52>0.05,表明不同方位角的保山茜更新分布株數差異不顯著。 由表5可知,保山茜距離母樹不同范圍的更新情況,在0 表4 保山茜不同方位角上的天然更新狀況 表5 保山茜不同距離范圍內的天然更新狀況 保山茜為云南特有[27],其分布較少,刀志林等[18]研究認為保山茜野外分布僅在100 km2的范圍內,數量不超過200株的成年單株,處于瀕臨滅絕狀態,云南省將其列為極小種群物種。因此,其天然更新和解瀕技術就更加受到關注。在更新過程中,任何一種因素都可能改變整個群落的動態[28],使一些種群自然結實率低,難以獲得種子,一些種群不能結實[29],從而影響種群的更新。通過項目組多年的觀察,保山茜天然更新受自身特點的影響較大,保山茜成年母樹大多能夠結實,結實率在90%以上,種子聚集在結實種球內,結實種球不易脫落,在受到風、鼠等自然或人為干擾才會脫落。保山茜種子在野外的發芽情況較好,但由于保山茜種子集中在各個種球內,在各個種球內的種子呈叢壯發芽,范圍基本就是種球的投影面積,種子發芽較為集中,不易擴散。從保山茜的更新研究來看,保山茜更新主要集中在以母樹為中心的10 m半徑范圍內,更新密度在254~9 522株/hm2,種群內分布格局(S2/m)為130.69,顯著大于1,說明保山茜天然更新在調查樣圓內為集聚型分布,種群個體分布不均勻,與種子較為集中的特點是一致的,導致了保山茜種群擴大較為困難。 天然更新直接影響植物種群的數量和分布格局[30],保山茜在10個樣圓中天然更新的株數是843株,Ⅲ級苗有742株,占更新株數的89.08%;Ⅱ級苗有62株,占更新株數的7.44%;Ⅰ級苗有29株,占更新株數的3.48%。保山茜在不同齡級的更新分布株數差異顯著(P<0.05),表明保山茜雖然更新幼苗較多,但隨著苗木齡級增長,數量急劇減少,能長成小樹或大樹的苗木不多。在方位角0<θ≤90°、90°<θ≤180°、180°<θ≤270°、270°<θ≤360°的更新株數所占比例分別為6.36%、18.37%、29.05%和46.22%,不同方位角保山茜更新分布株數差異不顯著(P>0.05)。同種個體密度較高或者距離母樹較近的地方,物種的種子、幼苗以及幼樹的存活率較低[28]。從離母樹不同距離范圍的保山茜更新情況可看出,離母樹2 m2.2 種群數量及分布格局
2.3 年齡結構劃分
3 結果與分析
3.1 保山茜的天然更新能力及空間格局
3.2 保山茜更新苗年齡分布特征
3.3 保山茜在不同方位角上的更新狀況
3.4 保山茜在不同距離的更新狀況

4 結論與討論
