卵巢癌組織中SSBP1的表達(dá)意義及作用
2021-03-06 04:20:30康安靜
實(shí)用癌癥雜志 2021年2期
關(guān)鍵詞:研究
苗 麗 王 棟 馬 莉 康安靜 張 潔
卵巢癌(ovarian cancer)是嚴(yán)重威脅女性健康的常見惡性腫瘤之一,與其他婦科腫瘤相比,卵巢癌死亡率最高,術(shù)后晚期患者的五年生存率不足30%[1]。近年來靶向藥物的應(yīng)用為腫瘤治療提供了新思路。
Warburg 效應(yīng)在腫瘤發(fā)生及進(jìn)展中具有關(guān)鍵作用,其調(diào)控機(jī)制研究一直是腫瘤領(lǐng)域的熱點(diǎn)問題[2]。Warburg效應(yīng)重要特點(diǎn)就是線粒體氧化磷酸化減弱及糖酵解顯著增強(qiáng)。作為真核細(xì)胞中唯一具有獨(dú)立基因組的細(xì)胞器,線粒體在能量代謝、鈣穩(wěn)態(tài)調(diào)節(jié)、活性氧產(chǎn)生和凋亡調(diào)控等細(xì)胞生理過程中發(fā)揮著重要作用。線粒體數(shù)量和功能異常參與了包括腫瘤在內(nèi)的多種人類疾病的發(fā)生與進(jìn)展。例如研究表明,線粒體DNA拷貝數(shù)異常與肝癌、結(jié)直腸癌和膠質(zhì)瘤等腫瘤發(fā)病風(fēng)險(xiǎn)和生存顯著相關(guān)[3-4]。但關(guān)于腫瘤中線粒體DNA復(fù)制及線粒體拷貝數(shù)增加的具體誘因及機(jī)制尚不明確。線粒體單聯(lián)DNA結(jié)合蛋白(SSBP1,又稱mtSSB)具有促進(jìn)線粒體DNA復(fù)制和損傷修復(fù)進(jìn)而調(diào)控線粒體生成的重要作用[5]。近年來,SSBP1在腫瘤中作用受到越來越多的關(guān)注。本研究通過從SSBP1在卵巢癌組織中的表達(dá)變化入手,通過從TCGA數(shù)據(jù)庫(kù)中選取正常卵巢及卵巢癌組織的轉(zhuǎn)錄組數(shù)據(jù),利用Kaplan-Meier模型分析SSBP1與患者生存情況,并進(jìn)一步研究SSBP1對(duì)卵巢細(xì)胞惡增殖的影響,以期為闡明SSBP1在卵巢癌發(fā)生發(fā)展中的作用提供思路和線索。
1 材料與方法
1.1 數(shù)據(jù)來源
從TCGA 數(shù)據(jù)庫(kù)(https://tcgadata.nci.nih.gov/tcga/)下載正常卵巢組織轉(zhuǎn)錄組數(shù)據(jù)97例以及有預(yù)后信息的373例卵巢癌數(shù)據(jù)集(年齡30~87歲,中位年齡59歲);并利用Bioconductor/TCGAbiolinks 函數(shù)包提取mRNA表達(dá)RNASEqV2數(shù)據(jù)。基因的表達(dá)量由Fregments Per Kilobase per Million(FPKM)表示。計(jì)算方法為FPKM=106×nf/L/N。nf是比對(duì)至目標(biāo)基因的片段(fragment)數(shù)量、L是目標(biāo)基因的外顯子長(zhǎng)度之和除以1000(L單位為kb)、N 是總有效比對(duì)至基因組的讀取次數(shù)(read數(shù)量)。
1.2 細(xì)胞培養(yǎng)及轉(zhuǎn)染
人卵巢癌細(xì)胞SKOV-3購(gòu)于ATCC細(xì)胞庫(kù),細(xì)胞采用含10%胎牛血清(四季青生物)的RPMI-1640培養(yǎng)基(上海生工生物)培養(yǎng),培養(yǎng)箱含5%濃度的CO2,溫度設(shè)定為37 ℃。
取對(duì)數(shù)期卵巢癌細(xì)胞系SKOV-3,胰蛋白酶消化后用無雙抗 10﹪血清培養(yǎng)液制成細(xì)胞懸液,接種到35 mm培養(yǎng)皿并培養(yǎng)至70%匯合率,進(jìn)行轉(zhuǎn)染。取出培養(yǎng)皿,吸去培養(yǎng)皿中的培養(yǎng)液,并每皿加入DMEM培養(yǎng)液約0.5 ml重復(fù)沖洗細(xì)胞兩遍,而后每皿加入 DMEM 培養(yǎng)液1.5 ml,轉(zhuǎn)染復(fù)合物混合液0.5 ml。 輕輕混勻,放入37 ℃、5% CO2孵育箱中培養(yǎng)6 h左右后更換培養(yǎng)基。用G418篩選穩(wěn)定細(xì)胞株。
1.3 Western Blot
首先,用RIPA蛋白裂解液裂解卵巢癌細(xì)胞并提取細(xì)胞總蛋白,BAC法對(duì)蛋白濃度進(jìn)行測(cè)定后加入2×上樣緩沖液并置于沸水中煮沸5 min。隨后進(jìn)行聚丙烯酰胺凝膠電泳,并用濕轉(zhuǎn)法將凝膠中蛋白轉(zhuǎn)印至PVDF膜上,用5%脫脂奶粉室溫封閉1 h后,分別加入用封閉液稀釋過的SSBP1與actin抗體于4 ℃反應(yīng)過夜,TPBS洗膜3次后加入二抗并在室溫下繼續(xù)反應(yīng)1.5 h,繼續(xù)用TPBS洗膜3次后用化學(xué)發(fā)光法對(duì)最終條帶進(jìn)行檢測(cè)。
1.4 線粒體定量
去除細(xì)胞培養(yǎng)液,加入配制好的并37 ℃預(yù)溫育的Mito-Tracker Red染色工作液,與細(xì)胞37 ℃共孵育15 min。 去除Mito-Tracker Red染色工作液,加入37 ℃預(yù)溫育的新鮮細(xì)胞培養(yǎng)液。隨后通常用熒光顯微鏡進(jìn)行觀察分析。
1.5 乳酸及氧氣消耗定量
依照索萊寶乳酸定量試劑盒(貨號(hào):BC2235)方法:收集5×106個(gè)細(xì)胞加入1 ml提取液,冰浴超聲波破碎細(xì)胞(功率300 w,超聲3秒,間隔7秒,總時(shí)間3 min),隨后4 ℃條件下12 000 g離心10 min取上清應(yīng)用試劑盒測(cè)定。細(xì)胞氧耗檢測(cè)采用氧電極法(Strathkelvin),信號(hào)采集間隔時(shí)間為0.5秒。
1.6 統(tǒng)計(jì)方法
應(yīng)用SPSS 19.0統(tǒng)計(jì)學(xué)軟件進(jìn)行數(shù)據(jù)分析。生存分析采用Kaplan-Meier法。為了選擇最佳FPKM截?cái)嘀?Best FPKM cut-offs)對(duì)患者進(jìn)行最顯著分組,本研究使用所有處于20~80%的FPKM值對(duì)患者進(jìn)行分組,檢查各組生存率的差異,并選擇產(chǎn)生最低log-rankP值的分組方式。利用Person函數(shù)對(duì)兩個(gè)變量之間的相關(guān)性開展分析。P<0.05為差異具有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
2.1 SSBP1在卵巢癌中的表達(dá)
TCGA數(shù)據(jù)集中正常卵巢組織中SSBP1的表達(dá)量最高為33.8 FPKM,最低為18.4 FPKM,平均數(shù)為26.5 FPKM。而在卵巢癌組織中SSBP1的表達(dá)量最高為58.2 FPKM,最低為5.3 FPKM,平均數(shù)為17.5 FPKM。統(tǒng)計(jì)分析可見,腫瘤組織中SSBP1表達(dá)水平顯著低于正常卵巢組織(P<0.001),見圖1。
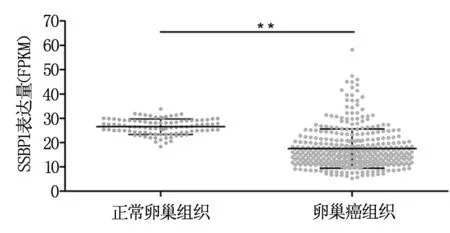
圖1 SSBP1在正常卵巢組織及卵巢癌組織中的表達(dá)分析
2.2 SSBP1表達(dá)水平與卵巢癌患者預(yù)后
利用373卵巢癌患者的轉(zhuǎn)錄組測(cè)序及預(yù)后數(shù)據(jù)的進(jìn)行Kaplan-Meier Plotter生存分析得出,低表達(dá)SSBP1患者的預(yù)后顯著差于高表達(dá)患者(P<0.05,圖2)。進(jìn)而提示,腫瘤組織中高水平的SSBP1是預(yù)后良好因素,低表達(dá)或不表達(dá)與患者生存期縮短顯著相關(guān)。
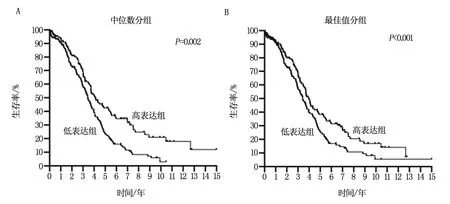
圖2 SSBP1表達(dá)水平與卵巢癌患者預(yù)后
2.3 SSBP1過表達(dá)對(duì)細(xì)胞糖代謝能力的影響
進(jìn)一步研究發(fā)現(xiàn),在卵巢癌細(xì)胞SKOV-3中穩(wěn)定過表達(dá)SSBP1(圖3A)可顯著增加線粒體含量及細(xì)胞氧耗速率(圖3B和3C),抑制糖酵解(3D),提示卵巢癌細(xì)胞中SSBP1的降低可促進(jìn)糖酵解、抑制線粒體氧化磷酸化,進(jìn)而促進(jìn)Warburg效應(yīng)。
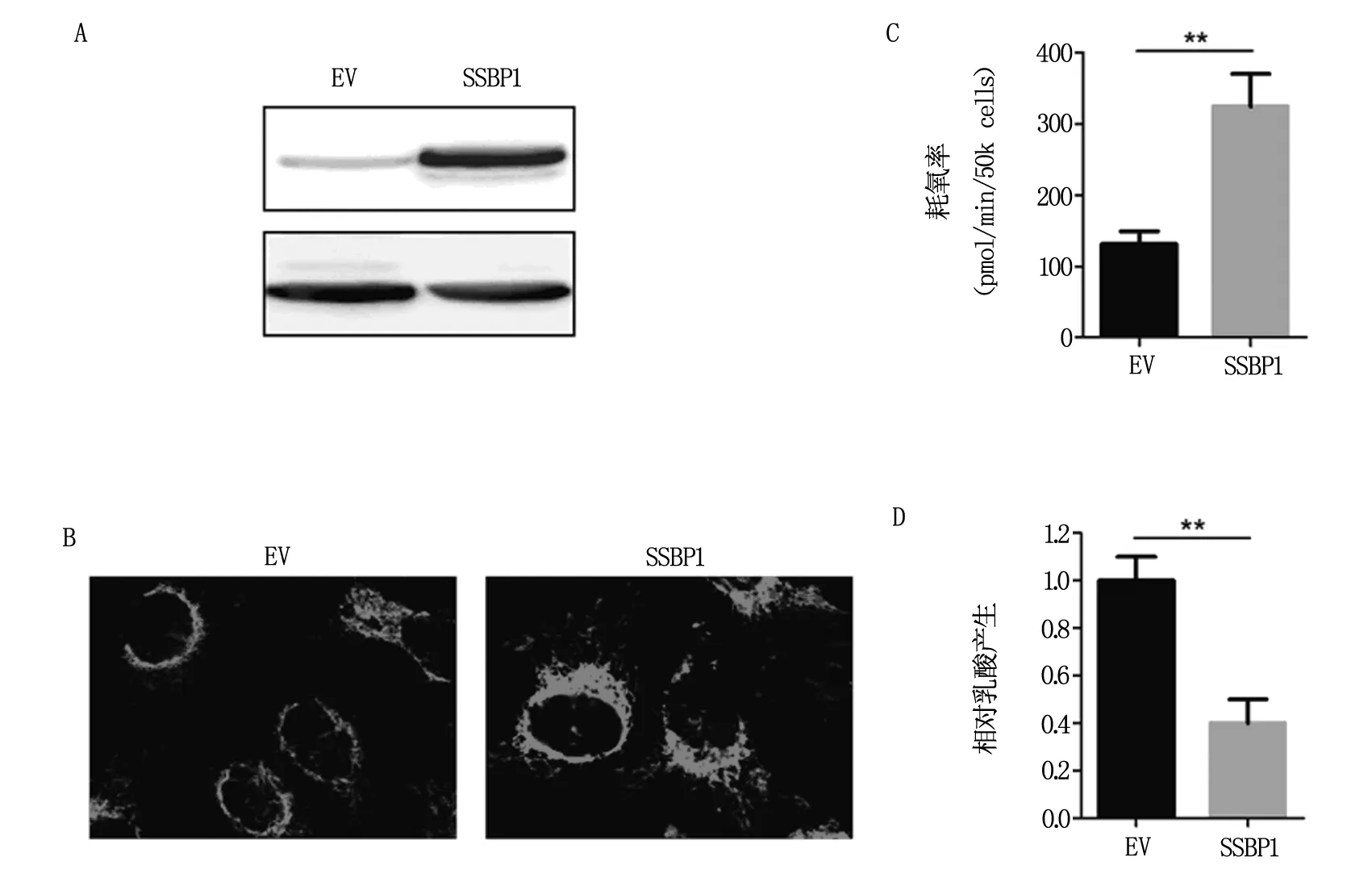
圖3 SSBP1過表達(dá)對(duì)細(xì)胞糖代謝能力的影響
2.4 SSBP1過表達(dá)對(duì)卵巢癌細(xì)胞生長(zhǎng)的影響
如圖4所示,過表達(dá)SSBP1可顯著抑制結(jié)卵巢癌細(xì)胞的生長(zhǎng),提示腫瘤內(nèi)低表達(dá)SSBP1與較較差預(yù)后可能存在因果關(guān)系。
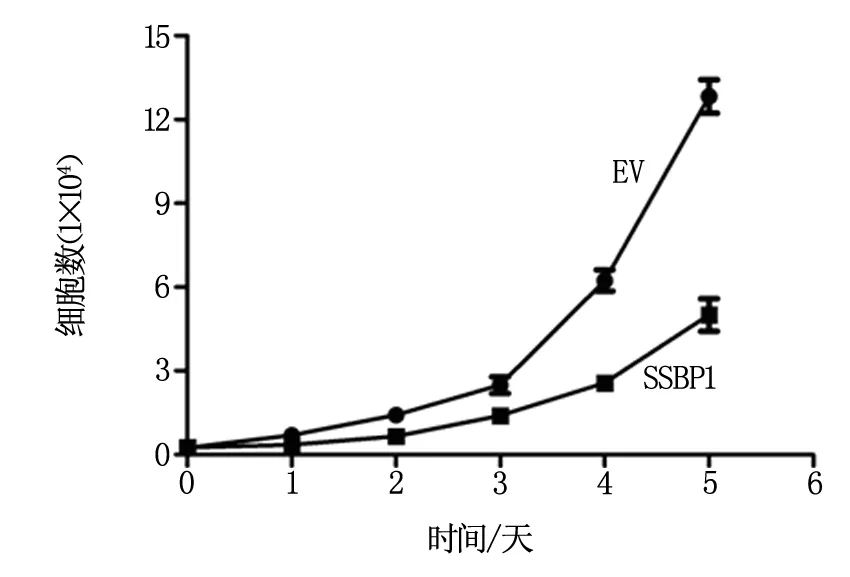
圖4 SSBP1過表達(dá)對(duì)卵巢癌細(xì)胞生長(zhǎng)的影響
3 討論
mtSSBP1在線粒體DNA(mtDNA)的復(fù)制、轉(zhuǎn)錄及修復(fù)過程發(fā)揮關(guān)鍵作用。因此,mtSSBP1突變或表達(dá)失調(diào)將導(dǎo)致mtDNA發(fā)生基因突變,最終可能會(huì)導(dǎo)致腫瘤的發(fā)生。在本研究中,我們首次發(fā)現(xiàn)了SSBP1在卵巢癌組織中的表達(dá)顯著低于正常卵巢組織,且腫瘤組織中低表達(dá)SSBP1的患者預(yù)后顯著差與高表達(dá)組患者。與之類似,Li等發(fā)現(xiàn)SSBP1基因多態(tài)性(rs6976500G)可顯著降低SSBP1的轉(zhuǎn)錄活性及表達(dá)水平,進(jìn)而與胃癌的不良預(yù)后顯著相關(guān)[6]。進(jìn)一步Jiang等研究發(fā)現(xiàn),三陰性乳腺癌組織中SSBP1的表達(dá)下調(diào)可通過“線粒體-核交互信號(hào)通路”激活上皮-間質(zhì)轉(zhuǎn)換(EMT)過程,繼而促進(jìn)腫瘤細(xì)胞的轉(zhuǎn)移[7]。Wang等發(fā)現(xiàn)在非小細(xì)胞肺癌中SSBP1表達(dá)下降、mtDNA拷貝數(shù)降低,并導(dǎo)致腫瘤細(xì)胞的放療敏感性升高[5]。此外,Ye等還利用蛋白質(zhì)印跡及免疫組化等方法對(duì)20例肝癌組織的進(jìn)行分析發(fā)現(xiàn),與對(duì)照組相比,SSBP1在腫瘤組織中表達(dá)異常上調(diào),推測(cè)可能是由于mtSSBP1蛋白不能正確折疊而導(dǎo)致其在細(xì)胞中的異常累積,并可能造成了mtDNA的復(fù)制障礙及突變[8]。以上研究結(jié)果都提示SSBP1的表達(dá)異常可通過抑制mtDNA復(fù)制及線粒體正常功能,進(jìn)而促進(jìn)腫瘤的發(fā)生進(jìn)展。
正常細(xì)胞通過線粒體氧化磷酸化產(chǎn)生ATP 提供能量,而腫瘤細(xì)胞則表現(xiàn)出了不同的能量代謝方式。因此,糖代謝重編程是腫瘤細(xì)胞的一個(gè)重要特征,其與腫瘤的發(fā)生發(fā)展互為因果。早在上個(gè)世紀(jì),德國(guó)生理學(xué)家Otto.Warburg就提出了著名的Warburg效應(yīng):相比于正常細(xì)胞,腫瘤細(xì)胞糖酵解顯著增強(qiáng),線粒體氧化磷酸化降低來滿足快速生長(zhǎng)的需求[9]。在很長(zhǎng)一段時(shí)間內(nèi),科研工作者一直認(rèn)為腫瘤細(xì)胞有氧呼吸存在不同程度的損傷,即線粒體功能的不可逆性損傷是所有腫瘤的共同起因。然而,進(jìn)來越來越多的研究表明絕大部分腫瘤細(xì)胞中線粒體數(shù)目下降。在本研究中我們發(fā)現(xiàn)SSBP1的表達(dá)下調(diào)可減少線粒體數(shù)目,并促使卵巢癌細(xì)胞糖酵解與增殖能力的增強(qiáng)。與之相類似,Huang等發(fā)現(xiàn)肝癌中高表達(dá)的CD147分子可通過抑制p53通路下調(diào)線粒體生物生成的關(guān)鍵調(diào)控分子PGC-1α、TFAM等進(jìn)而導(dǎo)致腫瘤細(xì)胞線粒體數(shù)目減少,與腫瘤細(xì)胞的增殖[10]。
上述研究結(jié)果提示,線粒體數(shù)目有望成為1種新型標(biāo)志物用于卵巢癌患者預(yù)后評(píng)估及藥物治療靶點(diǎn)。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國(guó)際商務(wù)財(cái)會(huì)(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19

- 實(shí)用癌癥雜志的其它文章
- 惡性腫瘤合并心房顫動(dòng)對(duì)抗凝治療后患者血栓栓塞和出血發(fā)生的影響
- 兒童神經(jīng)母細(xì)胞瘤化療后嚴(yán)重感染的影響因素及重組人粒細(xì)胞刺激因子的預(yù)防效果
- 兩種門冬酰胺酶治療兒童急性淋巴細(xì)胞白血病的近期效果及對(duì)患兒血清LDH、SF和ESR的影響
- 伊立替康聯(lián)合紫杉醇和卡鉑治療卵巢癌的療效及對(duì)患者血清CA199、CEA及CA242水平的影響
- 宮頸癌患者PICC導(dǎo)管多重耐藥菌感染的危險(xiǎn)因素及病原學(xué)特征
- 足月妊娠合并不同臨床分期宮頸癌患者母嬰結(jié)局的差異性研究