黃芪甲苷對博來霉素肺纖維化模型小鼠miR-29b及其相關因子表達的影響
2021-06-24 00:26:06童佳歡劉肇恒龍杞郝慶勛王域辰焦揚
環球中醫藥 2021年6期
童佳歡 劉肇恒 龍杞 郝慶勛 王域辰 焦揚
特發性肺纖維化(idiopathic pulmonary fibrosis, IPF)是一種原因尚不明確的,以肺部彌漫纖維化伴蜂窩囊樣改變為特征的疾病。患者診斷后中位生存期僅2~3年。轉化生長因子β1(transforming growth factor-β1,TGF-β1)是促進肺纖維化發生發展的最重要因素之一。研究發現,TGF-β1是MicroRNA-29b(miR-29b)的下游靶基因之一,miR-29b的過表達可以直接抑制TGF-β1/Smad3信號通路,從而達到抗纖維化的作用[1-2]。此外,miR-29b可顯著提高超氧化物歧化酶(superoxide dismutase, SOD)活性,降低活性氧(reactive oxygen species,ROS)、丙二醛(malondialdehyde,MDA)含量,從而抑制氧化應激,減輕肺纖維化[3-4]。miR-29b這種“多途徑”“多靶點”作用特點,與中醫學的調整陰陽平衡,扶正祛邪的治療理念相吻合。
課題組總結多年臨床經驗,認為IPF以肺氣虧虛為本,臨床運用肺痹湯治療取到了較好的療效[5]。課題組通過動物及細胞實驗研究證實,肺痹湯可調節肺組織TGF-β1水平,對成纖維細胞的TGF-β1/smad信號通路具有調控作用[6-7]。肺痹湯以黃芪為君,黃芪既能補益肺氣,亦能通調血脈。實驗研究發現,黃芪總皂苷可抑制博來霉素誘導的肺組織炎癥反應,下調TGF-β1表達[8],抑制肺纖維化發展,其中黃芪甲苷能顯著抑制博來霉素誘導的小鼠氧化應激狀態,延緩 IPF進程[9-10],但其抗肺纖維化機制仍需深入研究。本實驗基于miR-29b,研究黃芪甲苷對博來霉素肺纖維化模型小鼠的干預作用,探討其防治肺纖維化的可行性及作用機制,為臨床治療肺纖維化提供思路和方法。
1 材料與方法
1.1 實驗動物
SPF級C57BL雄性小鼠40只,體質量(22±2)g,購于北京維通利華有限公司,動物許可證號:SCXK(京)2006-009,飼養于北京中醫藥大學第二附屬醫院實驗動物中心。飼養環境:室溫22~26℃,相對濕度60%~80%,12小時/12小時明暗周期照射。
1.2 實驗藥物及試劑
黃芪甲苷(質量分數≥98%),購自上海源葉生物科技有限公司;miR-29b模擬劑(批號:miR4CM001)、miR-29b模擬劑陰性對照藥(批號:RIBOCSUTOM001)購于廣州銳博生物科技有限公司;HE染色試劑盒、Masson三色染色液購于北京索萊寶科技有限公司;羥脯氨酸(hydroxyproline,HYP)試劑盒購于南京建成有限公司;MDA試劑盒(貨號:DG30429M)、ROS試劑盒(貨號:DG30626M)、SOD試劑盒(貨號:DG30430M)購于北京冬歌博業生物科技有限公司。
1.3 模型制備
實驗小鼠適應性喂養1周后隨機分為空白組8只,造模組32只。造模組小鼠稱重后,以異氟烷麻醉。將小鼠上牙迅速掛于懸線,使小鼠呈豎直位,用彎鑷將小鼠舌頭拽出,用100 μL移液槍向氣管內快速滴入博來霉素溶液(2 u/kg)后,保持豎直狀態片刻,使藥液嗆入氣管中,旋轉1分鐘,使液體在小鼠肺內均勻分布,隨后放回鼠籠,待自然蘇醒。空白組以相同方法滴入等量PBS。
1.4 分組及給藥
造模組小鼠成功造模后,隨機分為模型組、miR-29b組、miR-29b陽性對照藥組(簡稱NC組)、黃芪甲苷組,每組8只。造模1天后開始給藥,每天上午9點給藥。黃芪甲苷組以黃芪甲苷40.8 mg/(kg·d)灌胃,空白組、模型組、miR29組、NC組分別以生理鹽水10 mL/(kg·d)灌胃。參照文獻[11],miR-29b組以miR-29b模擬劑10 mg/(kg·3d)氣道內滴入,NC組以miR-29b模擬劑陰性對照藥10 mg/(kg·3d)氣道滴入,空白組、模型組、中藥各組以等量PBS氣道滴入。
1.5 觀察指標及方法
1.5.1 一般狀態觀察 觀察各組小鼠精神狀態、毛發變化、進食、飲水及體質量、呼吸情況變化,記錄小鼠死亡情況,進行死亡原因分析。
1.5.2 肺組織病理學染色 將小鼠左肺置于中性福爾馬林液中固定,常規石蠟包埋切片,用HE染色觀察肺組織肺泡炎程度;Masson染色觀察肺組織肺纖維化程度。觀察各組小鼠肺組織病理學改變,主要包括肺泡炎性細胞浸潤、肺組織水腫、肺間質纖維化等。
1.5.3 肺組織羥脯氨酸檢測 取小鼠右側肺組織放入液氮速凍,保存在-80℃,采用羥脯胺酸試劑盒(堿水解法)測定肺組織中羥脯胺酸含量。
1.5.4 Elisa檢測血清中SOD、MDA、ROS水平 小鼠稱重后用異氟烷麻醉,摘眼球取血,分離上血清,采用Elisa試劑盒檢測血清中SOD、MDA、ROS水平。
1.5.5 qPCR法檢測肺組織中miR-29b、TGF-β1、smad3水平 根據目的基因設計引物(見表1),提取總RNA檢測純度,運用反轉錄合成cDNA,建立擴增體系,讀取擴增曲線、溶解曲線及目的基因的相對表達量。

表1 引物序列
1.5.6 Western-blot檢測肺組織中TGF-β1、smad3、P-smad3水平 將少量肺組織塊置于1~2 mL勻漿器中球狀部位,充分研磨組織塊。加入400 μL的裂解液進行勻漿,置于冰上。待樣本充分裂解完成后,在4°C下12000 rpm,離心5分鐘,取上清于-20℃保存。制作標準曲線,檢測樣品蛋白含量,進行電泳、轉膜、免疫反應、化學發光、顯影、定影,最后進行凝膠圖像分析。
1.6 統計學方法

2 結果
2.1 各組小鼠一般情況比較
空白組小鼠精神狀態好,活潑好動,毛發光澤,呼吸平穩。模型組及NC組小鼠精神較差,少動,體質量減輕,毛發干枯,可見喘息氣促。miR-29b組及黃芪甲苷組小鼠癥狀較輕。
從第1天到第9天,除空白組外,各組小鼠體質量均下降,第9天至第28天,各組小鼠體質量逐漸增加。與空白組相比,模型組及黃芪甲苷組第5、9、13、17、21、25、28天體質量減輕(P<0.05),NC組第5、9、17、21、25、28天體質量減輕(P<0.05),miR-29b組第9、28天體質量減輕(P<0.05)。與模型組和NC組相比,空白組、miR-29b組和黃芪甲苷組第9天時體質量較高(P>0.05)。見表2。
表2 各組博來霉素肺纖維化模型小鼠體質量變化比較
2.2 各組小鼠肺組織病理比較
空白組無炎性細胞浸潤,管腔無滲出,無膠原纖維沉積。模型組和NC組可見管腔大量炎性滲出,肺泡間隔增寬、充血,肺泡破壞嚴重,Masson染色可見大量膠原纖維沉積,纖維結節生成。miR-29b組和黃芪甲苷組可見部分炎性細胞浸潤,Masson染色可見少量膠原纖維沉積。見圖1~2。
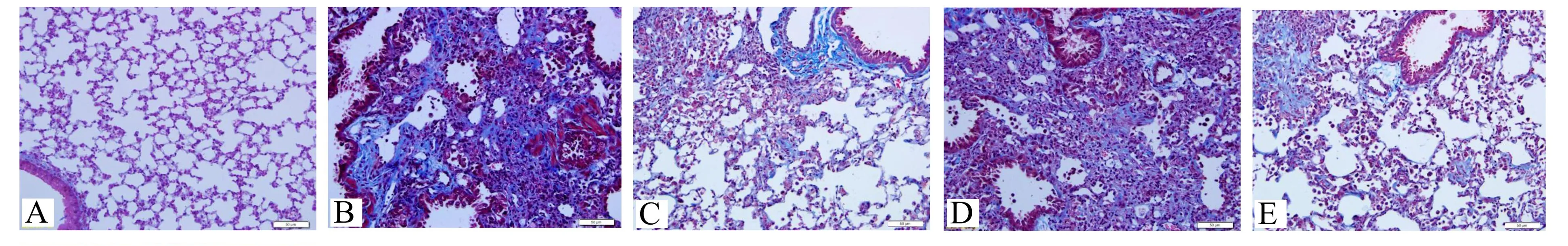
注:A空白組,B模型組,C miR-29b組,D NC組,E黃芪甲苷組。
2.3 各組小鼠肺組織HYP含量比較
與空白組相比,模型組、miR-29b組、NC組、黃芪甲苷組羥脯氨酸含量均升高(P<0.05)。與模型組相比,miR-29b組和黃芪甲苷組羥脯氨酸含量較低(P<0.05)。見表3。
表3 各組博來霉素肺纖維化模型小鼠肺組織HYP含量比較
2.4 各組小鼠血清SOD、MDA、ROS含量比較
與空白組相比,模型組、miR-29b組和黃芪甲苷組的SOD含量減少,MDA和ROS含量增加(P<0.05),NC組的SOD顯著減少、MDA增加(P<0.05)。與模型組相比,miR-29b組的MDA、ROS含量減少(P<0.05),NC組的SOD含量減少(P<0.05),黃芪甲苷組的SOD含量明顯增多,MDA和ROS含量減少(P<0.05)。與NC組相比,miR-29b組和黃芪甲苷組SOD含量增加,ROS含量減少(P<0.05)。見表4。

表4 各組博來霉素肺纖維化模型小鼠血清SOD、MDA、ROS含量比較
2.5 各組小鼠肺組織TGF-β1、Smad3、miR-29b mRNA含量比較
與空白組相比,模型組和NC組的TGF-β1、Smad3基因表達量明顯升高,miR-29b基因表達量減少(P<0.05);miR-29b組的Smad3基因表達量減少(P<0.05)。與模型組相比,miR-29b組和黃芪甲苷組的Smad3表達量減少、miR-29b表達量升高(P<0.05)。與NC組相比,miR-29b組的Smad3表達量減少、miR-29b表達量升高(P<0.05),黃芪甲苷組Smad3表達量減少(P<0.05);與miR-29b組相比,黃芪甲苷組的Smad3含量升高(P<0.05)。見表5。
表5 各組博來霉素肺纖維化模型小鼠肺組織TGF-β1、Smad3、miR-29b mRNA含量比較
2.6 各組小鼠肺組織TGF-β1、Smad3、P-smad3蛋白含量比較
與空白組相比,模型組和NC組的TGF-β1、P-smad3蛋白含量升高(P<0.05)。與模型組相比,miR-29b組和黃芪甲苷組的TGF-β1、P-smad3蛋白含量減少(P<0.05)。與NC組相比,miR-29b組的TGF-β1、P-smad3蛋白含量減少(P<0.05),黃芪甲苷組P-smad3蛋白含量減少(P<0.05)。見表6、圖3。
表6 各組博來霉素肺纖維化模型小鼠肺組織TGF-β1、Smad3、P-smad3蛋白含量比較
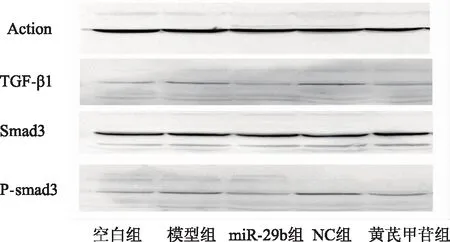
圖3 各組博來霉素肺纖維化模型小鼠肺組織TGF-β1、Smad3、P-smad3蛋白含量表達
3 討論
肺纖維化發病機制尚未完全明了,已有的研究認為多種因素共同參與了肺纖維化的發生、發展過程。MicroRNA是一種非編碼的單鏈小分子RNA,通過與mRNA互補配對的方式,在轉錄后調控蛋白的表達。研究發現,同一種microRNA可以調控多種mRNA的表達,而同種mRNA也可能受到不同的microRNA調控[12-13]。miR-29家族由miR-29a、miR-29b、miR-29c三種成員構成,這三種成員之間僅存在2~3個核苷酸位點的差異,且具有相同的種子序列,因此它們作用的靶基因也往往相同,具有高度保守性[14]。miR-29可直接抑制膠原纖維生成,或通過多種細胞因子抑制肺纖維化[15]。氧化應激是觸發肺纖維化形成的重要機制之一,肺損傷后釋放大量的ROS,使肺組織發生過氧化,刺激膠原蛋白的合成與分泌[16]。另一方面,ROS也可通過激活TGF-β1/Smad3信號通路,增加結締組織生長因子和基質金屬蛋白酶的表達,促進細胞外基質的重構和肺纖維化[17-18]。研究表明,miR-29家族的過表達可以降低ROS和MDA含量,清除氧化應激產物,同時顯著提高SOD含量,提高機體抗氧化水平,從而減輕肺纖維化程度[3,19]。miR-29也可直接作用于靶基因TGF-β1的3’-UTR[20],抑制TGF-β1表達,并通過TGF-β1/Smad3信號通路的抑制作用[21-22],減輕肺纖維化。這種多靶點、多途徑的調控方式與中醫整體觀具有相似性。
肺纖維化在古代文獻中無對應病名,根據其臨床表現,大多數學者認為可以歸為“肺痿”“肺痹”范疇。《醫門法律》有言“肺痿者,肺氣痿而不振也”,《辨證錄》中說:“肺痹之成于氣虛,盡人而不知也,……然肺痹即氣痹也。”肺纖維化多因氣虛而起,屬本虛標實之證,臨床也常以補氣之品治療肺纖維化。研究發現,補氣藥在治療肺纖維化的過程中使用頻率較高,其中黃芪為最常用的補氣藥之一[23]。
近年來對黃芪研究發現,黃芪甲苷是黃芪的主要活性成分,可有效改善抗氧化酶活性,抑制氧化應激反應[24],也可降低肺組織羥脯氨酸含量,下調致纖維化因子TGF-β1表達水平,明顯改善博來霉素誘導的肺纖維化程度[25]。但黃芪甲苷調節TGF-β1的機制還需進一步研究,對microRNA的干預也有待更多探索。
本實驗研究以miR-29b為切入點,研究黃芪甲苷對肺纖維化小鼠的作用機制。結果表明,博來霉素肺纖維化模型組小鼠早期出現體質量下降,miR-29b可以減輕小鼠體質量下降的程度,黃芪甲苷也有緩解體質量減輕的趨勢。病理結果顯示,模型小鼠肺組織結構破壞嚴重,大量膠原纖維沉積,黃芪甲苷可以減少膠原纖維形成,減輕肺纖維化程度。羥脯氨酸是膠原纖維的主要成分,本實驗結果提示黃芪甲苷可以通過減少羥脯氨酸含量抑制肺纖維化。同時,黃芪甲苷可以有效增加SOD含量,抑制MDA的生成,提示黃芪甲苷具有抑制氧化反應的作用。進一步實驗發現,模型組小鼠肺組織miR-29b的基因表達減少,而黃芪甲苷可以一定程度上提高miR-29b的表達,說明黃芪甲苷對miR-29具有一定的調控作用。黃芪甲苷也可減少Smad3基因的表達,抑制其磷酸過程,減少了P-Smad3蛋白水平,同時也可以減輕TGF-β1蛋白含量,從而起到調控TGF-β1/Smad3信號通路的作用。
綜上所述,本研究發現黃芪甲苷可以通過調控miR-29b減輕氧化應激水平,抑制TGF-β1/Smad3信號通路,減少膠原沉積,從而改善肺纖維化病理損傷。但黃芪甲苷的臨床干預效果仍需進一步研究。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中老年保健(2022年2期)2022-11-25 23:46:31
昆明醫科大學學報(2022年4期)2022-05-23 13:04:50
安徽醫科大學學報(2022年1期)2022-02-22 08:47:42
學苑創造·A版(2020年9期)2020-10-13 09:41:02
臨床醫藥文獻雜志(電子版)(2020年98期)2020-04-29 09:57:02
國際呼吸雜志(2019年21期)2019-11-25 09:52:20
吉林大學學報(醫學版)(2015年3期)2015-12-17 07:47:38
天津醫科大學學報(2015年3期)2015-06-05 12:21:49
中國當代醫藥(2015年20期)2015-03-01 02:04:38
