雙委夜蛾氣味結合蛋白AdisOBP6的原核表達、抗體制備及表達譜分析
2021-04-13 10:37:12宋月芹宋智煜董鈞鋒陳慶霄孫會忠
昆蟲學報 2021年2期
宋月芹, 宋智煜, 董鈞鋒, 陳慶霄, 孫會忠
(河南科技大學林學院, 河南洛陽 471023)
雙委夜蛾Athetisdissimilis隸屬鱗翅目(Lepidoptera)夜蛾科(Noctuidae)委夜蛾屬Athetis,是我國重要的農業害蟲,對小麥、玉米、花生、大豆和甘薯等作物造成嚴重損失。該害蟲主要分布于中國、日本、朝鮮、印度、印度尼西亞和菲律賓等國家(Takahashi, 1975; Banetal., 1998; Choetal., 2010; 李靜雯等, 2012)。雙委夜蛾首次在山東威海玉米地發現為害(李靜雯等, 2012),隨后在河南、陜西和安徽等地也陸續出現該蟲的報道(宋月芹等, 2015; 段愛菊等, 2016; 郭婷婷等, 2016a, 2016b)。雙委夜蛾幼蟲和成蟲在外形上與二點委夜蛾Athetislepigone極其相似,為害特征也相同,常隱蔽在秸稈、落葉和雜草中,在田間與二點委夜蛾混合發生。據統計,近6年二點委夜蛾為害面積近220萬hm2,缺苗率高達70%以上,給玉米產量造成巨大損失(王振營等, 2012),而其中可能有一部分是雙委夜蛾為害的。在室內用玉米苗飼養試驗發現,雙委夜蛾的幼蟲食量、幼蟲取食期和產卵量遠大于二點委夜蛾(郭婷婷等, 2016a)。在田間,農民常將雙委夜蛾與二點委夜蛾混在一起防治,長期施用廣譜性殺蟲劑使害蟲極易產生抗藥性,防治效果下降,對人畜不安全還污染環境。目前國內外專家通過研究昆蟲基因功能等方面,以探索新型的對害蟲專一性強、對人畜安全的綠色防控技術,已經取得了一定進展,而目前雙委夜蛾這方面的研究較少。
昆蟲具有靈敏的嗅覺系統,可以通過嗅覺精準地定位到配偶、寄主植物以及天敵(Leal, 2013),因此人們為了保護環境,探索新型害蟲防治技術,對昆蟲有關嗅覺基因開展了大量研究。如氣味結合蛋白(odorant binding proteins, OBPs)(Gaoetal., 2018; Zhangetal., 2019; 鞏雪芳等, 2020; Wangetal., 2020)、化學感受蛋白(chemosensory proteins, CSPs)(Qiuetal., 2018; Alietal., 2019)、嗅覺受體(olfactory receptors, ORs)(陳麗慧等, 2019; Tiwarietal., 2019)、味覺受體(gustatory receptors, GRs)(Poudeletal., 2015; Liuetal., 2020)、離子型受體(ionotropic receptors, IRs)(Abuinetal., 2019)、感覺神經元膜蛋白(sensory neuron membrane proteins, SNMPs)(胡穎穎等, 2013; Blankenburgetal., 2019)和氣味降解酶(odorant degrading enzymes, ODEs)(Prestwich, 1987; Chertempsetal., 2015)等。其中OBPs是一類可溶性的小分子蛋白,在昆蟲觸角感器中高度表達,為氣味識別的第一步(Pelosi and Maida, 1995)。根據半胱氨酸的數量,將OBPs分為5類:含有6個保守半胱氨酸位點的Classic OBPs;在第6個半胱氨酸后面多2~3個半胱氨酸殘基,并且還有一個保守的脯氨酸位點的Plus-C OBPs;只有4個或5個半胱氨酸的Minus-C OBPs;具有2個典型半胱氨酸排列模式的Dimer OBPs;基因序列較長,特征不明顯的Atypical OBPs(Zhangetal., 2013; McKenzieetal., 2014; Guetal., 2015)。
關于昆蟲觸角中OBPs的研究很多(Fanetal., 2011; Costa-da-Silvaetal., 2013; Ishidaetal., 2013; Duetal., 2019),但對昆蟲精巢中OBPs的報道很少。Li等(2008)發現埃及伊蚊AedesaegyptiOBP22蛋白在雄蛾生殖器內大量表達,并在交配過程中將該蛋白轉運到雌蛾的腹部末端和受精囊中,推測該蛋白可能作為某種信息素的載體,類似于脊椎動物的尿液和唾液。Sun等(2012)研究發現OBP10蛋白在棉鈴蟲Helicoverpaarmigera和煙青蟲H.assulta雄蛾昆蟲的精液中也大量表達,交配后將該蛋白轉移到雌蛾生殖器內,隨著精子與卵細胞的結合傳遞給受精卵,推測該蛋白可能攜帶保護卵的防御物質十二碳烯(dodecene),在雌雄成蟲交配時傳遞給受精卵,避免受孕雌蛾在其附近產卵,從而減少了種群內和種群之間的自相殘殺。同樣,我們前期在對雙委夜蛾OBPs研究時,也發現AdisOBP6在精巢中大量表達,其表達量遠大于在觸角中的(Sunetal., 2016)。為了進一步弄清AdisOBP6的表達規律,我們通過原核表達和抗體制備,用熒光定量PCR和Westen blot印跡方法分別對AdisOBP6在mRNA和蛋白水平上的表達情況進行分析,為今后進一步研究AdisOBP6基因的功能奠定基礎。
1 材料與方法
1.1 供試蟲源
本研究中所用的雙委夜蛾來自河南科技大學林學院昆蟲實驗室,該群體自2017年7月在洛陽周邊玉米田誘集成蟲,然后在室內繼代飼養至今。室內飼養條件為:溫度27±1℃,相對濕度80%±5%,光周期16L∶8D。
1.2 AdisOBP6蛋白的結構分析
首先確定模板,將AdisOBP6(GenBank登錄號: ALZ45421.1)氨基酸序列提交到Meta(http:∥bioinfo.pl/meta)在線網站,尋找同源序列,同源性在30%以上的氨基酸序列就可以作為模板用于三維結構的構建;然后采用Discover Studio 2.0軟件工具中的Homology模塊中的Modeler程序,以已知三維結構的同源序列模板預測AdisOBP6蛋白的三維結構。為了模擬蛋白的真實性,需要將目的蛋白的信號肽去除,根據模擬3D結構的得分情況,選取得分最高的結構作為最終結構。同時將比對文件用ESPript 3.0軟件進行二級結構優化(Robert and Gouet, 2014)。
1.3 總RNA的提取及cDNA第1鏈的合成
收集雙委夜蛾羽化后第3天的雌雄成蟲觸角、末齡幼蟲精巢(因為幼蟲雌雄難辨,我們采用隨機方法解剖,以觀察到精巢為準)、化蛹第3天的精巢以及成蟲羽化后第1, 2, 3和5天的精巢,還有未受精卵和受精卵。將雙委夜蛾各部分組織解剖后立即浸在液氮中的1.5 mL離心管內,然后保存在-80℃超低溫冰箱中備用。
總RNA的提取采用RNAiso Plus Kit(TaKaRa,北京)試劑盒進行,并使用RNase-free DNase I(TaKaRa,北京)對提取的RNA進行除DNA處理。采用1%的瓊脂糖凝膠電泳和NanoDrop 2000c分光光度計(Thermo Scientific)進行質量檢測。采用PrimeScriptTM1st Strand cDNA Synthesis Kit(TaKaRa,北京)試劑盒對RNA進行反轉錄合成cDNA第1鏈。
1.4 重組表達載體的構建
去掉AdisOBP6蛋白序列N端信號肽,在蛋白C端加入His標簽和終止密碼子TAA,序列經過密碼子優化后,并添加NdeI/XhoI為酶切位點序列(引物中下劃線序列),由上海生工合成引物序列,AdisOBP6-NdeI-F: 5′-CCCATATGGGAGCCTGGATG AACTG-3′; AdisOBP6-XhoI-R: 5′-CCCTCGAGGGGA TGATGGTGATGATGT-3′。以1.3節合成的雙委夜蛾末齡幼蟲精巢cDNA為模板,用帶有NdeI和XhoI酶切位點的AdisOBP6特異引物進行PCR擴增,PCR反應體系(20 μL): cDNA模板1 μL, 上下游引物(10 μmol/L)各1.5 μL, dNTPs (2.5 μmol/L)1.6 μL,ExTaqDNA聚合酶(TaKaRa,大連)0.2 μL, 10×ExTaqbuffer 2 μL, ddH2O為12.2 μL。PCR反應條件: 94℃變性5 min; 94℃變性30 s, 55℃退火30 s, 72℃延伸30 s, 35個循環; 最后72℃延伸10 min。膠回收目的片段,將目的片段克隆到pMDTM19-T載體上,轉化大腸桿菌EscherichiacoliDH5α感受態細胞,挑取陽性克隆培養過夜,提取質粒。將該質粒酶切后,用T4 DNA連接酶連接到酶切過的pET-28a(+)載體上,克隆到大腸桿菌DH5α感受態細胞中,涂板過夜后挑去陽性克隆,搖菌提取組合后的pET-28a/AdisOBP6質粒,雙酶切測序檢測,挑取正確的質粒備用。
1.5 AdisOBP6重組蛋白的表達、純化及抗原制備
取1 μL重組的pET-28a/AdisOBP6質粒轉化到大腸桿菌BL21(DE3)感受態細胞中,涂布在LB固體培養基(含卡那霉素)上,37℃培養過夜。挑取陽性單菌落于5 mL的LB培養基(含卡那霉素)中37℃ 220 r/min搖菌培養過夜培養。將培養的菌液按1∶100(v/v)比例接種于3 L的LB液體培養基(含卡那霉素)中,37℃ 220 r/min培養,當OD值達到0.6時,添加終濃度為0.5 mmol/L IPTG,220 r/min,20℃和37℃分別誘導過夜,離心收集細胞菌體。
將收集的菌液超生破碎,4℃ 12 000 r/min離心20 min,收集上清和沉淀,SDS-PAGE蛋白電泳檢測蛋白的可溶性。采用鎳瓊脂糖親和層析法純化蛋白,用20, 50和500 mmol/L咪唑洗脫,純化的蛋白經SDS-PAGE檢測后,在1×PBS(pH 7.4)中4℃透析過夜,再用PEG20000濃縮,0.45 μm濾膜過濾后,進行SDS-PAGE電泳檢測。檢測合格后,分裝1 mL/管,共5管,-80℃凍存,用于抗體制備。
1.6 AdisOBP6抗體的制備
初免抗原為蛋白抗原與等體積弗氏完全佐劑混勻乳化,二免、三免、終免抗原為蛋白抗原與等體積弗氏不完全佐劑混勻乳化,免疫新西蘭大白兔4次。采用ELISA法檢測抗體效價,將AdisOBP6抗體分別稀釋后,在450 nm的波長下測OD值,血清效價值≥2.5×陰性值為合格。檢測抗體為陽性后保存于-80℃備用。抗體由生工生物工程(上海)股份有限公司制備。
1.7 AdisOBP6表達譜測定
熒光定量PCR在StepOne Plus型熒光定量PCR(ABI, Foster, CA, 美國)儀器上進行操作,使用2×SG Fast qPCR Master Mix(High Rox,B639273,BBI)試劑盒測定。雙委夜蛾AdisGAPDH(GenBank登錄號: KT361883)基因作為內參基因。采用Primer3在線工具設計引物,AdisOBP6正向引物: 5′-AGACAGAAGACAATGCCTGACAGTG-3′;反向引物: 5′-CTGACCTCAACATCATTCACGCTT-3′。AdisGADPH正向引物: 5′-CTGCTCATTTAGAGGGTGGTGC-3′;反向引物: 5′-TCTTCTGGGTAGCGGTGGTAG-3′。 將1.3節合成的cDNA樣品稀釋10倍作為模板上機檢測。PCR反應體系: SybrGreen qPCR Master Mix 10 μL, 正反引物(10 μmol/L)各0.4 μL, cDNA模板(200 ng/μL)2 μL, 7.2 μL ddH2O補齊至20 μL。反應程序: 95℃預變性3 min; 95℃變性5 s, 60℃退火并延伸30 s, 共45個循環。熔點曲線設置為95℃持續15 s,降到60℃持續60 s,再升到95℃保持15 s。每個組織3次生物學重復,每生物學重復雌雄觸角各來自50頭個體、不同時期的精巢來自20頭個體、受精卵和非受精卵各500粒,技術重復3次。以雙委夜蛾雌成蟲觸角中基因表達量為對照,利用相對表達量公式2-ΔΔCt進行計算(Livak and Schmittgen, 2001)。
提取雙委夜蛾羽化后第3天的雌雄成蟲觸角、末齡幼蟲精巢、化蛹第3天的精巢以及成蟲羽化后第1, 2, 3和5天的精巢,還有未受精卵和受精卵10種組織的總蛋白,每種組織重復3次。蛋白變性后,使用5%濃縮膠和15%的分離膠進行SDS-PAGE蛋白電泳。然后在200 mA電流下持續50 min,進行轉膜。用含5%脫脂奶粉的TBST封閉液,室溫搖床封閉2 h,一抗(AdisOBP6抗體)孵育4℃過夜,二抗37℃搖床孵育2 h。ECL顯色曝光,拍照。為了檢測樣品蛋白情況,用小鼠單抗β-actin(預實驗顯示蛋白表達穩定)做一抗為對照組,進行Westen blot檢測。
1.8 數據分析
數據采用SPSS 17.0(IBM, Chicago, IL, 美國)軟件中的單因素方差分析(one-way ANOVA)和Tukey氏HSD法進行差異顯著性分析,P<0.05表示樣品間存在顯著性差異。
2 結果
2.1 AdisOBP6的結構特征
通過結構比對得出AdisOBP6與岡比亞按蚊AnophelesgambiaeAgamOBP4結構最為相似,該結構為X衍射得到的蛋白結構,可靠性強,因此選擇AgamOBP4的結構作為AdisOBP6的三維結構預測模板。比對可見,AdisOBP6具有氣味結合蛋白保守的6個半胱氨酸,并具有7個α-螺旋結構(圖1: A, B),蛋白N端和C端明顯可見(圖1: B),通過表面結構模擬,可見α-螺旋形成一個結合口袋,用于運輸氣味分子(圖1: C)。
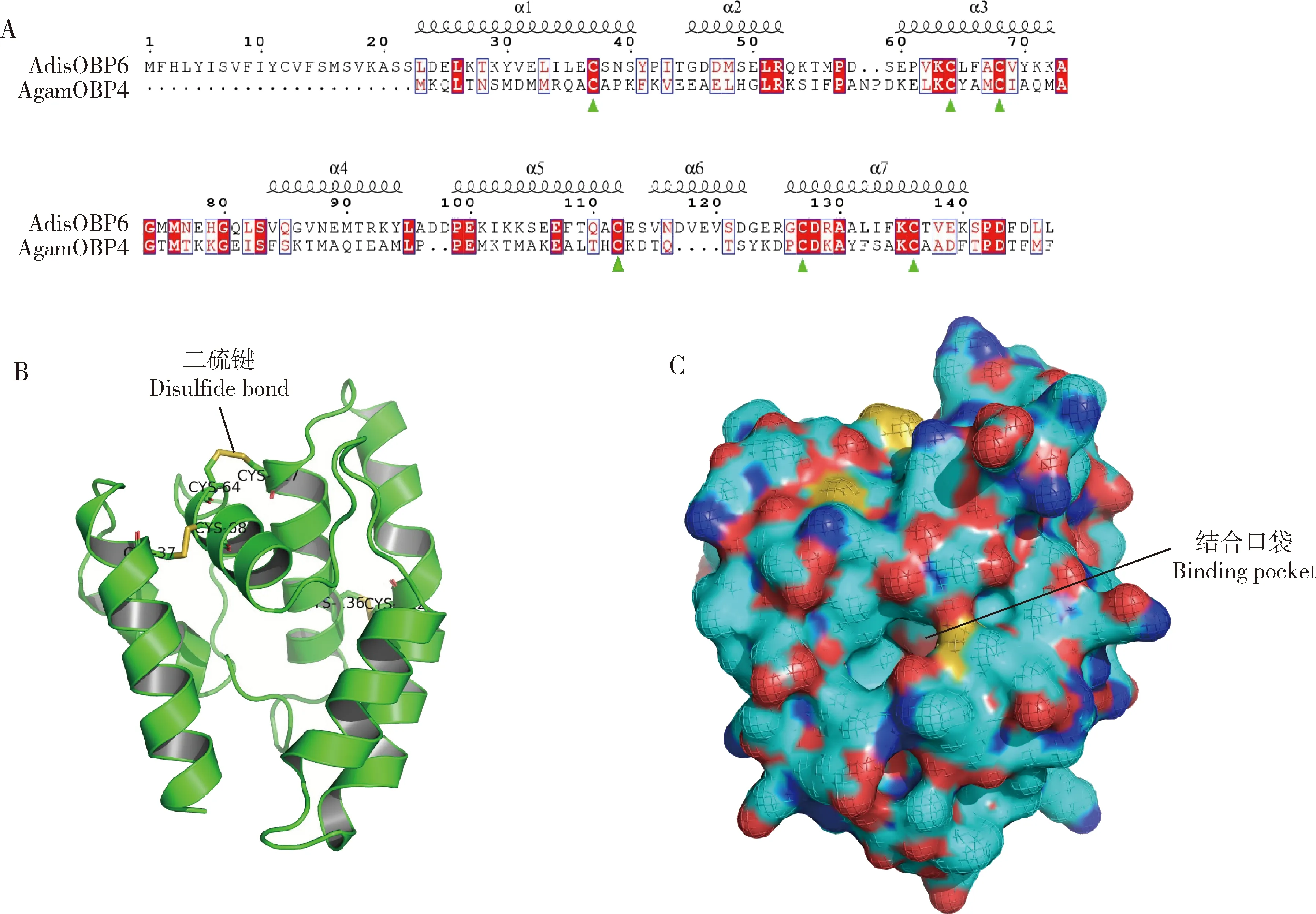
圖1 雙委夜蛾AdisOBP6二級結構及三級結構模擬圖
2.2 原核表達載體的構建和鑒定
用帶有酶切位點的引物PCR擴增重組質粒pET-28a/AdisOBP6,如圖2所示,在400 bp左右的地方出現目的條帶,與理論大小基本相符合。挑選pET-28a/AdisOBP6陽性轉化子經質粒小提后分別用NdeI和XhoI進行雙酶切鑒定,出現5 kb和400 bp左右的兩個序列片段,與預測目的條帶的結果相一致。
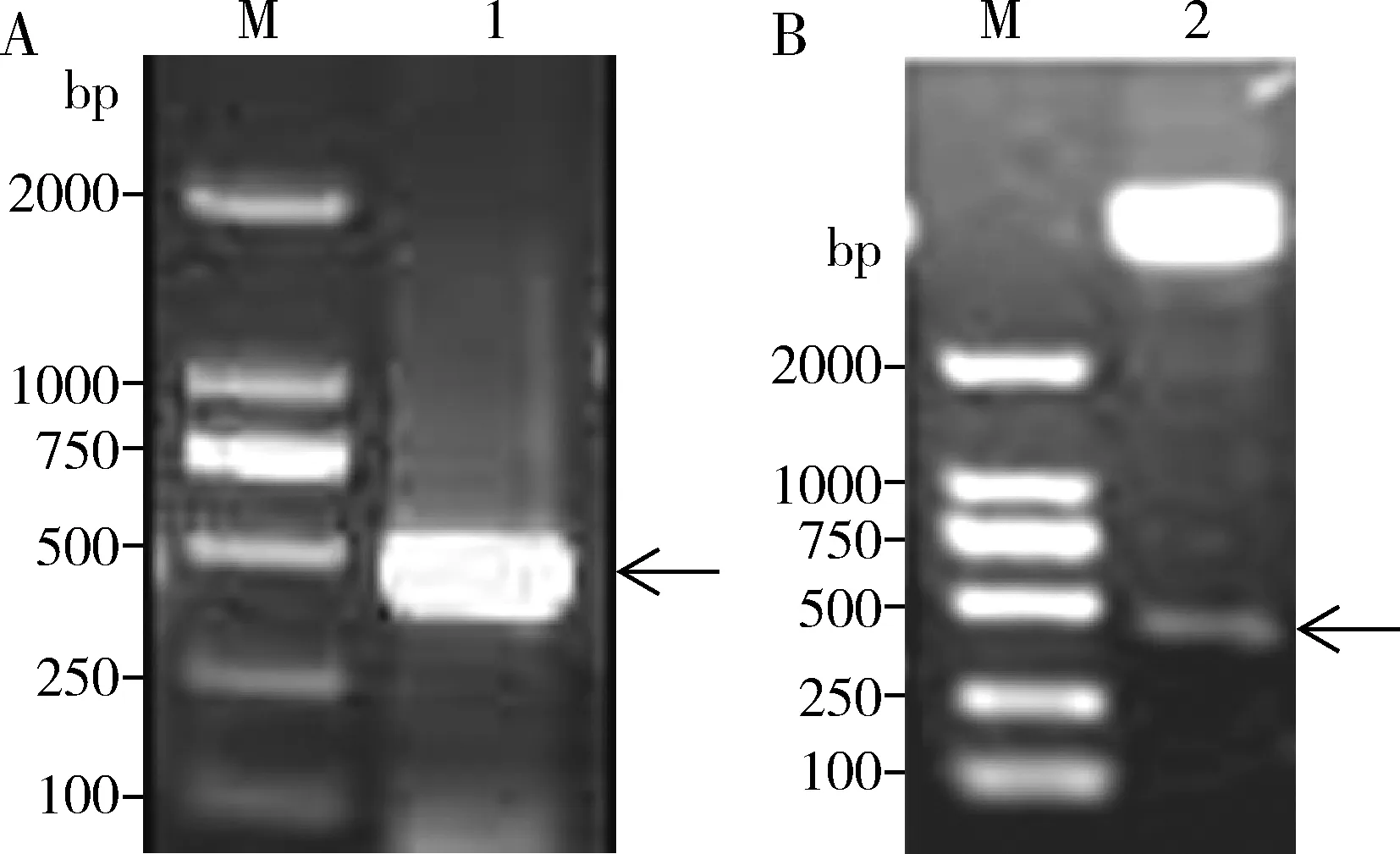
圖2 pET-28a/AdisOBP6重組質粒PCR(A)和酶切鑒定(B)
2.3 AdisOBP6重組蛋白在大腸桿菌中的誘導表達
為了達到理想的表達效果,我們采用小樣分別在20℃和37℃條件下用IPTG(0.5 mmol/L)誘導。從圖3中可知,在20℃條件下,誘導效果最佳,目的蛋白表達量最高。在20℃和37℃條件下蛋白都大量表達在上清中,沉淀中很少,說明蛋白主要為可溶性蛋白。
2.4 AdisOBP6蛋白純化
SDS-PAGE檢測結果表明,在20 mmol/L咪唑洗脫時出現單一清晰的目的條帶,隨著咪唑濃度增加條帶清晰度越來越低(圖4: A);將透析和濃縮后的純化蛋白進行SDS-PAGE電泳檢測結果顯示,AdisOBP6蛋白條帶清晰,雜帶少,說明蛋白純化效果較好(圖4: B)。
2.5 AdisOBP6抗體特異性
ELISA法測定抗體的效價,吸光值隨著抗體稀釋倍數的增加逐漸降低;4次重復的抗體分別稀釋到512 000, 512 000, 512 000和64 000倍時,抗體效價值≥2.5×陰性值。為了鑒定所制備的AdisOBP6抗體的特異性,以Western blot法檢測抗體對AdisOBP6重組蛋白的識別(圖5),結果顯示,重組蛋白與AdisOBP6抗體很好地結合在一起,說明該抗體確實為AdisOBP6抗體,故所制備的抗體可用于AdisOBP6功能的相關研究。

圖3 重組蛋白AdisOBP6的SDS-PAGE電泳檢測
2.6 AdisOBP6基因在mRNA水平和蛋白質水平上的表達譜
熒光定量PCR結果顯示,AdisOBP6在雙委夜蛾精巢中的表達量遠高于在觸角中的,在幼蟲期的精巢中AdisOBP6就已經開始表達,蛹期表達量顯著低于幼蟲期的(P<0.05)。成蟲羽化后,AdisOBP6在精巢中的表達量逐漸進入高峰。未受精卵和受精卵中AdisOBP6幾乎不表達或表達量極低(圖6: A)。
Western blot檢測分析發現,AdisOBP6蛋白主要在精巢中表達,在雌雄成蟲觸角、受精卵和未受精卵中表達量很低。在幼蟲精巢中AdisOBP6蛋白就開始表達一直延續到成蟲期,在成蟲期精巢AdisOBP6的表達量達到高峰(圖6: B)。
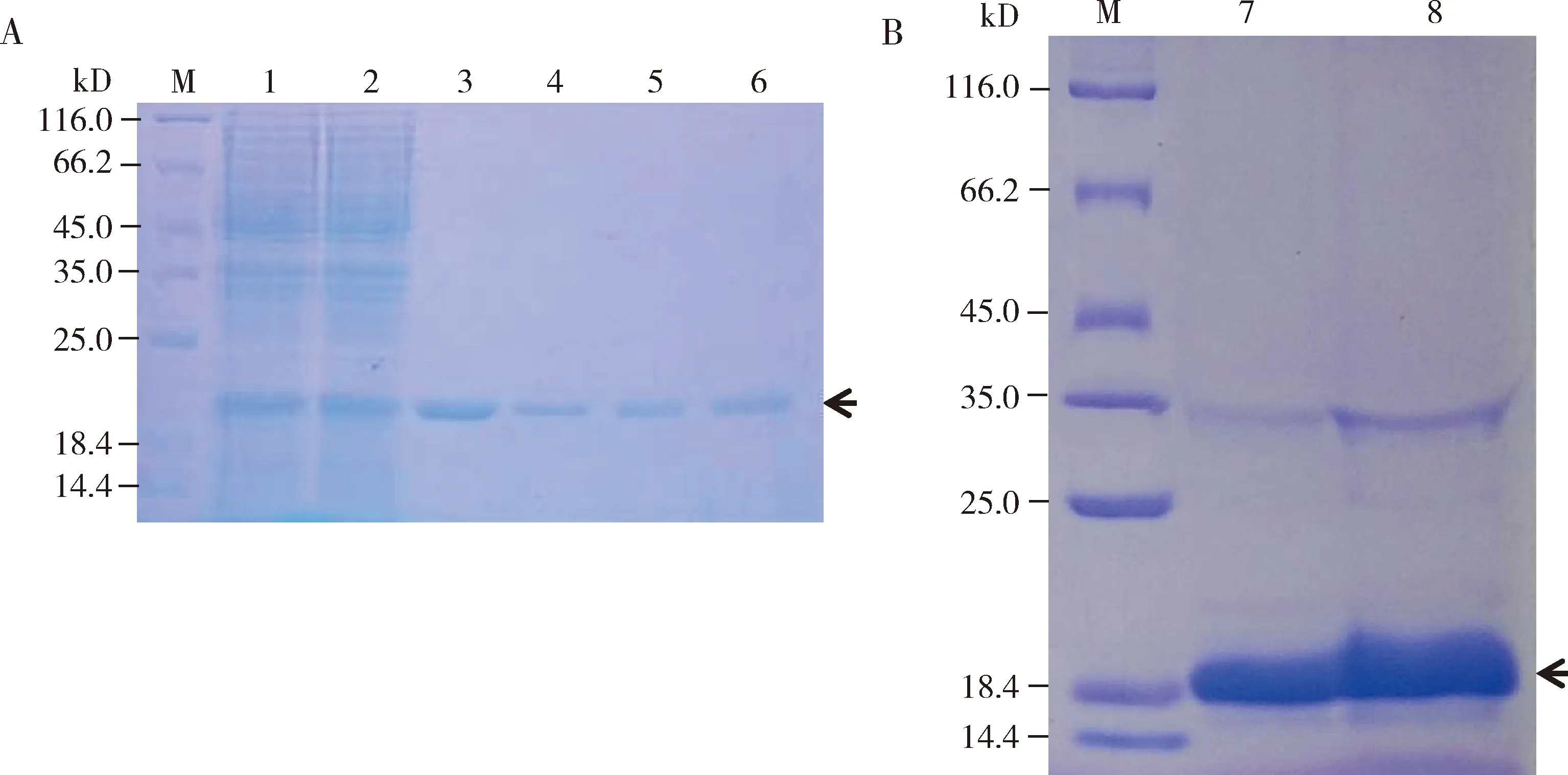
圖4 AdisOBP6蛋白純化(A)和濃縮后(B)的SDS-PAGE電泳檢測

圖5 Western blot檢測抗體對重組蛋白AdisOBP6的特異性
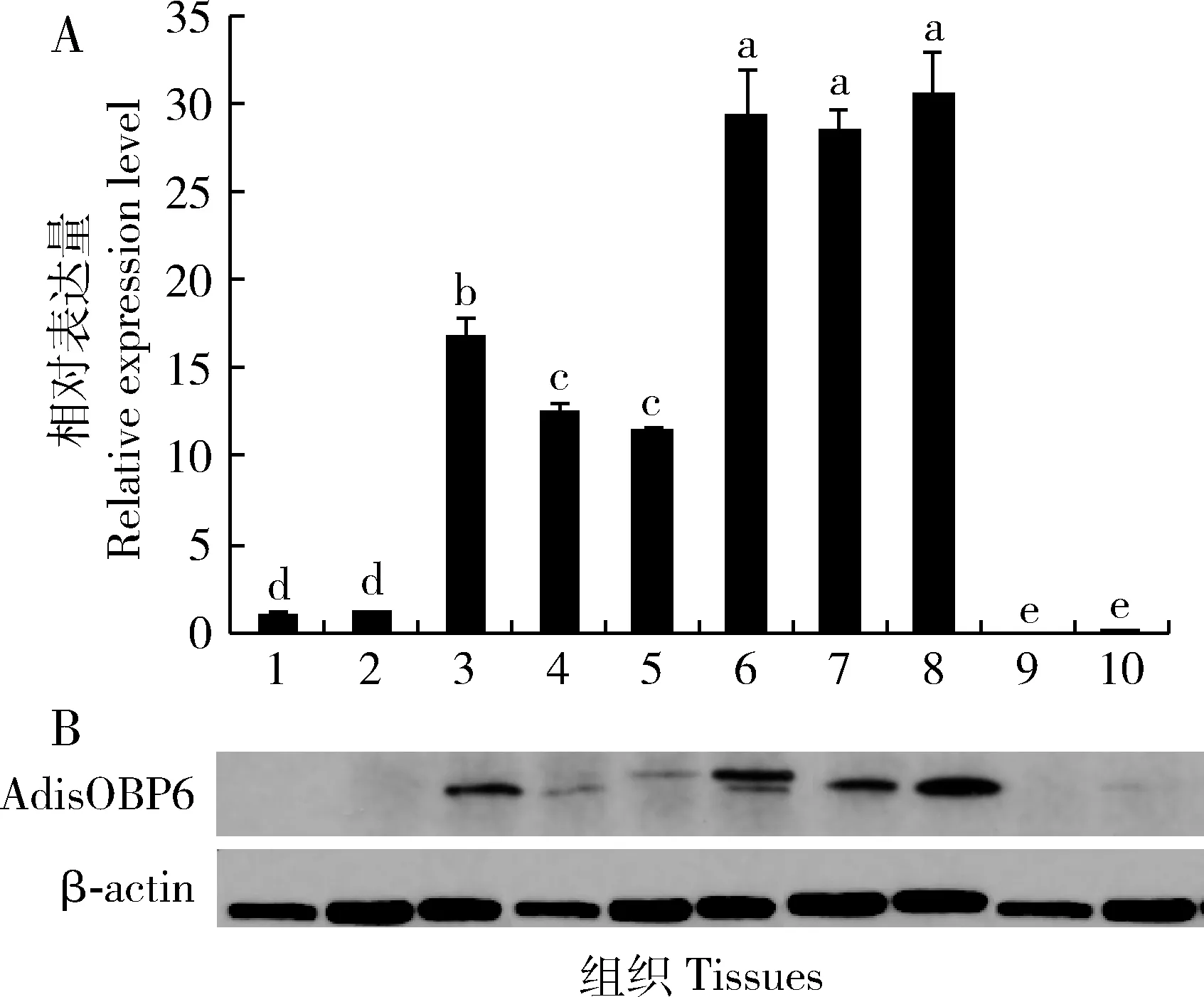
圖6 雙委夜蛾AdisOBP6基因在mRNA(A)和蛋白(B)水平上的表達譜
3 討論
目前獲得蛋白三維結構的方法主要是核磁共振(NMR)和X-射線衍射等方法,昆蟲多種氣味結合蛋白三維結構也被解析(Sandleretal., 2000; Lartigueetal., 2002; Leeetal., 2002; Pesentietal., 2009)。從昆蟲氣味結合蛋白三維結構來看,OBPs包含有6個α-螺旋,并命名為α1, α2, α3α, α4α, α5和α6,其中α1和α3, α3和α6, 以及α5和α6分別連接成3個二硫鍵支撐蛋白結構,使蛋白結構更穩定,并且圍成一個口袋形狀,將親水性氨基酸分布到口袋外面,疏水氨基酸分布在口袋內,形成疏水性口袋,主要用于結合一些親脂性氣味分子,將這些氣味分子轉運到受體上(Jiangetal., 2009; Xuetal., 2010)。采用同源建模的方法,特別是序列一致性較高時,能夠準確預測到OBPs的三維結構,促進OBPs功能的研究。我們通過序列比對,AdisOBP6與岡比亞按蚊AgamOBP4氨基酸序列一致性最高,并進行同源建模,預測了AdisOBP6的三維結構,發現AdisOBP6具有7個α-螺旋。昆蟲OBPs的構象變化具有pH依賴性(Xuetal., 2010; Tianetal., 2016),在pH酸性條件下,氨基酸序列的C端形成第7個α-螺旋占據結合腔釋放配體;pH中性時,第7個α-螺旋從結合腔排出,漏出結合腔內的疏水性殘基,重新結合氣味分子(Xu and Leal, 2008; Xuetal., 2010),此現象并非所有鱗翅目OBPs共有。
原核表達蛋白純化是直接關聯著后期蛋白功能的研究,當前對原核表達可溶性His標簽融合蛋白純化最常用的方法是鎳瓊脂糖親和層析法純化蛋白。該方法容易獲得高純度的蛋白,純化效率高,操作方便。然而在原核表達的時候經常出現包涵體,還需經過變性復性才能恢復生物活性(張庶民和祁自柏, 1995),操作復雜。因此在原核表達時,經常降低溫度,以便使蛋白盡量為可溶性蛋白。本研究采用不同溫度(20℃和37℃)進行原核表達,結果表明兩種溫度下都能成功表達出可溶性的AdisOBP6蛋白(圖3),且在20℃時蛋白表達量更高。此研究為后期進行結合實驗等蛋白功能研究奠定基礎。
昆蟲大多數OBPs主要表達在觸角中,也有少量OBPs在頭、口器、足、翅和中腸等非嗅覺器官中表達(Zhuetal., 2013; Liuetal., 2015; 鞏雪芳等, 2020),暗示它們可能還參與了除嗅覺以外其他的生理功能。例如,黑花蠅Phormiaregina的OBP56a在口器中高度表達,推測對膳食脂肪酸具有增溶作用(Ishidaetal., 2013);埃及伊蚊的OBP45在卵巢中大量表達,可能參與卵膜的形成(Costa-da-Silvaetal., 2013);中華蜜蜂Apisceranacerana的OBP15在足中特異性高度表達,推測可能在采集花粉和花蜜時用于味覺的識別(Duetal., 2019)。我們發現AdisOBP6基因在精巢中大量表達,并且在幼蟲期AdisOBP6就已經開始表達,蛹期表達量低于幼蟲期,成蟲期表達量逐漸進入高峰,未受精卵和受精卵中AdisOBP6幾乎不表達或表達量極低;Western blot印跡試驗也證實了,AdisOBP6蛋白在精巢中大量表達,但在觸角、受精卵和未受精卵中幾乎沒有檢測到AdisOBP6蛋白的存在(圖6),推測AdisOBP6可能參與了授精過程。同樣,埃及伊蚊的OBP22和棉鈴蟲、煙青蟲的OBP10也在精液中高度表達(Lietal., 2008; Sunetal., 2012),具體的生理功能只是推測,還需要進一步去證實。
本研究成功地純化出AdisOBP6可溶性蛋白,并制備了相應的抗體,同時針對AdisOBP6在mRNA水平和蛋白質水平上的表達情況進行了研究,為今后研究AdisOBP6在雙委夜蛾精巢內的具體生理功能奠定了基礎,同時也豐富了昆蟲氣味結合蛋白基因功能的認識。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
哲學評論(2021年2期)2021-08-22 01:53:34
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
海峽科技與產業(2016年3期)2016-05-17 04:32:12
