微生境對耐旱石生毛尖紫萼蘚水分保持與散失特性的影響
2021-02-26 08:40:06邱東張靜吳楠,2陶冶*
植物研究 2021年2期
邱 東 張 靜 吳 楠,2 陶 冶*
(1. 中國科學院新疆生態與地理研究所,干旱區生物地理與生物資源重點實驗室,烏魯木齊 830011;2. 魯東大學資源與環境工程學院,煙臺 264025)
裸巖是森林生態系統的特殊生境,且因巖面大小、朝向、平整度、坡度及有無遮蔽物的不同,裸巖又可衍生出諸多微生境類型,并在溫度、光照和水分條件等方面表現出極大差異性[1]。在山地裸巖生境中,除苔蘚以外的其他陸生高等植物幾乎難以生存和繁衍[2~3]。因此,這些石生苔蘚既是所在生態系統的先鋒物種,同時也是維持生態系統功能不可缺少的組成部分[4~7]。在這些石生苔蘚中,耐旱的種類生存環境更為殘酷,其基質巖石溫度日變幅大,白天可達50~60℃,而夜間又很快降到最低點[3]。而且干燥巖石缺乏土壤成分,無法有效持水,石生蘚類只能利用自己的表面來吸收降雨、雪融水、隱匿水等,以維持生存和繁衍[2]。在長期進化適應過程中,耐旱蘚類在外部形態(如具有葉片毛尖)、細胞結構、生理調節、基因調控等方面均形成了有效的適應機制[7~13],但仍然需要面臨高不確定性的變水環境和水分脅迫。
微環境對耐旱蘚類形態和生理等功能性狀有著非常顯著的影響。如遮陰環境下荒漠齒肋赤蘚(Syntrichia caninervis)的蓋度和生長狀況好于暴露環境下的植株[14],且前者具有更大的葉綠體及更多的類囊體和基粒[15]。而干燥暴露環境下的苔蘚其葉片毛尖與葉長的比例較蔭蔽環境下植株更大,這有利于獲取更多凝結水并延長水分保持時間[12~13]。對生理功能的研究指出,暴露環境下的苔蘚植物比隱蔽環境個體具有更強的脅迫耐受性[16~17]。由此可見,在長期適應進化過程中,同種苔蘚植物的不同功能性狀在不同微環境下產生了不同的適應性變異和權衡[7]。那么,與荒漠極端干旱環境相比,生于干燥裸巖環境的耐旱蘚類對不同微生境具有何種功能性狀權衡,需做進一步研究。
毛尖紫萼蘚(Grimmia pilifera)是我國廣布的苔蘚植物之一[18],屬典型的耐旱石生蘚類,業已成為開展苔蘚植物耐旱機理研究的模式物種之一[11,19~22]。充足的水分是植物生命活動的前提,尤其是生于極端干燥和無規律變水環境(裸巖)中的石生蘚類植物。研究表明,毛尖紫萼蘚必須維持一定的含水量(相對含水量不低于35%)才能保持有效的光化學效率[23]。在水分來源上,苔蘚植物除了通過體內吸收和保持水分外,其體表所具有從納米、微米到厘米級的多尺度水分收集與傳輸系統,可以幫助苔蘚植株固持大量體外水,是苔蘚植物生命代謝活動的重要保障[2,8,13]。因此,能否吸收足量水分(包括內部和外部水)、有效保持水分、延緩水分散失并延長水合時間是耐旱蘚類生存和生長所面臨的首要挑戰。
當前對毛尖紫萼蘚的研究多集中于耐旱相關基因[11,20~22]、脅迫(溫度、光照、重金屬等)生理[19]及脫水——復水過程中光化學效率[23]等方面,而關于微生境對毛尖紫萼蘚水分生理影響的研究尚未見報道。對喀斯特地區耐旱石生蘚的水分生理有一些研究,但關注點在于不同物種之間[2,24]或干旱脅迫程度的比較[8,25~26]。鑒于此,本研究以位于北亞熱帶的安徽省安慶市大龍山國家森林公園為研究區,選擇向陽裸巖及兩種不同植物遮蔽裸巖共3種典型微生境,對比研究不同微生境下毛尖紫萼蘚個體大小、內外含水量及脫水過程中水分保持和散失的差異性,綜合判斷微生境對該物種水分脅迫耐受性的影響及其權衡特征。研究結果將為深入了解毛尖紫萼蘚等耐旱石生蘚的功能性狀權衡及耐旱機制提供科學依據,也能為該物種耐旱基因挖掘、基因調控機制的實驗材料選取提供指導。
1 研究地區與研究方法
1.1 研究區概況
研究區位于安徽省安慶市大龍山國家森林公園(116.994°~117.087°E,30.624°~30.692°N),面積為40.18 km2。該地區屬于北亞熱帶濕潤季風氣候,四季分明,氣候溫和,雨量充沛;年均太陽輻射總量為407~491 kJ·cm-2,年平均相對濕度77%,年平均氣溫14.5~16.6℃,≥10℃的積溫在5 200℃左右;年平均降水量在1 300~1 500 mm,主要集中在4~8 月,占年降水總量的70%以上;常年主導風向為東北風,占全年風向的52%,其次為西南風;霜少雪稀,無霜期250 d 左右,年日照時數2 012 h[27]。大龍山國家森林公園為大別山南麓余脈,地形復雜陡險,多裸巖。山體坡度多在20°~45°,部分地段在45°以上。植被以馬尾松(Pinus massoniana)林和雜闊林為主,土壤以紅壤、黃壤和黃棕壤為主。
1.2 研究方法
1.2.1 苔蘚樣品采集與處理
2018 年1 月初,在大龍山森林公園東部靈山石樹景區開展毛尖紫萼蘚調查采樣。選擇竹林邊緣蔭蔽巖石(記為S-1)、向陽裸巖(記為E)及薜荔(Ficus pumila)灌叢下巖石(記為S-2)3個典型微生境,測量記錄巖石面積、苔蘚斑塊面積、苔蘚密度、坡面的坡向、坡度及有無成土等指標,隨后用小型取樣鏟輔助采集適量苔蘚樣品,裝入封口袋內標記保存。苔蘚樣品帶回實驗室后,用多級篩網過濾雜質,平鋪于報紙上,置于實驗臺自然干燥。
1.2.2 植株飽和含水量及個體大小測定
實驗于2018年1月24日開展。提前對各微生境毛尖紫萼蘚植株進行人工篩選,去除夾雜泥沙,剪除下部無葉部分。選取不同微生境毛尖紫萼蘚植株各25 株(為1 份),每個微生境取6 份(即6 個重復),共18份苔蘚樣品。將苔蘚植株用蒸餾水充分淘洗,以去除夾雜的泥沙。隨后放入盛有蒸餾水的培養皿(高1.5 cm,內徑6 cm)內充分水合2 h,期間翻動幾次,以去除植株葉片間夾雜的微小氣泡和雜質。然后用手術鑷子分別將水合的苔蘚樣品夾出放在對應干燥培養皿內(培養皿質量記為W1),稱重記錄(質量記為W2)。上述操作在2 min內完成,且夾出樣品時不可甩動,以免植株外部水分丟失。將培養皿內的苔蘚樣品倒在多層濾紙上,迅速吸干外部水分,放入另一批干燥培養皿內(培養皿質量記為W3),稱重記錄(質量記為W4)。隨后開始脫水實驗(見1.2.3)。脫水實驗完成后,將各苔蘚樣品連同培養皿放入75℃烘箱內干燥24 h,放至室溫后稱重(質量記為W5)。3 個微生境毛尖紫萼蘚樣品分別采用3 臺精度0.000 1 g 電子天平同時測定。相關指標計算方法為:
含內外水的飽和鮮重:
僅含內部水的飽和鮮重:
單株干重:
包含內外水的飽和含水量:
飽和外含水量:
飽和內含水量:
包含內外水的含水率:
內含水率(即一般講的含水率):
式中,n=25。同時計算外吸水量與內吸水量之比(EW∶IW),以對比不同生境苔蘚內外吸水量的差異。
1.2.3 苔蘚脫水過程中植株含水量的測定
將上述吸干體外水分的苔蘚樣品用于脫水實驗。以稱量飽和內含水量為開始(即第0 min,W4為始重),隨后以不同時間間隔累計稱量28 次,共計305 min(即第0、5…75、85…125、145…185、215…305 min)。每個時間點(用i 表示)苔蘚樣品重量(含培養皿)記為Wi,其中0 點時(即i=1)的重量為W4,烘干后質量為W5,培養皿質量為W3。3 組實驗使用3臺電子天平同時開展,時間為2018年1月24 日10:02~15:07,實驗過程中室內溫度15℃,相對濕度65%。
分別采用基于植株干重的絕對含水量(AIW,%)、基于飽和水重量的相對含水量(RW,%)、基于鮮重的含水量(WC,%)以及蒸散率(ER,%)、累積蒸散率(CER,%)等指標來反映不同生境下毛尖紫萼蘚脫水過程中含水量和水分散失的變化。
式中,i 表示時間序列(1~28),當i=1 時,AIW=SIW。
1.2.4 統計分析方法
對不同微生境毛尖紫萼蘚鮮重、干重、各含水量及不同脫水階段的含水量指標進行One-way ANOVA分析,采用Duncan法進行多重比較。對脫水過程中相對含水量的變化進行最優曲線擬合,并計算達到指定含水量所需的時間。以飽和狀態不同含水量參數、蒸散率及脫水時間等多個指標綜合判斷不同微生境毛尖紫萼蘚耐水脅迫能力的差異。常規統計分析和作圖在Excel 2013 完成,ANOVA和曲線擬合采用SPSS 19.0軟件實現。
2 結果與分析
2.1 不同微生境毛尖紫萼蘚個體大小及內外飽和含水量的差異
毛尖紫萼蘚單株最大飽和鮮重(FW-A)以向陽裸巖生境最高(55.5 mg),竹林遮蔽生境最低(47.2 mg),薜荔灌叢遮蔽生境居中(52.3 mg)(見圖1A)。僅含內吸水量的單株飽和鮮重(FW-B)也以向陽裸巖生境最高(10.8 mg),薜荔灌叢遮蔽生境次之(9.3 mg),竹林遮蔽生境最低(7.7 mg)(見圖1B)。樣品烘干以后,向陽裸巖和薜荔灌叢遮蔽生境單株干重(DW)最高且無顯著差異(分別為3.1和3.0 mg),而竹林遮蔽生境最低(2.2 mg)(見圖1C)。
毛尖紫萼蘚總絕對飽和含水量(SW)以竹林遮蔽生境最高,達到2 025.9%,即內外總吸水量是自身干重的20.26倍(見圖2A);向陽裸巖(1 697.5%)和薜荔灌叢遮蔽生境(1 657.2%)顯著低于竹林遮蔽生境,且二者無顯著差異。絕對外含水量(SEW,分別為1 779.0%、1 448.9%和1 444.0%)在3 個生境間的變化趨勢與SW 相同(見圖2B)。絕對內含水量(SIW)在3 個生境間的差異性不同于前兩個指標,表現為竹林遮蔽(246.9%)和向陽裸巖生境(248.6%)無顯著差異,但顯著高于薜荔灌叢遮蔽生境(213.2%)(見圖2C)。對3 種生境外吸水量和內吸水量進行對比發現,盡管竹林遮蔽和薜荔灌叢遮蔽生境的內、外吸水量和吸水總量呈現出顯著差異,但在外部吸水能力上(相對于內吸水量)二者沒有顯著差異(EW:IW 分別為7.2 和6.8),且顯著高于向陽裸巖生境(EW:IW 為5.8)(見圖2D)。
從基于鮮重的含水量來看,毛尖紫萼蘚總飽和含水量(TWC,包含內外水)的變化趨勢同SW 和SEW,以竹林遮蔽生境最高(見圖2E)。而含水量(WC,僅內吸水量)的差異性則與SIW 相同,即以竹林遮蔽(71.1%)和向陽裸巖生境(71.3%)最高,薜荔灌叢遮蔽生境最低(68.0%)(見圖2F)。
2.2 不同微生境毛尖紫萼蘚脫水過程中植株含水量變化
分別采用基于干重的絕對含水量(AIW,圖3A)、基于飽和水重量的相對含水量(RW,圖3B)和基于鮮重的含水量(WC,圖3C)共3 個指標來反映不同生境下毛尖紫萼蘚脫水過程中含水量的變化。實際脫水時間為305 min,但185 min 以后各生境苔蘚含水量不再發生變化,因此圖中只保留脫水時間到185 min。無論哪種含水量表示方法,向陽裸巖生境的毛尖紫萼蘚含水量降低的速度最慢,且在第185 min 之前均顯著高于薜荔灌叢遮蔽生境(見圖3)。竹林遮蔽生境與向陽裸巖生境苔蘚的初始含水量相同(見圖3C),但隨后竹林遮蔽生境苔蘚含水量降低速度逐漸加快,至第25 min時與向陽裸巖生境呈現顯著差異,并一直保持到第165 min。用相對含水量表示脫水過程時(見圖3B),3 個微生境均符合“S”型邏輯斯蒂曲線,即含水量呈“緩慢—快速—緩慢”的下降過程。相比之下,竹林遮蔽和薜荔灌叢遮蔽生境苔蘚含水量始終無顯著差異,但自第5 min 開始直至第145 min,二者含水量均顯著低于向陽裸巖生境。
采用基于飽和水的RW 來擬合脫水過程中毛尖紫萼蘚含水量的變化趨勢。結果表明,3個微生境毛尖紫萼蘚脫水最優擬合曲線均為二次方程,決定系數(R2)均在0.998 以上(見表2)。給定了5個RW 值,計算了各擬合方程到達指定含水量所需的時間。可以發現,竹林遮蔽和薜荔灌叢遮蔽生境毛尖紫萼蘚在到達任何指定含水量所需時間均較一致,且所有時間均短于向陽裸巖生境。也即向陽裸巖生境毛尖紫萼蘚比竹林遮蔽和薜荔灌叢遮蔽生境植株到達指定含水量所需時間更長,失水更慢。不僅如此,隨著脫水進程持續,這種時間差逐漸擴大,由RW=80%時的5 min 時差擴大到RW=20%時的26 min 時差(見表2)。在毛尖紫萼蘚有效光合作用的轉折點RW=35%時[23],向陽裸巖生境苔蘚植株的有效光合作用時間比另兩個生境延長了18 min。
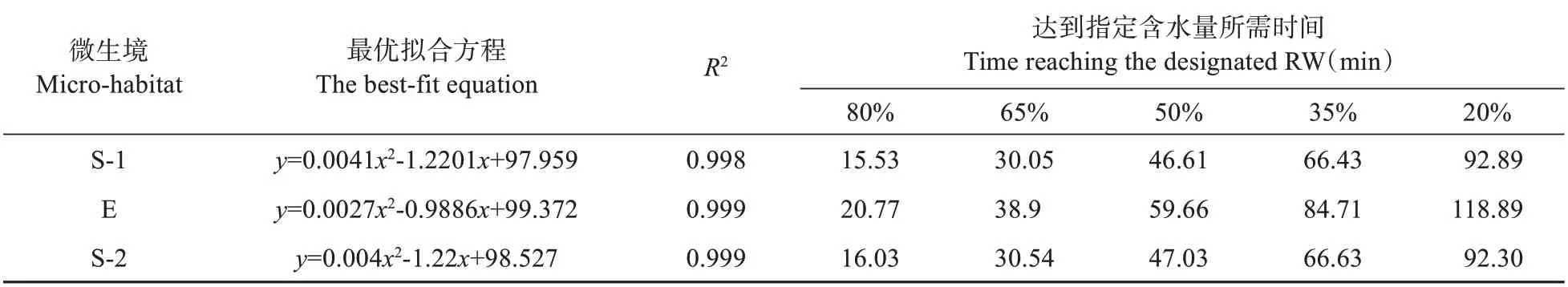
表2 基于相對含水量(RW)的不同微生境毛尖紫萼蘚最優脫水擬合方程及達到指定含水量所需時間Table 2 The best-fit equations for RW of G. pilifera in different micro-habitats during dehydration and the time reaching the designated RW
2.3 不同微生境毛尖紫萼蘚脫水過程中植株水分散失特征
脫水過程中,各微生境毛尖紫萼蘚水分蒸散率(ER)呈兩個階段的減小趨勢(見圖4A):脫水開始至第70 min 為第一個下降階段,隨后第75 min開始上升,至85 min 時蒸散率達到新高(分別為5.9%、5.9%和6.5%),進而進入第二個下降階段。在第一階段,向陽裸巖生境毛尖紫萼蘚植株水分散失率顯著低于竹林遮蔽和薜荔灌叢遮蔽生境(第30 min 除外),即后兩者一直維持較高的失水率。而在第二階段,情況發生反轉,即向陽裸巖生境苔蘚水分散失率高于竹林遮蔽和薜荔灌叢遮蔽生境。盡管如此,由于脫水前期竹林遮蔽和薜荔灌叢遮蔽生境苔蘚維持較高的脫水率,在第一階段蒸散率最低的第70 min 時,其累積蒸散率已達67.0%和66.7%,而向陽裸巖生境的苔蘚累積蒸散率僅為55.3%(見圖4B),相差近12%。因此,無論從蒸散率還是累積蒸散率來看,向陽裸巖生境總比另外兩個生境表現出更強的保水能力。
此外,從蒸散率的變異性來看,整個脫水期間(0~185 min)竹林遮蔽和薜荔灌叢遮蔽生境的毛尖紫萼蘚蒸散率變化曲線近乎一致,且表現出較大波動性(見圖4A),其變異系數(0.46 和0.47)明顯高于向陽裸巖生境(0.30)。這表明,向陽裸巖生境的毛尖紫萼蘚在應對外界水分變化和自身脫水時能保持相對穩定的脫水速度,這也為其進行脫水后期的生理和結構適應性調節贏得更多時間。
3 討論
植物生命活動離不開水。不同類型植物對水分的吸收和保持能力有很大差異。種子植物具有發達的維管系統,表現出高效的水分吸收、運輸和儲存,因而具有較強的體內水分調節能力。但苔蘚植物個體矮小,沒有維管結構,葉片常為單層細胞;其假根基本不具吸水能力,水分吸收以葉片為主,且極易失水[9],因此屬于典型的變水植物(poikilohydric plant)[7]。外界環境對苔蘚植物含水量的影響較大,對于生長于陰濕環境的苔蘚種類而言,其外部水分環境相對穩定,植物體也可以保持較穩定的水合狀態。而對于旱生蘚類而言,除非特殊天氣(如連續陰雨)或人工干預,否則植物體很難長時間保持水合狀態,其體內水分會在很短的時間內蒸發殆盡。因此,相比于其他高等植物,苔蘚植物保水能力差,變水性強,其中以旱生蘚類最為典型。
盡管耐旱蘚類面臨嚴峻的水脅迫且自身控水能力差,但它們進化出了多種延緩水分散失和增加植株含水量的結構性適應[28~29]。紫萼蘚屬植物通常成密集墊狀、叢狀或毯狀的蘚叢,提高了毛細管系統的持水力,并能有效減少水分蒸發;其葉片干燥時強烈卷縮且具有白色毛尖,不僅能反射強光,減少輻射傷害和水分散失[8,23,28],而且“毛尖—葉片—植物體”還是一個高效的多尺度的水分收集系統,對耐旱蘚類的水分收集、運輸和保持都發揮巨大作用[12~13]。耐旱蘚類通常葉細胞小、細胞壁厚,其表面的疣狀突起也有利于減少水分散失[8~9,23]。此外,在變水過程中,耐旱蘚類葉細胞內膜系統的完整性也為其快速恢復生理活性提供了保障[9]。相比之下,具有明顯葉片毛尖結構的毛尖紫萼蘚也具有上述耐旱蘚類相似的形態結構特征,但細胞層面上的微結構特征在不同微生境間有無顯著差異尚不清楚,這部分研究工作還在進行之中。
就目前已測不同微生境毛尖紫萼蘚植株形態進行對比分析可知,竹林遮蔽生境下苔蘚個體最高,但寬度最小,個體重量也最小;然而,該生境苔蘚植株各飽和含水量指標均較高,表明其內外吸水能力均較強。向陽裸巖和薜荔灌叢遮蔽生境毛尖紫萼蘚個體大小接近,但向陽裸巖生境苔蘚外吸水量低于而內吸水量卻高于薜荔灌叢遮蔽生境。從內外吸水量比例來看,竹林遮蔽和薜荔灌叢遮蔽生境EW:IW 值相近,而向陽裸巖生境較低。可見,3個微生境毛尖紫萼蘚的內外吸水能力各不相同,但實際水分保持能力卻以向陽裸巖生境為高。自脫水伊始,向陽裸巖生境苔蘚植株就表現出較強的水分保持能力,其RW 始終高于另兩個生境,且隨著脫水時間延長,這種達到相同含水量的時間差愈來愈明顯,表明向陽裸巖生境中生長的毛尖紫萼蘚具有更強的脫水耐受性。
目前對不同生境下耐旱蘚類形態結構、生理活性差異性了解最多的是荒漠生境中生存的齒肋赤蘚。研究發現,灌叢的存在能夠為蘚類植物提供一個低輻射、高含水量及低蒸發量的相對較優越的生存環境[16~17],有利于蘚類生存。因此,就微生境對耐旱蘚類的利弊來講,蔭蔽生境應當是一種更好的生存選擇[30]。但是,一旦遮蔽環境變為裸露環境,蘚類植物將面臨嚴峻的生存考驗。去除灌叢后(即蔭蔽生境變為向陽暴露生境),齒肋赤蘚植株葉綠素熒光活性持續時間顯著縮短,主要歸因于灌叢移除導致UV-B 輻射增加及“濕島效應”消失所致,而植株自身較低的保水能力(即水分散失過快)也是其中一個重要原因[31]。由此可見,本研究中2個遮蔽生境毛尖紫萼蘚水分散失速度顯著高于向陽裸巖生境;那么,一旦其生境光照增強(如秋冬季節落葉植物葉片脫落導致的遮陰效果減小以及不可預知的人為擾動),毛尖紫萼蘚將面臨一定的環境脅迫,不利于生存。
野外調查發現,與灌叢生境相比裸露地齒肋赤蘚個體較小但密度較高,且其葉長、中下部葉片的中肋寬以及頂部葉片的毛尖長也顯著高于灌叢生境,這些特征證明裸露環境苔蘚具有更強的抗旱性[32]。在生理方面,裸露環境下齒肋赤蘚的光合活性雖然顯著低于灌叢下,但其實際光化學效率卻明顯高于灌叢下[31],而且裸露生境植株在光合活性、抗氧化酶活性、滲透調節物質含量等方面均具有比活灌叢和死灌叢下生長的植株更強的抗脅迫能力[16~17],這顯然是極端環境下耐旱蘚類長期鍛煉和適應進化的結果[28]。沙培實驗表明,取樣于裸露地的齒肋赤蘚,沙培植株表現出較優的生長態勢(如蓋度、密度及株高等均高于灌叢生境),表明裸露地的齒肋赤蘚沙培植株具有高度的可塑性和環境適應性,具有潛在培養價值[32]。相比之下,本研究中向陽裸巖生境毛尖紫萼蘚也應具有裸露地齒肋赤蘚相似的可塑性和環境適應性,故向陽裸巖生境的毛尖紫萼蘚是開展耐旱蘚類植物相關研究的首選材料來源。
水分是苔蘚植物啟動光合作用的關鍵因子。苔蘚植物脫水過程中,葉綠素含量及光合作用強度都會下降,并與含水量呈顯著正相關。研究表明,毛尖紫萼蘚相對含水量(RW)在降到35%之前,其Fv/Fm相對恒定,光合作用較強;而RW 一旦低于35%,其光合效率迅速下降,這說明RW<35%對光合作用已沒有實際意義[23]。本研究中,在相同實驗室環境下向陽裸巖生境下的毛尖紫萼蘚RW 達到35%比竹林遮蔽和薜荔灌叢遮蔽兩個蔭蔽環境下的植株多用了18 min,即向陽裸巖生境的苔蘚植株有了更多的時間進行光合作用,這對于脫水后期苔蘚啟動防御機制大量耗費能量的補充積累至關重要[9]。研究表明,荒漠耐旱蘚類齒肋赤蘚在形態和生理功能上均表現出較強的環境適應性。在脫水6 h 后,其葉綠體發育更加充分,外被雙層膜清晰,類囊體片層豐富,基粒發達,膜系統完整,嗜鋨顆粒增多,而此時葉綠素含量也達到最大值;研究認為齒肋赤蘚在含水量適中時具有最大的光合速率,而非完全濕潤或完全干燥時[9]。這進一步證明,延長水合時間、延緩到達特定含水量(毛尖紫萼蘚為35%)的脫水時間對于耐旱蘚類的生存極為重要[23]。由此可見,本研究中向陽裸巖生境中的毛尖紫萼蘚比遮蔽生境中的類群具有更強的生存能力。
此外,即使RW<35%后毛尖紫萼蘚光合作用微乎其微,但脫水速度對維持正常細胞結構和生理代謝極為重要[33]。在正常慢速脫水的情況下,苔蘚葉片細胞可以通過調節滲透調節物質含量、轉換細胞質為玻璃態、進行細胞壁折疊、細胞器表面積最小化、合成耐旱相關蛋白等方式應對脫水脅迫,并以使植物體保持穩定的質膜和天然蛋白二級結構;而快速脫水的植物細胞內的磷脂易丟失,自由脂肪酸增多,蛋白易變性與聚合,氫鍵聚合減少,而且滲漏的原生質膜也會變得不可修復[28,33]。因此,快速脫水對植物細胞的破壞性顯然大于慢速脫水。本研究中,相比于竹林遮蔽和薜荔灌叢遮蔽生境,向陽裸巖生境毛尖紫萼蘚脫水速度明顯較慢且蒸發速率較為穩定,這為植株啟動脫水應答/抵御機制贏得了寶貴的時間,也為再水化夯實了細胞結構完整性的基石[28~29]。
本研究從苔蘚植物個體水平的水分吸收、保持與散失的角度研究了不同生境毛尖紫萼蘚的耐旱性。不同微生境毛尖紫萼蘚個體大小、飽和含水量及脫水速度各有差異,其中向陽裸巖生境毛尖紫萼蘚比遮蔽生境具有更強的脫水耐受性,而后者可以通過增加外部吸水量來彌補失水過快的缺陷,這體現了不同微生境毛尖紫萼蘚對水分吸收、保持和散失的權衡策略。但這一發現是否為耐旱蘚類共有的特征尚不能定論,還需要開展大量的研究加以驗證。相比之下,向陽裸巖生境毛尖紫萼蘚是研究蘚類植物耐旱機制的首選材料。有必要繼續深入開展不同微生境毛尖紫萼蘚葉、莖微結構、細胞亞顯微結構、光合生理等生理生化特征等方面的研究,從多功能性狀角度綜合分析微生境對毛尖紫萼蘚性狀權衡的影響,為揭示耐旱石生蘚的進化和適應策略提供科學依據。
