干露對克氏原螯蝦成蝦存活、相關代謝酶及組織結構的影響
2021-10-23 02:28:56王源源成永旭李晨露劉明明奚業文朱傳忠李嘉堯
南方水產科學 2021年5期
王源源,成永旭,李晨露,劉明明,奚業文,朱傳忠,李嘉堯
(1. 上海海洋大學/農業農村部淡水種質資源重點實驗室/水產科學國家級實驗教學示范中心/上海水產養殖工程技術研究中心,上海 201306; 2. 安徽省水產技術推廣總站,安徽 合肥 230000;3. 大北農水產科技集團股份有限公司,福建 詔安 363500)
克氏原螯蝦 (Procambarus clarkii),俗稱小龍蝦,屬于淡水螯蝦[1]。2019年中國克氏原螯蝦養殖總產量達208.96×104t,成為產量第一的淡水養殖經濟蝦類[2]。克氏原螯蝦在養殖過程中,可離水存活較長時間,故在其流通過程中多采用干法運輸[3]。
干法運輸時水產動物常遭受干露脅迫,干露脅迫主要包括失水脅迫和低氧脅迫[4]。失水脅迫使機體水分喪失,打破細胞內外水平衡,造成組織結構損傷;低氧脅迫下機體氧供應不足,制約有氧呼吸,最終導致生理代謝失衡[5-6]。已有研究表明,干露脅迫會影響脊尾白蝦[7](Exopalaemon carinicauda) 的呼吸代謝和日本囊對蝦[8](Marsupenaeus japonicus) 的抗氧化應激,降低三疣梭子蟹[9](Portunus trituberculatus) 的成活率。克氏原螯蝦幼蝦免疫系統尚未發育完全,體質弱、抗逆性差,在進行干法運輸時成活率低,影響經濟效益,有些地區專以運輸成蝦的方法獲得親本,通過繁育獲得蝦苗以保證當地的規模養殖[10]。
目前關于干露對克氏原螯蝦成蝦影響的研究尚未見報道,本研究采用溫度為 (16±1) ℃、相對濕度 (Relative humidity, RH) 為 (55±5)% 的干露條件對雌、雄成蝦進行不同時間的干露脅迫,通過記錄成蝦死亡時間、測定相關代謝酶、H.E染色的方法,研究了干露對克氏原螯蝦成蝦的存活、相關代謝酶和組織結構的影響,為調整其運輸時間,改善其運輸方式,提高其運輸成活率提供參考依據。
1 材料與方法
1.1 實驗材料
實驗時間為2019年9月,克氏原螯蝦取自上海海洋大學崇明基地,選擇附肢健全、活力旺盛的雌、雄蝦各90尾,將雌、雄各30尾分別暫養于單個循環水槽 (長 293 cm×寬 80 cm×高 20 cm,水深15 cm) 7 d。養殖用水為經充分曝氣的自來水,水溫為 (23.0±1.0) ℃,pH 為 7.5±0.5,持續充氧以保證水中溶氧充足。在養殖水桶中放入適量假水草,供克氏原螯蝦棲息,且減少個體之間互相打斗。每天17時用虹吸管吸出殘餌與糞便,并換水1/3以確保養殖水質清潔,投喂蝦體質量3%~5%的配合飼料 (大北農水產科技集團股份有限公司)。實驗前 24 h 停止投喂;雄蝦體質量為 (44.4±6.5) g,體長為 (79.2±3.6) mm;雌蝦體質量為 (41.9±5.9) g,體長為 (82.5±3.4) mm。
1.2 實驗方法
實驗在崇明基地實驗室內進行,提前24 h調節加濕器 (廣東容聲電器,PH-88) 和除濕機(GEDRYC20C),并通過溫濕度記錄儀 (精創RC-4HC) 顯示情況,控制房間溫度為 (16±1) ℃,RH為 (55±5)%。
1.2.1 預實驗 取雌、雄蝦各 8尾,單尾分別裝于1 000 mL的聚丙烯 (PP) 盒 (14.9 cm×6.2 cm×15.0 cm),觀察并統計克氏原螯蝦死亡時間。根據死亡率與時間進行回歸分析,建立回歸方程,根據方程求得干露條件下的半致死時間 (LT50)。
1.2.2 正式實驗 雌、雄蝦各 60尾,干露前擦干體表水分后稱質量,即為初始濕體質量 (WO),于上述溫濕度條件下進行干露脅迫,每6 h觀察蝦體存活情況,以晃動PP塑料盒蝦體包括觸角無反應判定蝦體死亡,分別在第0、第3、第6、第12、第24、第36、第48、第60、第72、第96、第132和第144小時隨機挑選雌、雄蝦各4尾進行采樣。
1.2.3 樣品采集與處理 采樣時將克氏原螯蝦體表水分擦干后稱質量 (WT),計算其體質量消耗率后 [體質量消耗率=(WO?WT)/WO×100%],置于解剖盤上,剪開頭胸甲和腹部,取出肝胰腺和肌肉,解剖盤上將其快速切割成約0.2 g的小塊,分裝于0.2 mL離心管中,液氮速凍后放入?80 ℃冰箱中保存用于酶活力測定。另取適量肝胰腺、鰓和肌肉組織于Bouin's液中固定,用于后續組織切片的制作與觀察。
1.2.4 酶活力測定 樣品按質量體積比 1∶9加入0.9%生理鹽水,冰上研磨將其制成10%的組織勻漿,離心取上清液待測。乳酸脫氫酶 (LDH) 活力采用蘇州科銘生物技術有限公司的試劑盒進行測定,琥珀酸脫氫酶 (SDH) 活力和乳酸 (LA) 濃度采用南京建成生物工程研究所試劑盒測定,具體方法參照試劑盒說明書操作。LDH酶活的單位定義為每克組織每分鐘催化產生1 nmol丙酮酸定義為1個酶活力 (U·g?1)。SDH酶活的單位定義為每毫克蛋白每分鐘使反應體系的吸光度降低0.01為1個比活性單位。
1.2.5 組織切片 Bouin's液固定 24 h 后換70%乙醇并進行乙醇梯度脫水,二甲苯透明,石蠟包埋,切片機 (LEICA RM2125 RTS) 5~7 μm 切片,蘇木精-伊紅染色,中性樹膠進行封片,研究級顯微鏡 (ECLIPES 80i) 觀察并拍照。
1.3 數據分析
采用SPSS 19.0軟件進行數據分析,實驗結果以“平均值±標準差 ()”表示,不同干露時間點數據運用單因素方差分析 (One-way ANOVA)進行比較,差異顯著時采用Duncan's多重比較,P<0.05認為差異顯著。
2 結果
2.1 預實驗結果分析
預實驗雌、雄克氏原螯蝦死亡情況見圖1。實驗開始第75小時雄蝦開始出現死亡,在干露258 h后全部死亡;雌蝦第82小時開始死亡,在干露272 h后全部死亡。將雌、雄蝦死亡率與時間進行回歸分析,分別得到干露時間與累積死亡率的直線關系,線性顯著相關。由線性回歸方程可以得到該干露脅迫條件下克氏原螯蝦雄蝦的LT50為148.36 h,雌蝦的 LT50為 144.01 h。
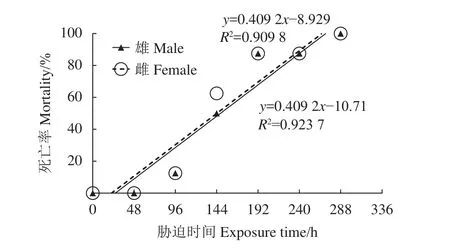
圖1 干露脅迫下克氏原螯蝦的死亡率Figure 1 Mortality of P. clarkii in air exposure periods
2.2 克氏原螯蝦的體質量消耗率分析
克氏原螯蝦成蝦的體質量消耗率隨干露時間的延長逐漸增大 (圖2)。干露前期雌、雄蝦體質量消耗率無顯著差異,雄蝦體質量消耗率在干露第72和第132小時出現顯著升高 (P<0.05),雌蝦體質量消耗率在干露第36、第60和第72小時出現顯著升高 (P<0.05),干露48 h后,在同一干露時間雌蝦的體質量消耗率均高于雄蝦。
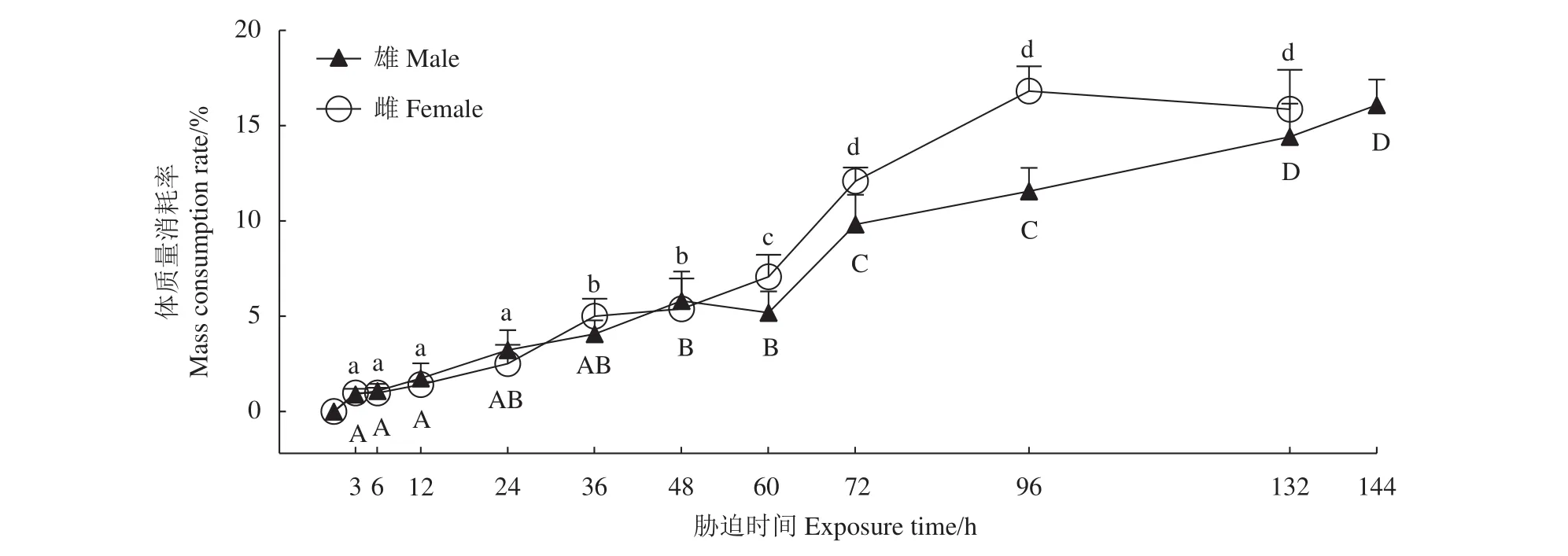
圖2 干露脅迫下克氏原螯蝦的體質量消耗率大寫字母不同表示雄蝦不同脅迫時間組的差異顯著 (P<0.05),小寫字母不同表示雌蝦不同脅迫時間組的差異顯著 (P<0.05)。Figure 2 Mass consumption rate of P. clarkii in air exposure periodsValues with different uppercase and lowercase letters indicate significantly difference in male and female crayfish among different groups, respectively (P<0.05).
2.3 克氏原螯蝦LDH活力的變化
2.3.1 干露脅迫對肝胰腺 LDH 活力的影響 雄蝦肝胰腺LDH活力在第6、第12、第24和第36小時呈逐級遞增趨勢 (圖3-a),在第36小時達到最大(204.16 U·g?1),在隨后的第 48、第 60和第 72小時 LDH 活力仍顯著高于 0 h 組 (P<0.05),但相比前期脅迫稍有降低。在第96、第132和第144小時雄蝦LDH活力無明顯變化 (P>0.05)。雌蝦肝胰腺LDH活力在整個干露時間段內呈現先上升后下降的趨勢 (圖3-b),在干露第3和第6小時LDH活力顯著降低 (P<0.05),在第12、第24和第48小時LDH相比0 h組有所升高 (P<0.05),在第24小時達到峰值 (154.17 U·g?1)。干露第 36小時無顯著差異 (P>0.05)。在干露后期的第60、第72和第96小時雌蝦肝胰腺LDH顯著低于干露0 h組(P<0.05)。
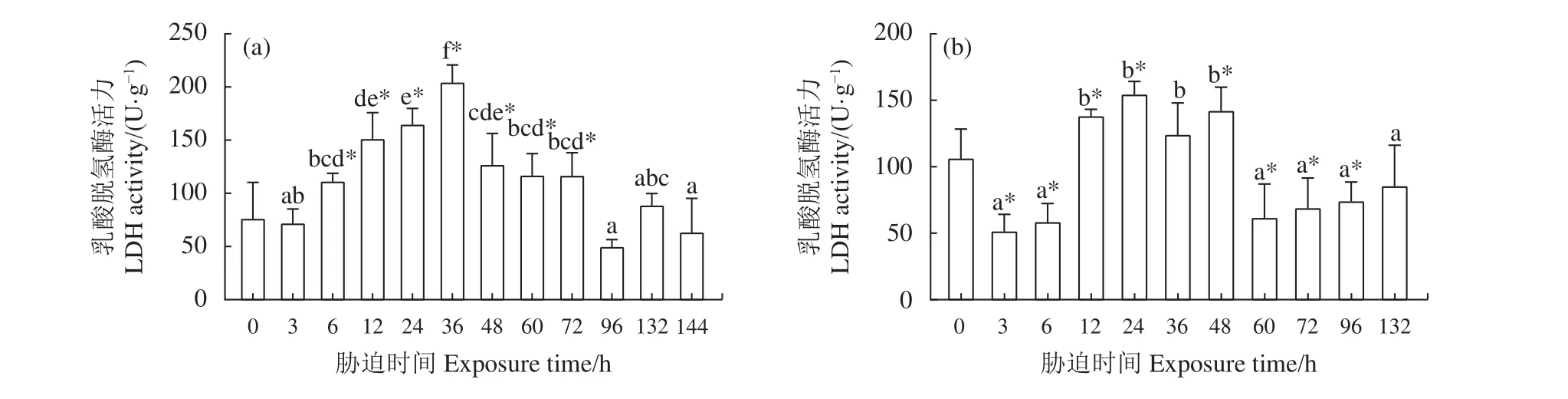
圖3 干露脅迫對克氏原螯蝦雄蝦 (a) 和雌蝦 (b) 肝胰腺乳酸脫氫酶活力的影響*. 各組與0 h組的差異顯著 (P<0.05);不同字母上標表示除0 h組外不同脅迫時間組的差異顯著 (P<0.05);后圖同此。Figure 3 Effect of dry exposure on hepatopancreas LDH activity of male (a) and female (b) P. clarkii*. Significant difference between each group and 0 h group (P<0.05); different letters represent significant difference among different groups except 0 h group (P<0.05). The same case in the following figures.
2.3.2 干露脅迫對肌肉 LDH 活力的影響 雄蝦肌肉LDH活力見圖4-a,在干露24、第36、第48和第60小時顯著高于0 h組,在第48小時達到峰值 (120.22 U·g?1),隨后 LDH 活力與 0 h 組無顯著差異 (P>0.05)。隨著干露時間的延長,雄蝦肌肉LDH活力整體呈現先升高后下降的趨勢。干露期間雌蝦肌肉LDH活力先升高后下降 (圖4-b),在第 132小時達到最低 (40.88 U·g?1)。雌蝦肌肉LDH活力在第6、第12、第24和第36小時顯著高于0 h組 (P<0.05),在第36小時達到最大活力值 (104.63 U·g?1),干露 48~96 h 階段 LDH 活力與干露 0 h 組無顯著差異 (P>0.05)。
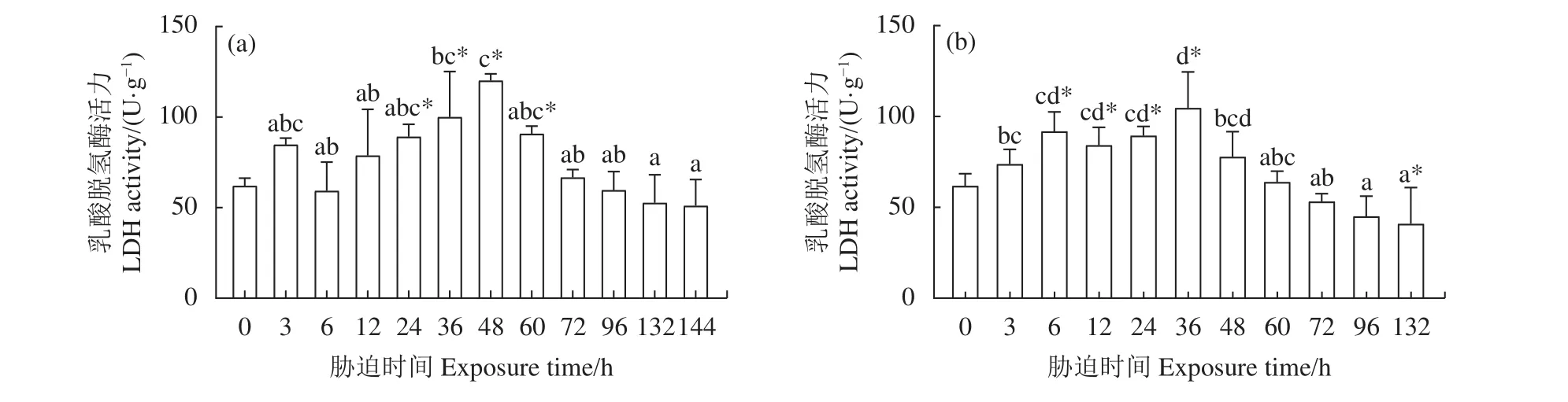
圖4 干露脅迫對克氏原螯蝦雄蝦 (a) 和雌蝦 (b) 肌肉乳酸脫氫酶活力的影響Figure 4 Effect of dry exposure on muscle LDH activity of male (a) and female (b) P. clarkii
2.4 克氏原螯蝦SDH活力變化
2.4.1 干露脅迫對肝胰腺 SDH 活力的影響 雄蝦肝胰腺SDH活力在干露期間呈現先下降后升高趨勢 (圖5-a),在干露第48小時達到最低 (4.36 U·mg?1),第 60小時達到峰值 (11.65 U·mg?1)。而雌蝦肝胰腺SDH活力在干露前期梯度遞減 (圖5-b),第 36小時達到最低 (1.97 U·mg?1),在第 48和第60小時明顯激增,但與0 h組無顯著差異 (P>0.05),在干露 60 h 后活力顯著降低 (P<0.05)。

圖5 干露脅迫對克氏原螯蝦雄蝦 (a) 和雌蝦 (b) 肝胰腺琥珀酸脫氫酶活力的影響Figure 5 Effect of dry exposure on hepatopancreas SDH activity of male (a) and female (b) P. clarkii
2.4.2 干露脅迫對肌肉 SDH 活力的影響 雄蝦肌肉SDH活力見圖6-a,在干露第24和第60小時明顯降低 (P<0.05),在第36小時達到最大 (9.76 U·mg?1)。雌蝦肌肉SDH活力在第12小時達到最低 (7.18 U·mg?1),第 36小時后隨干露時間的延長顯著降低 (P<0.05,圖 6-b)。

圖6 干露脅迫對克氏原螯蝦雄蝦 (a) 和雌蝦 (b) 肌肉琥珀酸脫氫酶活力的影響Figure 6 Effect of dry exposure on muscle SDH activity of male (a) and female (b) P. clarkii
2.5 克氏原螯蝦肌肉LA濃度變化
雌、雄蝦肌肉LA水平隨干露時間的延長呈現先升高后下降的變化 (圖7),其中雄蝦LA水平在第 48小時達到最大值 (732.33 μmol·g?1),雌蝦在第 60小時達到峰值 (706.01 μmol·g?1)。

圖7 干露脅迫對克氏原螯蝦雄蝦 (a) 和雌蝦 (b) 肌肉乳酸質量摩爾濃度的影響Figure 7 Effect of dry exposure on muscle LA molar concentrations of male (a) and female (b) P. clarkii
2.6 干露脅迫對克氏原螯蝦組織結構的影響
2.6.1 干露脅迫對肝胰腺的影響 干露 0 h 組雌、雄蝦肝胰腺肝小管排列緊密,細胞結構正常,細胞界限清晰 (圖8和圖9)。隨著干露時間的延長,雄蝦肝小管形態有明顯膨脹,管壁外側基膜在第72小時開始有離散現象,在第96小時更為明顯;管腔內壁基膜逐漸擴張,形狀呈不規則多邊形,尤其在第36小時之后明顯變大;肝小管內部細胞收縮,B細胞以及內部運轉泡體積增大,數量增多,雌蝦此現象較雄蝦明顯。雌蝦肝小管在持續干露時間段內除第132小時外始終緊密排列,內部基膜擴張明顯,管腔隨干露時間逐漸變大,在第132小時達到最大;內部細胞在第48、第96和第132小時結構模糊,細胞界限不清晰。隨著干露時間的增加,雌、雄蝦柱狀上皮細胞的細胞質內均出現許多空泡,空泡隨干露時間數量增多、體積增大,干露終期細胞空泡化嚴重。雌、雄蝦干露期間肝胰腺R細胞數量無明顯變化。
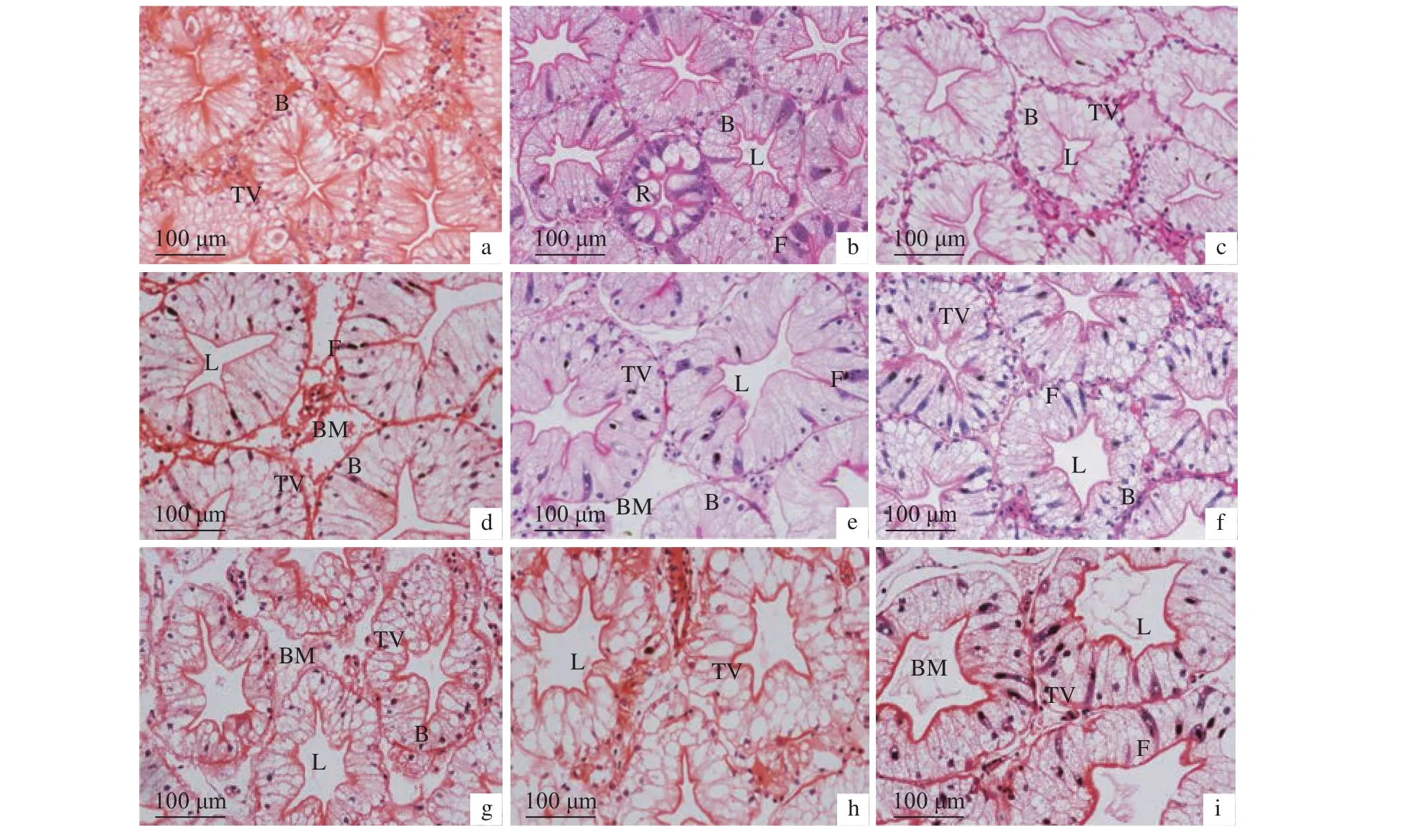
圖8 干露脅迫對雄性克氏原螯蝦肝胰腺顯微結構的影響a—i分別代表干露脅迫 0、12、24、36、48、60、72、96 和 132 h;B. B 細胞;R. R 細胞;F. F 細胞;L. 管腔;BM. 基膜;TV. 轉運泡;圖 9同此。Figure 8 Effect of dry exposure on hepatopancreas microstructure of male P. clarkiia–i represent dry exposure of 0, 12, 24, 36, 48, 60, 72, 96 and 132 h, respectively; B. B cell; R. R cell; F. F cell; L. Lumen;BM. Basement membrane; TV. Transferred vacuoles; the same case in Figure 9.

圖9 干露脅迫對雌性克氏原螯蝦肝胰腺顯微結構的影響Figure 9 Effect of dry exposure on hepatopancreas microstructure of female P. clarkii
2.6.2 干露脅迫對鰓的影響 干露 0 h 組雌、雄蝦鰓葉結構完整,鰓膜薄且平整光滑、形狀規則整齊(圖10和圖11),正常內部呼吸上皮細胞單層逐個相連圍成微血管腔,微血管腔內有血細胞和血淋巴液。在干露第12小時出現雄蝦鰓膜邊緣結構模糊,雌蝦鰓絲輕微腫大現象。在隨后的干露時間,鰓膜皆出現褶皺彎曲,表面有嚴重粗糙感,尤其在干露第24和第36小時上皮細胞有部分脫落現象,隨著干露時間延長,雌、雄蝦鰓組織上皮細胞脫落、壞死愈發嚴重。干露后期的第96和第132小時鰓細胞數量減少,鰓膜受損嚴重,微血腔中的血淋巴液彌散嚴重,基本失去正常鰓細胞結構。

圖10 干露脅迫對雄性克氏原螯蝦鰓顯微結構的影響GM. 鰓膜;H. 血細胞;CL. 微血管腔;REC. 呼吸上皮細胞;圖 11同此。Figure 10 Effect of dry exposure on gill microstructure of male P. clarkiiGM. Gill membranes; H. Hemocytes; CL. Capillary lumen; REC. Respiratory epithelium cells; the same case in Figure 11.

圖11 干露脅迫對雌性克氏原螯蝦鰓顯微結構的影響Figure 11 Effect of dry exposure on gill microstructure of female P. clarkii
2.6.3 干露脅迫對肌肉的影響 肌肉組織變化見圖12和圖13,干露0 h組肌肉組織結構排列較為整齊,肌纖維間隙緊致、平整,雄蝦在干露第12、第36、第60、第72和第96小時,雌蝦在干露第24、第36、第48、第60、第96和第132小時組織肌纖維彎曲呈波浪狀,間隙褶皺疏松。
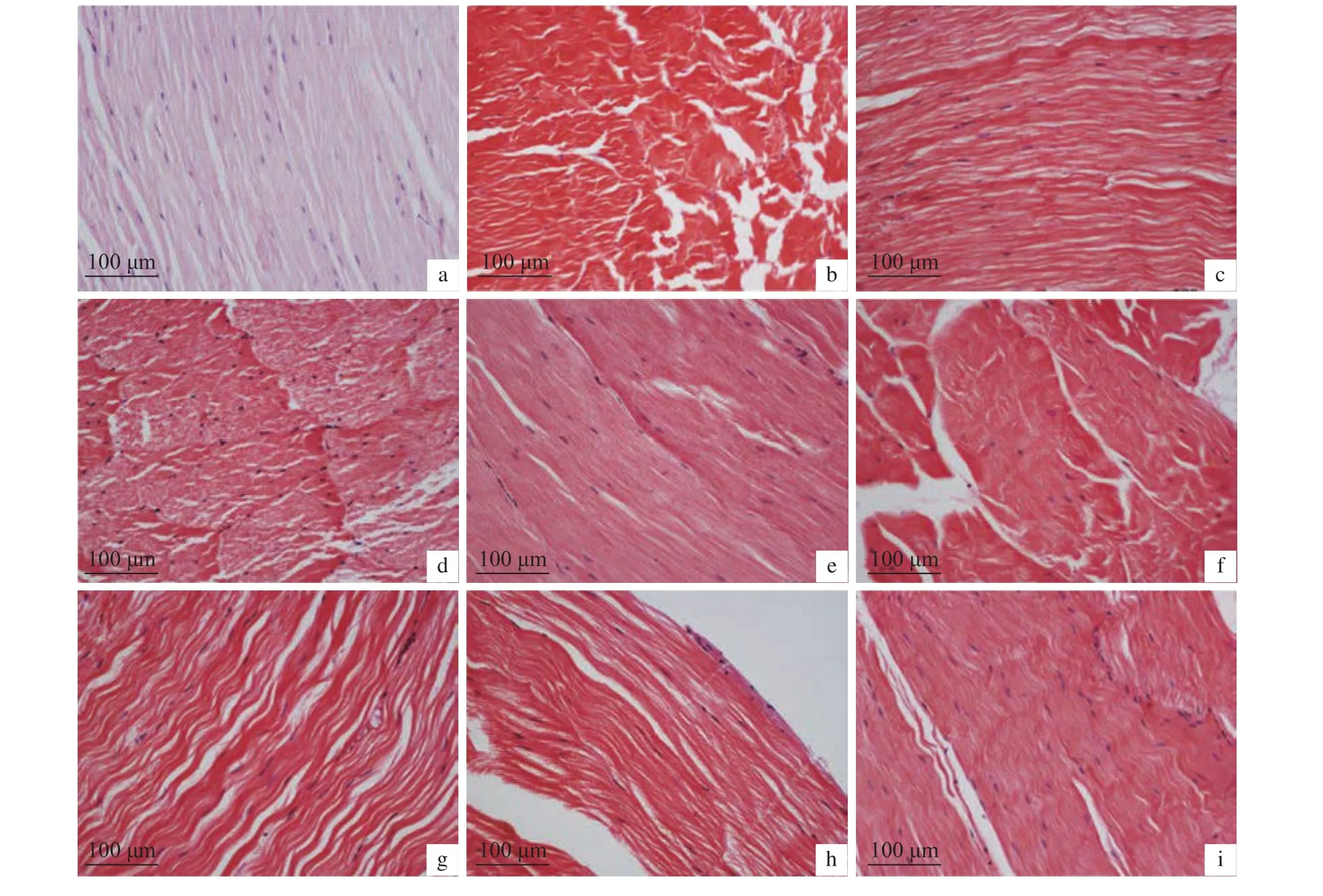
圖12 干露脅迫對雄性克氏原螯蝦肌肉顯微結構的影響Figure 12 Effect of dry exposure on muscle microstructure of male P. clarkii
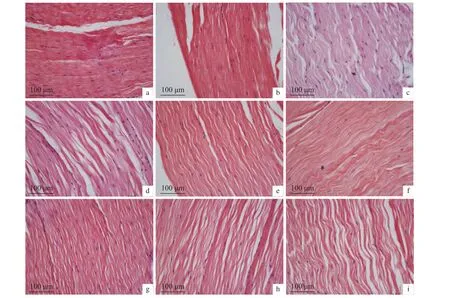
圖13 干露脅迫對雌性克氏原螯蝦肌肉顯微結構的影響Figure 13 Effect of dry exposure on muscle microstructure of female P. clarkii
3 討論
3.1 克氏原螯蝦成蝦的耐干露能力
半致死時間能反映生物對脅迫因子的耐受能力[11]。克氏原螯蝦成體在水中的窒息點低至0.061 mg·L?1[12],在本研究 (16±1) ℃、RH (55±5)% 干露脅迫條件下,雄蝦LT50為148 h,雌蝦為144 h。三疣梭子蟹[9]幼蟹在15 ℃干露4 h后全部死亡;未抱卵天津厚蟹[13](Helice tientsinensis) 在 RH 50%~60% 下5 d后全部死亡;克氏原螯蝦[14]幼蝦在20 ℃、RH 50% 干露 24 h 后成活率為 53.3%。相比來說,克氏原螯蝦成蝦對干露環境的耐受能力較好。機體的耐干露時間與自身的保水能力有關[15-16],克氏原螯蝦具有獨特的羽毛狀鰓絲,在擴大與水體接觸面積的同時也能保留大部分水分[17],從而能夠在失水環境中存活一段時間。另外,干露耐受時間與自身水分喪失速度有關[15-16],體質量消耗率可反映機體的水分喪失,雄、雌蝦體質量消耗率分別在第60和第48小時后急劇增加,第48小時后雌蝦體質量消耗率皆高于雄蝦,因而其干露耐受時間小于雄蝦。雌蝦體質量消耗率急劇升高的時間早于雄蝦的原因還有待進一步研究。
3.2 干露脅迫對克氏原螯蝦呼吸代謝酶的影響
本研究干露初期第3和第6小時,雌、雄蝦肝胰腺、肌肉的LDH活力及LA濃度與0 h組相比變化不顯著,機體脫離水體剛進入干露環境,鰓部仍留有一定水分,以維持正常的呼吸代謝[18]。隨著干露時間的延長,在干露12~48 h 階段,雌、雄蝦的肝胰腺和肌肉LDH活力呈上升趨勢,LA濃度變化趨勢與LDH相同。LDH作為機體無氧代謝的標志酶[19],陶易凡等[20]在研究低pH對克氏原螯蝦鰓的呼吸代謝酶的影響時指出,LDH活力上升,表明此時機體有氧代謝受阻,會迅速提高無氧代謝強度來滿足自身缺氧條件下的能量需求。類似的呼吸代謝變化在日本囊對蝦[21]干露脅迫第1和第3小時也有發生。本研究在12~48 h干露時間內LDH活力升高,表明機體無氧呼吸代謝強度增加,LDH催化還原丙酮酸致使LA逐漸積累[22],同時SDH作為有氧呼吸中琥珀酸轉變為延胡索酸的關鍵脫氫酶,其在干露期間活力降低也同樣反映了機體有氧代謝水平的下降[23-24],克氏原螯蝦的呼吸方式逐漸從有氧代謝向無氧代謝轉變。在干露48~132 h階段,LDH活力和LA濃度逐漸降低,脊尾白蝦[7]在常溫干露脅迫末期也出現類似的酶活力下降變化,推測過長的脅迫時間導致機體的代謝系統受到影響,LA濃度的下降是機體應對嚴重的低氧脅迫將LA轉化為葡萄糖來維持內環境穩態所做的調整[25],這種調整是機體短時間內為消耗過量LA而采取的糖異生作用[25],但長時間的干露脅迫,使機體耐受能力到達極限,致使酶的調節功能喪失,內穩態失衡,造成不可逆損傷[5]。
3.3 干露脅迫對肝胰腺、鰓和肌肉組織結構的影響
組織結構變化反映出機體生理狀態的改變[26]。肌肉組織擁有致密的結締組織,蛋白質是肌肉組織的主要組成成分。有研究指出,肌肉蛋白溶解性與蛋白持水性息息相關[27],尼羅羅非魚 (Oreochromis niloticus) 在冷藏期間肌肉肌原纖維蛋白含量下降,肌肉蛋白溶解性降低[28]。本研究中雄、雌蝦在干露第24小時后肌肉纖維出現有規律的疏松、褶皺、彎曲現象,推測可能是在干露條件下肌肉肌原纖維蛋白含量和持水量下降,導致肌肉收縮,從而產生組織損傷。
鰓作為參與氣體交換和離子調節的重要器官,通常最先受到環境脅迫影響并造成損傷[29]。本研究中克氏原螯蝦的鰓組織在干露第12小時出現輕微損傷,第24和第36小時大部分呼吸上皮細胞脫落。有研究表明呼吸上皮細胞的脫落使機體氣體交換的能力減弱,造成組織缺氧[20],而本研究中干露第24和第36小時無氧代謝酶LDH處于活力最大值,LA濃度增大,無氧代謝水平增強,結合組織結構變化可知,干露脅迫導致機體鰓組織損傷,損傷的鰓組織使機體獲氧能力下降,缺氧條件下機體通過提高無氧代謝水平滿足自身能量需求[20,30]。克氏原螯蝦鰓組織隨著干露時間的延長損傷加劇,本研究干露第96和第132小時的損傷與過量的硫酸鋅 (ZnSO4) 脅迫[30]條件下克氏原螯蝦鰓組織出現情況類似,包括鰓膜和呼吸上皮細胞分離,許多呼吸上皮細胞壞死、脫落,失去正常鰓細胞結構。干露條件下鰓缺少用于有氧呼吸的溶解氧,同時鰓組織的嚴重損傷進一步降低了自身氧氣的利用,嚴重影響了機體的離子代謝和滲透調節功能[31],制約了呼吸代謝的進行,造成克氏原螯蝦死亡。
肝胰腺是克氏原螯蝦重要的代謝器官,具有消化、吸收、儲存和排泄的功能[32]。對于不同脅迫,甲殼動物的肝胰腺細胞會出現一些相似的變化,鹽度9.0暴露下的三疣梭子蟹[33]、氨氮 (NH4-N) 脅迫 15 d 后的中華絨螯蟹[34](Eriocheir sinensis) 以及在低pH脅迫下的克氏原螯蝦[20]皆出現了B細胞內部空泡增多、體積增大的現象。B細胞具有消化、吸收營養物質的功能,其數量多少反映機體消化能力的強弱[35]。本研究中B細胞及內部轉運泡增多、體積增大,可能有助于加快營養物質的運輸[33],進而有助于克氏原螯蝦消耗營養物質來提供能量[26]。R細胞除具有吞噬作用外,還可以儲存脂肪和糖原。本研究中R細胞空泡化可能是細胞中營養物質的消耗造成的[33,36],尤其在干露48 h后空泡化的細胞數量增多,約占據整個細胞的80%,甚至出現肝細胞溶解的現象。此外,隨著干露脅迫時間的延長,肝胰腺細胞管腔逐漸變大,在干露第132小時約占細胞體積的50%。干露脅迫下伴隨失水脅迫,肝小管收縮變形,進一步表明機體細胞嚴重失水,過長時間的脅迫已嚴重破壞肝胰腺的正常組織結構,可能影響機體的正常生理功能。
綜上,鰓組織在干露第12小時即出現損傷,隨著干露時間的延長肝胰腺組織損傷愈嚴重,兩者在干露第36小時均受到嚴重損傷,影響了基本結構功能。雌蝦體質量在干露第48小時后下降顯著,而雄蝦在第60小時后下降明顯,同時以無氧呼吸代謝為主的呼吸強度減弱,機體面臨耐受極限。因此,克氏原螯蝦雌、雄成蝦抗干露脅迫能力存在差異,在進行成體運輸時干露時間不宜超過36 h。
