色氨酸調控畜禽熱應激研究進展
2021-03-03 02:51:10李秋粉歐陽經鑫黎觀紅
動物營養學報 2021年2期
關鍵詞:氧化應激
李秋粉 歐陽經鑫 黎觀紅
(江西農業大學動物科學技術學院,江西省動物營養重點實驗室,南昌 330045)
隨著全球氣候變暖趨勢的加劇,熱應激已成為影響畜牧業發展的重要因素之一。反芻動物、豬、家禽是恒溫動物,在熱中性區通過自身體溫調節機制維持體溫,當環境溫度達到動物等熱區上限,超過機體自身調節能力時,引發一系列非特異性免疫應答反應,導致畜禽熱應激,從而增加畜禽養殖成本、損害畜禽健康、降低生產力和產品品質,最終造成經濟損失。
作為畜禽機體功能性必需氨基酸之一,色氨酸(Trp)及其代謝產物可以提高采食量、增加機體蛋白質沉積和消化吸收功能、提高生長性能、增強免疫功能、緩解腸道黏膜屏障損傷、提高機體抗應激能力[1-4]。本文從熱應激的產生機理和對畜禽的影響、色氨酸的代謝及其代謝產物生理功能和色氨酸及其代謝產物對熱應激的可能調節機制進行綜述,討論色氨酸及其代謝產物在熱應激條件下與機體免疫的交叉點,為色氨酸在調節畜禽熱應激的應用研究提供新的思路。
1 熱應激的產生機理及對畜禽的影響
1.1 熱應激的產生機理
當外界溫度高于機體調節上限,畜禽不能通過自身神經內分泌系統維持正常體溫,動物便處于熱應激狀態。在熱應激環境中,畜禽機體最初通過增加水攝入量、出汗和呼吸頻率,降低心率和采食量等行為和生理反應的啟動,將體溫維持在正常范圍內[5];隨后,下丘腦-垂體-腎上腺皮質(HPA)軸和交感神經-腎上腺髓質(SAM)軸的激活,導致循環中的兒茶酚胺和皮質醇含量升高;隨著熱應激時間的持續,細胞和體液免疫功能遭到損傷,影響消化、生長、生殖和其他代謝過程[6-7]。
1.2 熱應激對畜禽生產和免疫的影響
動物處于熱應激狀態,機體難以通過增加飲水、呼吸頻率和降低采食量等維持恒定體溫時,首先表現為體溫升高、生產性能降低[8-9];免疫反應的中樞神經系統(CNS)是一個復雜的網絡系統,包括神經內分泌系統和免疫系統,HPA軸和SAM軸調節是免疫調節的2個主要途徑。有研究表明,在熱應激條件下,HPA軸激活,導致循環中的兒茶酚胺和皮質醇含量升高,其中急性熱應激比慢性熱應激誘導糖皮質激素水平更高,并引起一系列生化反應,導致脫水,影響能量消耗和蛋白質代謝[6,10]。熱應激會增加活性氧(ROS)的產生,誘導氧化應激,造成肝臟、腸道等器官氧化損傷,破壞腸道黏膜完整性,損害畜禽健康和降低生產性能[11-12]。腸道是機體最大的消化吸收器官,也是重要的防御屏障。在發揮吸收功能的同時,健康腸道的黏膜屏障保護宿主免受具有致病力的毒素和微生物的侵襲。腸道微生物群與腸黏膜免疫密切相關,可同時發揮促炎和抗炎作用。眾多研究發現,持續的熱應激會導致畜禽腸道的菌群結構和多樣性紊亂、有益菌比例降低而有害菌比例增高、黏膜屏障損傷、機體生長性能及免疫力下降[13-15]。
2 色氨酸對熱應激的影響
2.1 色氨酸代謝和生理功能
色氨酸是構成蛋白質的20種氨基酸之一,是所有動物的必需氨基酸,同時也是生成血清素[如5-羥色胺(5-HT)]、褪黑激素、色胺等一些重要的生物活性分子的前體。色氨酸的代謝途徑主要有3種(圖1),其中2種是宿主途徑,分別是犬尿氨酸代謝途徑和5-HT代謝途徑,犬尿氨酸代謝途徑限速酶為吲哚胺2,3-雙加氧酶(IDO)和色氨酸2,3-雙加氧酶(TDO)[16],5-HT代謝途徑限速酶主要為腸嗜鉻細胞中的色氨酸羥化酶1(TPH1)[17];另一種是微生物代謝途徑,也就是吲哚途徑,其中一些代謝產物可以作為芳香烴受體(AhR)配體。除了色氨酸的直接代謝外,微生物群和微生物代謝物對宿主色氨酸的2種途徑也有重要的調節作用[18-20]。研究表明,哺乳動物在感染、應激狀態下,大多數色氨酸通過犬尿氨酸途徑被分解轉化為可能與應激反應相互作用的生物活性物質[21-23]。而急性應激對哺乳動物的大腦色氨酸含量的調節有相反的影響[24],這種腦色氨酸含量增加可能與交感神經激活和血漿兒茶酚胺含量升高有關[25]。陶絲雨等[26]的試驗亦表明,腸易激綜合征提高結腸TPH1基因表達以及5-HT的含量。那么熱應激狀態下畜禽機體色氨酸代謝又是怎樣的?是否是機體發生急性熱應激使大腦色氨酸含量增加,5-HT含量升高,抑制血漿皮質醇含量升高,通過HPA軸緩解應激反應;而隨著熱應激的持續,色氨酸代謝轉向犬尿氨酸代謝途徑調控熱應激導致的氧化應激和炎癥反應,尚不明確,因此其具體代謝途徑還需進一步試驗研究。
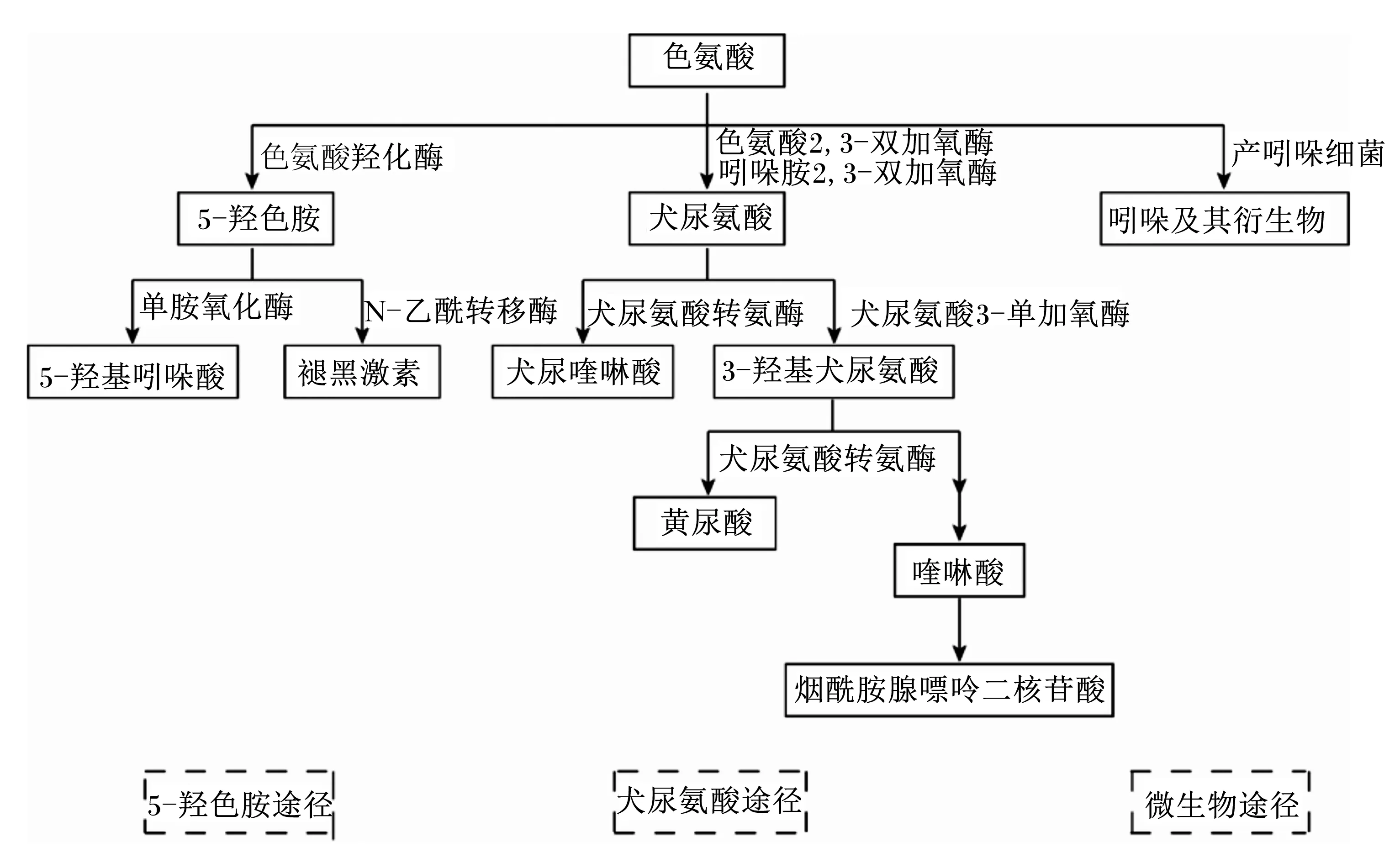
圖1 色氨酸主要代謝途徑
研究表明,除了作為起始氨基酸參與蛋白質合成外,色氨酸及其代謝產物在調節消化吸收、能量代謝、蛋白質沉積、免疫功能和應激反應等方面有重要作用[28-30]。5-HT是由色氨酸通過色氨酸羥化酶和芳香氨基酸脫羧酶兩級酶促反應生成的。在動物體內,血清素主要存在于胃腸道、血小板和中樞神經系統,參與HPA軸調節,促進腸道蠕動[31-32]。另外,5-HT還能通過調節免疫細胞中白細胞介素-6(IL-6)和白細胞介素-10(IL-10)產生影響機體免疫應答[3]。褪黑激素是由動物松果腺分泌,其主要作用是調節晝夜節律,但也被證明影響包括免疫功能、凋亡、增殖和氧化應激在內的多種分子途徑[33-34]。犬尿氨酸是犬尿氨酸代謝途徑的中間產物,具有抗菌活性,可直接影響腸道菌群的增殖[35],在犬尿氨酸代謝中,犬尿氨酸(KYN)、犬尿喹啉酸(KA)和黃尿酸(XA)等作為AhR的直接配體,以濃度依賴的方式刺激AhR和AhR依賴的基因表達,調節腸道內穩態,AhR自身也在調節IDO1和TDO1表達量方面發揮作用[36]。因此,AhR可能是腸道微生物群、犬尿氨酸代謝途徑與免疫應答之間復雜相互作用的重要中介物質。此外,跨膜G蛋白偶聯受體(GPCRs)也能感受其代謝中間體,激活信號通路,調節胃腸道穩態和腸道免疫[28]。煙酰胺腺嘌呤二核苷酸(NAD)是生物氫的載體,作為輔酶,在糖酵解、檸檬酸循環、三羧酸循環、糖異生和電子傳遞等方面發揮重要作用。吲哚及其衍生物是在色氨酸經多種產吲哚細菌產生的色氨酸酶作用下產生的,包括吲哚、吲哚乙醇(IE)、吲哚丙酸(IPA)、吲哚乙酸(IAA)、吲哚醛(IAld)和吲哚丙烯酸(IA)等,這些物質對宿主的生理機能有多種影響。吲哚、IPA和IA通過降低腸通透性影響黏膜內平衡。吲哚還可以誘導腸內分泌細胞釋放胰高血糖素樣肽1(GLP1),調控食欲、胰島素分泌和延緩胃排空[37]。
2.2 色氨酸及其代謝產物對熱應激的影響
飼糧添加色氨酸可緩解畜禽熱應激狀態。給閹公牛每千克體重靜脈滴注38.5 mg色氨酸,可通過增加腦5-HT含量減弱牛在急性熱應激環境中直腸溫度的升高[38]。在豬研究上,色氨酸亦可提高下丘腦5-HT含量和降低血漿皮質醇含量,通過HPA軸緩解應激反應[39-41]。熱應激常伴隨嚴重的氧化應激,飼糧中補充0.15%色氨酸能減輕敵草快注射引起的仔豬腸道損傷、氧化應激和線粒體功能障礙[42]。Zimbelman等[43]研究表明,哺乳期奶牛補充膠囊狀的尼克酸會增加熱應激狀態下的皮膚表面蒸發熱損失,降低陰道溫度。相似的,王雪瑩等[44]在牦牛熱應激條件下飼喂煙酸,發現可以通過緩解HPA軸應激反應,調節機體免疫反應,增加血氧供給,提高其對熱應激環境的抵抗,提高采食量和營養物質消化率,減少生長性能由于熱應激引起的降低,添加煙酸增加煙酰胺腺嘌呤二核苷酸含量可能是其原因之一。褪黑激素是色氨酸代謝產物之一,有研究表明,褪黑激素增加熱應激處理后白細胞、單核細胞、粒細胞等免疫細胞數量,清除機體氧自由基,增加超氧化物歧化酶(SOD)、谷胱甘肽過氧化物酶(GSH-Px)等抗氧化酶活性,調節免疫功能,緩解熱應激,緩解體重下降程度[45-46]。也有研究表明,飼糧中添加0.75%的高水平色氨酸顯著提高仔豬血清二胺氧化酶(DAO)和D-乳酸含量,降低空腸中咬合蛋白(Occludin)和緊密連接蛋白-1(ZO-1)的基因表達,損傷腸道黏膜完整性[47]。因此,熱應激條件下各個物種、各個日齡階段的色氨酸添加量如何仍需試驗進一步研究探討。
3 色氨酸調控畜禽熱應激可能的作用機制
腸道微生物影響免疫系統響應;反過來,免疫系統亦參與調節腸道菌群的定位和組成。微生物與宿主免疫系統之間的聯系是由一系列分子和信號因子介導,繼而影響腸道、肝臟、大腦和其他器官。熱應激影響腸道菌群組成和多樣性,以及腸道菌群與免疫途徑相互作用。色氨酸是動物必需的營養物質,其代謝物參與腸道免疫穩態和多種免疫疾病的發生。本文從色氨酸及其代謝物與調節HPA軸,色氨酸及其代謝物與線粒體ROS產生和氧化應激相關信號通路,色氨酸及其代謝產物與腸道黏膜屏障功能3個方面概述色氨酸及其代謝產物與熱應激和腸道黏膜屏障功能研究概況,探討色氨酸及其代謝產物調控畜禽熱應激可能的作用機制。
3.1 色氨酸與畜禽HPA軸
在熱應激狀態下,HPA軸和SAM軸被激活,維持體內平衡[48]。其中,去甲腎上腺素(NOR)和5-HT參與HPA軸的激活和促腎上腺皮質激素(ACTH)的垂體釋放,HPA軸的激活會導致皮質醇分泌增加引起免疫反應,而NOR和5-HT與外周神經免疫激活有關,受白細胞介素-1(IL-1)、IL-6和腫瘤壞死因子-α(TNF-α)誘導[49]。在急性應激期,皮質醇的分泌對免疫系統起到刺激作用,而在慢性應激期,皮質醇的分泌與免疫抑制有關[50-51]。研究表明,灌胃色氨酸促進慢性應激小鼠下丘腦5-HT的合成并增強其抵抗應激能力[52];飼糧中添加色氨酸提高慢性不可預知溫和應激肉雞和小鼠TPH1的表達,提高外周血5-HT的含量,緩解機體應激反應[53]。5-HT通過調節下丘腦促腎上腺皮質激素釋放因子(CRF)的釋放進而對動物HPA軸發揮重要的調控作用[54-55],并調節行為和神經內分泌對應激的應答[56-57]。此外,應激會導致色氨酸代謝通路改變,而色氨酸代謝通路改變反過來影響機體應激反應,加之5-HT神經傳遞和HPA軸反應敏感性的個體差異在脊椎動物中是普遍存在的現象[58],因而各種脊椎動物飼糧中最佳色氨酸含量的確定仍有待進一步研究。
3.2 色氨酸與畜禽氧化應激相關信號通路
線粒體作為產生細胞內絕大多數活性氧的細胞器,在調節氧化應激的產生和反應中起著關鍵作用。與細胞核DNA相比,線粒體ROS產生水平高,且對損傷防御能力相對較差,線粒體DNA更容易受到氧化損傷[59]。大量研究表明,畜禽熱應激降低畜禽機體抗氧化能力,引發氧化應激[60-62]。熱應激是活性氧產生的強誘導劑,一旦由谷胱甘肽(GSH)、GSH-Px、SOD和血紅素氧化酶1(HO1)組成的細胞氧化防御系統耗盡,就會導致組織損傷[63]。抗氧化系統的表達主要受紅系衍生的核因子2相關因子2(Nrf2)的調控,在生理條件下,Nrf2在細胞質中被Kelch樣ECH相關蛋白1(Keap1)抑制。氧化應激時Nrf2從Keap1中解離,導致Nrf2轉位到細胞核中與抗氧化反應元件(ARE)結合,誘導抗氧化蛋白的轉錄,提高應激條件下的細胞存活率[64]。
熱應激雛雞血漿中鹽酸和喹啉酸含量升高,而煙酰胺含量降低。在哺乳動物中,色氨酸主要代謝為犬尿氨酸,然后進一步代謝為喹啉酸和煙酰胺,而在雞研究中,這些代謝過程尚未得到深入研究。此外,從功能的角度來看,犬尿氨酸通過激活芳基烴受體調節免疫系統[36,65]。喹啉酸通過激活N-甲基-D-天門冬胺酸受體發揮神經毒性作用,并誘導游離自由基的產生和不依賴于受體的氧化應激。然而,這些可能與環境溫度的有害影響有關的功能,亦未在雞中得到證實[65-66]。有研究表明,IA可使LPS刺激下IL-6和白細胞介素-1β(IL-1β)分泌減少,Nrf2-ARE通路激活,AhR激活,黏蛋白2(Muc2)基因表達上調,表明了IA抗氧化和抗炎功能。同時發現,IPA在小鼠巨噬細胞中顯示了抗炎作用,而在人類單個核細胞中卻沒有,這表明IPA誘導的信號通路在小鼠和人類細胞中并不保守[67]。在色氨酸代謝過程中,產生褪黑激素、5-HT和3-羥基犬尿氨酸等幾種內源性抗氧化代謝物。Liu等[68]研究表明,飼糧中色氨酸從0.18%增加到1.08%可以提高畜禽機體總抗氧化能力、GSH-Px和過氧化氫酶活性,從而提高機體抗氧化活性,控制ROS的形成。由此,我們推測色氨酸可能在降低機體線粒體ROS產生和調控氧化應激相關信號通路基因表達2個方面調控氧化應激。
3.3 色氨酸與畜禽腸道黏膜屏障功能
腸道除了具有消化吸收功能,也是機體重要的免疫器官。健康的腸腔黏膜發揮屏障功能將有害細菌、病毒和其他大分子有害物質與機體內環境分開。腸道黏膜屏障根據結構和功能可劃分為:機械屏障、化學屏障、免疫屏障和微生物屏障。
3.3.1 機械屏障
細胞間連接的完整性對上皮細胞層的正常功能至關重要,可將腸道內環境與外界隔離開來,形成緊密連接,使細胞緊密結合,控制腸道黏膜通透性。Occludin、閉合蛋白(Claudins)、連接黏附分子(JAMs)和ZO等是細胞間連接重要的結構和功能性蛋白[69]。熱應激導致腸道完整性受損和通透性增加,與緊密連接蛋白的表達和分布改變相關[70]。體外和體內研究均表明,吲哚通過增加參與維持上皮細胞結構和功能的基因表達增強細胞間的上皮緊密連接,降低腸道黏膜通透性,改善腸上皮屏障功能[28,37]。
3.3.2 化學屏障
黏蛋白、膽汁、溶菌酶和抗菌肽等是腸上皮細胞的主要化學成分,可以抑制有害細菌的黏附和定植,維持腸黏膜的完整性。熱應激損傷腸黏膜絨毛結構,降低堿性磷酸酶活性和抗菌肽含量,減少單位面積內杯狀細胞數量,使得黏蛋白分泌水平下降,損傷腸道化學屏障[13,71-72]。AhR和孕烷X受體(PXR)調節與腸道Muc2基因的表達有關的信號通路,缺乏PXR的小鼠表現出腸道滲漏和小腸中Muc2表達減少[28]。IA作為一種重要的色氨酸微生物源性代謝物,能有效激活小鼠結腸AhR和上調Muc2基因表達,進一步證實了AhR激活和杯狀細胞功能之間的聯系[67]。此外,吲哚也可以激活AhR信號,進而促進局部白細胞介素-22(IL-22)的產生,并進一步促進抗菌肽的分泌,保護機體免受致病性感染[73]。
3.3.3 免疫屏障
免疫屏障主要由腸黏膜相關淋巴組織及分泌型抗體構成,分泌型免疫球蛋白(sIgA)為黏膜免疫的主要抗體,防止有害微生物定植[74]。Toll樣受體(TLRs)、核苷酸結合寡聚化結構域(NOD)為廣泛表達于各種先天免疫細胞和非免疫細胞上的進化保守的膜受體,作為主要的傳感器,可以通過響應細菌、病毒等病原體相關的分子模式來啟動先天免疫應答[69]。炎癥的啟動由促炎細胞因子和抗炎細胞因子之間的不平衡激活。研究表明,熱應激狀態下過度產生促炎細胞因子,如TNF-α,導致包括腸道在內的器官出血和壞死[75]。AhR是一種胞質配體激活的轉錄因子,介導外源性代謝,是免疫和炎癥的關鍵調節因子,調節適應性免疫和黏膜屏障功能、腸道穩態的維持和癌變[28,76-77]。AhR對免疫穩態的促進作用主要有2種機制:首先,AhR介導先天淋巴樣細胞的激活,在腸道中產生IL-22,IL-22通過調節微生物組成,調節抗菌肽的釋放,影響免疫與菌群間的穩態平衡[78];其次,有證據表明,AhR通過調節上皮內淋巴細胞和先天淋巴樣細胞的發育而具有抗炎作用,使得這些細胞在抵御病原微生物滲透和促進腸道內穩態方面發揮重要作用[76]。KYN、KA、XA等幾種微生物代謝物作為AhR的配體對宿主免疫至關重要,特別是在保護黏膜免受炎癥感染方面[79]。AhR通過與配體結合,直接靶向調控炎癥過程中包括IL-6、IL-22、前列腺素G/H合酶2、血管內皮生長因子A、細胞色素P4501A1在內的基因[28]。其中,吲哚可以降低TNF-α介導的核因子-κB(NF-κB)激活,增加抗炎細胞因子IL-10的水平[80]。IPA可顯著提高LPS刺激后IL-10的生成,并降低TNF-α的生成[81]。吲哚-3-乙酸(I3A)至少可以通過2種方式直接調節肝細胞和巨噬細胞的炎癥反應,I3A作用于巨噬細胞,可減弱促炎細胞因子的釋放,誘導肝臟合成游離脂肪酸,進而刺激巨噬細胞;I3A作用于肝細胞,可減弱細胞因子介導的脂肪生成上調[82]。有研究表明,在人樹突狀細胞中,AhR信號能改變TLRs調節的反應,進一步表明色氨酸分解代謝物對細胞因子產生的影響可能依賴于AhR的激活[83]。此外,有報道稱,短鏈脂肪酸(SCFAs)增強AhR介導的基因表達[84]。在熱應激條件下,色氨酸及其代謝與腸道微生物代謝物的關系,色氨酸及其代謝物能否通過AhR影響TLRs、NLRs的表達,進而影響其下游因子在熱應激條件下的表達均還需進一步探討。
3.3.4 微生物屏障
腸道中存在大量與宿主共生的微生物,在調節宿主營養和代謝、刺激腸道成熟、發育、增殖和免疫穩態等方面發揮重要作用[64,85]。飲食、免疫反應、感染和抗生素的使用都會影響各種宿主腸道菌群結構和多樣性。除機體條件改變,環境高溫等外界應激刺激亦會引起菌群平衡改變,導致腸道病原體定植[86]和腸道炎癥反應[87]。微生物群的作用不僅局限于腸道,還向大腦發出信號,影響中樞神經系統炎癥,并參與神經精神疾病[88],Trp-AhR通路是沿微生物-內臟-腦軸傳遞信號的重要通路之一,在宿主免疫系統、腸道免疫耐受和腸道菌群之間平衡的維持中起著至關重要的作用[81]。微生物代謝產物如SCFAs和多種微生物色氨酸代謝產物可以激活腸道或肝臟中的AhR和AhR靶基因,AhR信號反過來又能影響小腸內的微生物組成,AhR是宿主-微生物通訊的重要調節因子,影響宿主的新陳代謝,調節免疫系統[28]。外源性補充色氨酸可優化益生菌在腸道感染或化學損傷過程中抵抗有害細菌入侵的能力,其中乳酸菌促進AhR依賴的IL-22轉錄,維持微生物多樣性,抑制白色念珠菌定植,保護腸道黏膜免受炎癥[81]。KYN具有抗菌活性,可直接影響腸道菌群的增殖[35]。在腸道中,色氨酸被微生物降解為吲哚及其衍生物,產吲哚細菌能抑制非產吲哚細菌的生長和存活[89]。研究發現,吲哚和IE等某些吲哚類化合物對革蘭氏陰性腸桿菌有抑菌作用,尤其是沙門氏菌屬和志賀氏菌屬。此外,研究發現,IAA可抑制乳酸菌的生長和生存,特別是對副干酪乳桿菌[90]。還有一個有趣的發現,飼糧中添加0.42%的色氨酸在肉雞中具有類似益生元的作用,對致病性和非致病性菌種進行定量分析發現,非致病性菌(雙歧桿菌屬和腸球菌)數量隨著致病性大腸桿菌、梭狀芽孢桿菌、腸桿菌和大腸桿菌數量的減少顯著增加[91],其背后的確切原因尚不清楚,需要進一步的多學科研究來闡明這個問題。然而,可以推測,產吲哚的細菌可能會抑制不產吲哚的細菌如腸桿菌的生長和存活,特別是沙門氏菌和志賀氏菌屬[89];Nowak等[90]也報道了添加到乳酸菌培養物中的IAA具有同樣的生長抑制作用,但其背后的作用機制還需進一步探討。
4 小 結
色氨酸具有調節生長性能、抗應激和免疫反應的重要功能。越來越多的研究表明,色氨酸及其代謝產物(5-HT、褪黑激素、犬尿氨酸、吲哚及其衍生物等)對腸道微生物組成和功能、宿主免疫系統與中樞神經系統的作用具有深遠的影響。相應地,腸道菌群影響宿主對色氨酸的吸收和代謝,并直接或間接地調節隨后宿主的生理和代謝。在熱應激狀態下,飼糧中補充色氨酸可以增加采食量、調節能量代謝、增強腸道黏膜屏障功能、調節畜禽免疫功能和抗應激能力。熱應激是一個綜合反應,色氨酸是一系列生物活性物質前體,畜禽處于熱應激,機體內色氨酸的3條主要代謝途徑會如何變化,其與機體免疫的交叉點仍有待研究。熱應激導致腸道微生物紊亂,腸道菌群的改變又是如何通過調節色氨酸代謝影響宿主免疫?從細胞到整個機體,熱應激狀態下色氨酸復雜代謝及其調控熱應激作用機理需要進一步的研究,并針對色氨酸代謝物或色氨酸-腸道菌群-免疫相互作用設計可行的可能緩解畜禽熱應激的方法。另外,國內外研究關于熱應激條件下色氨酸在各個物種、品種、日齡階段的添加量存在較大差異,目前亟需確定畜禽不同品種、不同日齡色氨酸最佳需要量,來指導和完善抗應激飼糧的綜合配制。
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
