板栗與錐栗雜交F1代葉片表型變異及雜種優勢研究
2021-02-26 08:40:34章平生江錫兵龔榜初賴俊聲吳聰連
植物研究 2021年2期
章平生 江錫兵 龔榜初* 徐 陽 賴俊聲 吳聰連
(1. 中國林業科學研究院亞熱帶林業研究所,杭州 311400;2. 南京林業大學,南京 210037;3. 慶元縣林業局,慶元 323800)
殼斗科(Fagaceae)栗屬(Castanea Miller)植物板栗(Castanea mollissima Blume.)是中國三大特有栗屬樹種之一,素有“鐵桿莊稼”和“木本糧食”之稱[1~2],在我國已有兩三千年的人工栽培歷史,北起遼寧鳳城,南至海南省,全國26 個省(市、自治區)都有其分布[3~4]。長期的人工、自然條件的影響,板栗已形成多種生態類型和地方品種,據不完全統計,我國板栗已形成6 個地方品種群,地方品種更是多達350個(尚不包括有價值的遺傳材料和東北的丹東栗)[5~6],種質資源極為豐富,然而,在實際的生產與應用中,除北方燕山一帶的板栗堅果品質較為優良外,廣大南方地區特別是長江流域和東南地區板栗果實肉質硬、偏粳性,優質且適于加工的板栗優良品種(糯性強、含糖量高)匱乏。錐栗(C.henryi(Skam)Rehd.et Wils.)是中國南方特色栗屬樹種,主要分布于秦嶺、淮河以南的浙江、福建、湖南等14 個省[7],相比南方板栗其堅果含糖量高、糯性好且口感更加清甜脆嫩,非常適于糖炒及生產加工,但目前錐栗野生資源尚未得到充分發掘和利用,大果、高產的錐栗品種資源稀少,且適應性不及板栗[8]。因此,基于板栗、錐栗各自特性,采用雜交育種技術,培育出優質兼大果、高產且適宜加工的栗屬雜交新品種,對于提升中國南方栗良種化水平、推動栗產業發展具有重要的現實意義。
雜交育種作為種質創新和新品種培育最為有效的方法,已在經濟林果樹育種中得到廣泛的應用[9~11]。雜交育種過程中雜交后代能否產生雜種優勢是關乎育種成功的關鍵,同時,對雜交子代表型性狀遺傳變異規律研究更是進行新品種培育的前提及基礎。因而,自20 世紀60 年代國內研究者們就開始對栗屬植物雜交育種展開研究,并相繼取得了一定的研究成果[12~14],同時,針對栗屬雜交后代表型性狀遺傳變異也進行了一些探索,但研究主要集中于產量、營養品質等果實性狀[2,15-16],而關于栗屬雜交子代葉片性狀遺傳變異規律及雜種優勢的研究報道較少。
葉片是果樹進行光合碳同化的主要器官,也是構成果樹生長發育的基本結構和功能單位,能為果樹的生長發育及開花結實提供充足的養分保障。葉片性狀作為果樹幼林期主要的農藝性狀,其生長發育狀況的優劣能夠在很大程度上影響到果樹的果實經濟產量與品質[17~18]。此外,相關研究表明,果樹的葉片性狀與果樹產量、品質等重要經濟性狀之間也存在較大的關聯[19~21],因而,對果樹雜交子代幼林期葉片性狀遺傳變異規律和雜種優勢進行研究,可起到早期鑒定和預先選擇的作用[18,22~24]。因此,本研究以板栗和錐栗種間、種內雜交子代235株幼樹及其親本為研究對象,利用統計學方法對其14個葉表型及光合生理指標的遺傳變異規律及雜種優勢進行分析,并結合主成分分析法對雜交子代進行初步篩選,旨在了解栗屬雜交子代葉片性狀變異特征,探索雜種優勢表現,以期為栗類雜交子代預先選擇提供一定的理論依據。
1 材料與方法
1.1 試驗地概況和試驗材料
試驗地位于浙江省慶元縣黃田鎮關門岙錐栗園區(118.93°E,27.77°N),海拔300 m,土壤類型為紅黃壤,肥力中等。氣候屬東亞副熱帶季風區,溫暖濕潤,四季分明,年平均氣溫20.02℃,年均降雨量1 417.3 mm,年均無霜期345 d。
供試材料為板栗、錐栗種內和種間雜交F1代群體及其親本,共9個雜交組合(見表1)。2011年通過人工控制授粉獲得雜交種實,2012 年播種育苗,2013年春季按照完全隨機區組設計建立雜交子代測定林,10株一小區,3次重復;同時種植所有組合的父本和母本1年生嫁接苗若干株,株行距4 m×4 m,每年進行精細撫育管理,目前子代單株生長發育狀況良好。9個雜交組合共有7個親本,包括1個板栗品種和6個錐栗品系,其中“魁栗”為浙江省板栗優良品種:YLZ 1 號、YLZ 2號、YLZ 24號、YLZ 26號為浙江省審認定錐栗良種,YLZ 14號、YLZ 15號為錐栗優株無性系,均是從浙南、閩北錐栗實生群體中選優而來。
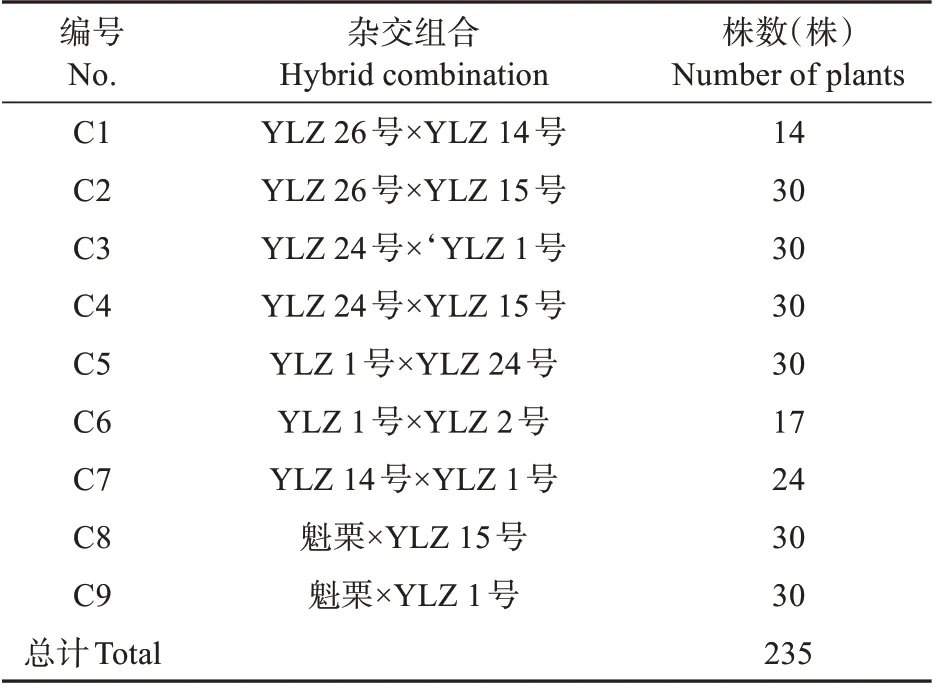
表1 雜交組合編號Table 1 No.of hybrid combination
1.2 試驗方法
分別于2018、2019 年7 月份,即栗樹葉片生長發育完全、干物質含量較為穩定時,對所選取的235 個雜交子代及其親本的葉表型及相關光合色素含量進行測定。具體方法如下:
葉表型性狀測定:從每個單株樹體中上部選擇成熟且無病蟲害的葉片,分東、南、西、北4 個方向采集30~50 枚,帶回實驗室進行葉表型性狀測定。其中葉片長度(LL/cm)、葉片寬度(LW/cm)、葉形指數(LSI)、葉面積(LA/cm2)等采用CI-202 便攜式葉面積儀(精度為0.01 cm 或0.01 cm2)測量,葉片厚度(LT/mm)、葉柄長度(PL/mm)采用游標卡尺(精度為0.01mm)測量,葉片鮮重(LFW/g)采用百分之一天平稱量,測量完成后對葉片進行105℃殺青處理后置于65℃的烘箱中烘干至恒重后稱量其干重,并根據葉片鮮重(LFW/g)、葉片干重(LDW/g)和 葉 面 積(LA/cm2)分 別 計 算 葉 片 含 水 率(LMC/%)和比葉重(SLW/g·cm-2)。
光合色素含量測定:從所采集235個雜交子代及其親本的葉片中,分別隨機抽取5 枚,用于光合色素含量測定,利用簡易提取法(95%乙醇避光浸提48 h)提取其葉綠素[25],BMG LABTECH 多功能酶標儀在波長470、663 和645 nm 下測定其吸光度,計算類胡蘿卜素(Cx.c)、葉綠素a(Chla)、葉綠素b(Chlb)及總葉綠素含量(Chla+b),公式如下:
式中:A470、A663、A645分別表示470、663、645 nm波長下的吸光值,V表示提取液體積(25 mL),M表示樣品質量(0.2 g)。
1.3 數據處理
對各性狀采用巢式設計方差分析,線性模型為:
式中:Yijk為第i 個組合第j 個單株第k 個觀測值,μ為總均值,Si為區組效應(固定),T(i)j為區組內家系效應(隨機);?(ij)k為試驗誤差。
各性狀遺傳參數分別以變異系數(CV)、遺傳傳遞力(Ta,%)、中親優勢(Hm)、中親優勢率(RHm,%)、超親優勢(Hb)、超親優勢率(RHb,%)、表型分化系數(VST)表示[18,24,26],計算公式如下:
式中:S 表示標準差,Fm表示雜交F1代某一性狀的平均值,MPV(中親值)表示親本性狀的平均值,BPV(高親值)表示雙親中較大的親本值。
采用Excel 2003和SAS 9.4軟件對數據進行統計與方差等分析,Duncan’s多重比較。
2 結果與分析
2.1 雜交F1代葉片性狀遺傳變異分析
栗屬雜交F1代葉片性狀遺傳變異見表2,14個葉表型及光合生理性狀在組合間和組合內的差異均達到極顯著或顯著水平,表明雜交子代葉片在組合間和組合內兩個層次上均存在廣泛的變異。各組合中C6的葉綠素a、葉綠素b、葉綠素總量及類胡蘿卜素含量等8個性狀均最小,表明C6的子代葉片發育狀況較差;而組合C8和C9的葉綠素a、葉綠素b、葉綠素總量及類胡蘿卜素含量均最高,且其葉長、葉寬及葉面積等6 個葉表型性狀也均較大,表明C8和C9的子代葉片發育狀況較好。
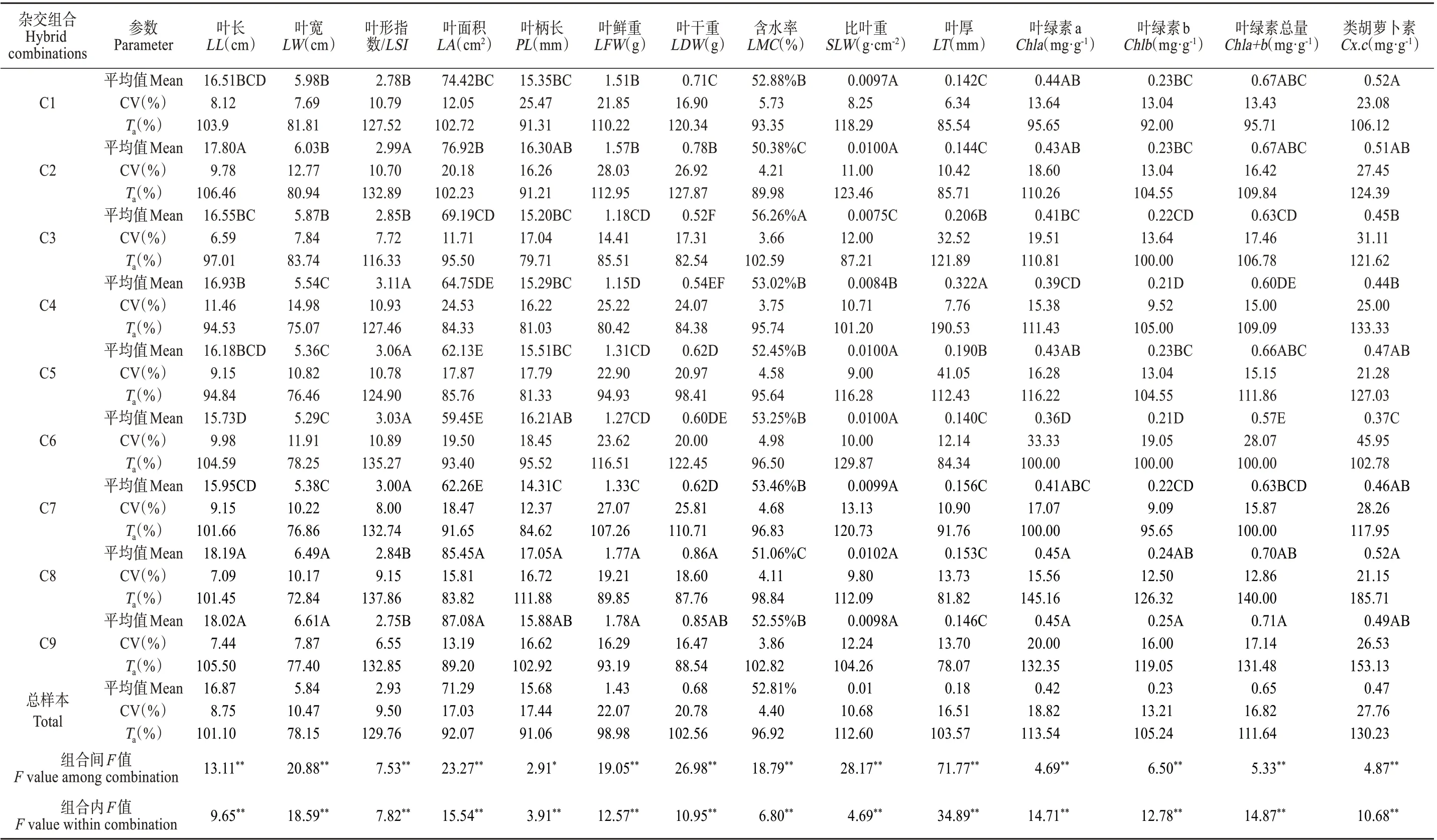
表2 栗屬雜交F1代葉表型及光合生理性狀遺傳變異分析Table 2 Genetic variation analysis of leaf phenotype and Photosynthetic physiological characters in cross F1 generation of Castanea
變異系數是性狀遺傳穩定性的具體體現,變異系數越大,性狀離散程度越大,遺傳背景越豐富,越具有選擇優株的潛力[27]。由表2 可知,F1代葉表型及光合生理14個性狀的平均變異系數范圍為4.40%~27.76%,各性狀中以類胡蘿卜素的變異系數最高,均在21%以上,較其他性狀具有較高的遺傳多樣性;其次是葉面積、葉柄長、葉鮮重、葉干重、葉綠素a 及葉綠素總量,變異系數均在11%以上,變異程度略低于類胡蘿卜素;而含水率的變異程度最低,各組合的變異系數均在6%以下,遺傳多樣性程度最低。不同雜交組合間性狀的變異系數亦不相同,其中,10 個葉表型性狀中葉長、葉寬和葉面積的變異系數均以組合C4 最高,分別為11.46%、14.98%和24.53%;葉鮮重和葉干重均以組合C2 最高,分別為28.03%和26.92%;而4 個光合生理性狀的變異系數均以組合C6 最高,表明組合C2、C4和C6具有較大的選擇潛力。
栗屬雜交F1代葉表型及光合生理14個性狀遺傳傳遞力分析表明:葉形指數的遺傳傳遞力最高,在9 個雜交組合中均大于100%,表現出較高的遺傳穩定性。而葉寬的遺傳傳遞力最低,各組合中均小于85%。不同雜交組合間的遺傳傳遞力存在差異,其中,葉柄長、葉形指數、葉綠素a、葉綠素b、葉綠素總量和類胡蘿卜素含量6 個性狀的遺傳傳遞力均以組合C8 最大,而葉長和葉干重、比葉重和葉鮮重的遺傳傳遞力均分別以組合C2、C6最大。
2.2 雜交F1代葉片性狀群體間表型分化
按巢式設計方差分析將栗屬雜交F1代葉表型和光合生理14 個性狀的變異分為組合間的變異、組合內的變異、機誤三部分,各部分的方差分量百分比反映了F1代葉片性狀的變異來源。由表3 可知,14 個性狀組合間和組合內的方差分量百分比分別在4.045%~69.051%和28.432%~75.546%,表明F1代葉片性狀群體遺傳組成存在較大差異。各性狀表型分化系數VST變動范圍為7.89%~70.83%,其中,葉厚的表型分化系數VST最大,為70.83%,表明其在組合間變異明顯大于組合內的變異;比葉重次之,為53.52%,而其他性狀的表型分化系數VST均小于50%。進一步分析發現,各性狀表型分化系數VST平均值為32.32%,組合內變異平均值為67.68%,表明雜交F1代葉片性狀組合內變異遠高于組合間變異,組合內變異是葉片表型變異的主要來源。
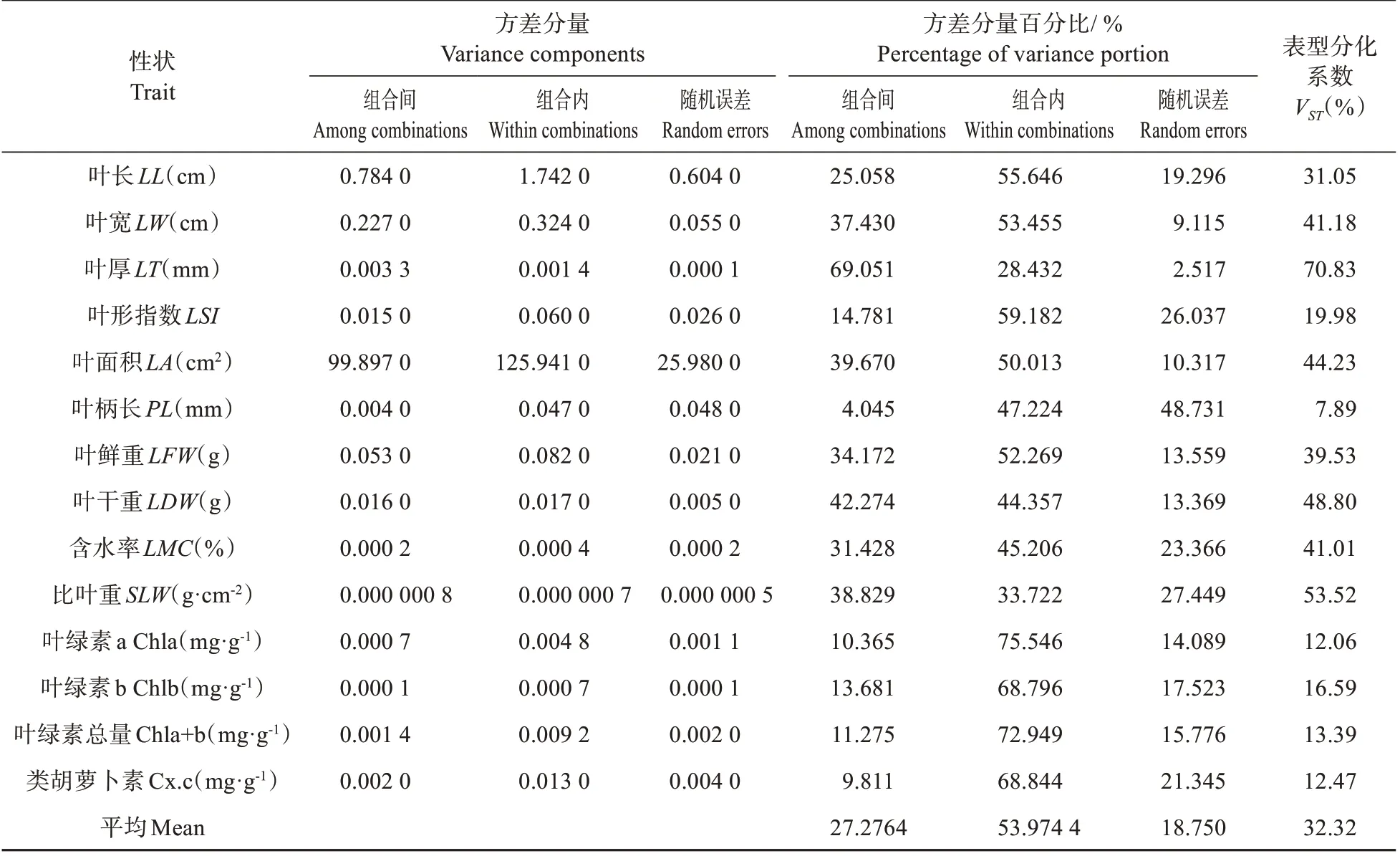
表3 栗屬雜交F1代葉片性狀方差分量及表型分化系數Table 3 Variance portions and differentiation coefficients of leaf characters in cross F1 generation of Castanea
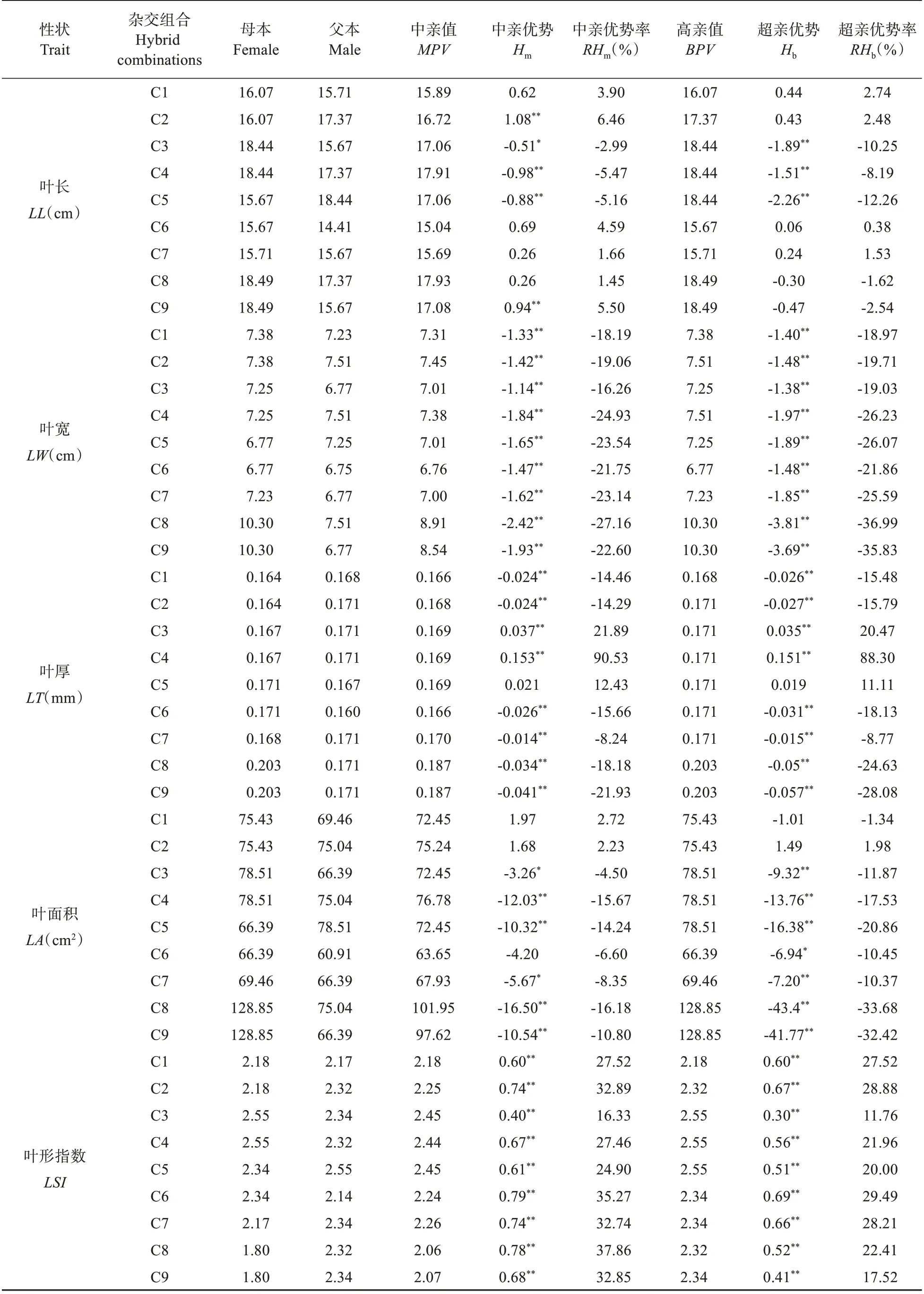
表4 栗屬雜交F1代葉表型及光合生理性狀雜種優勢分析Table 4 Heterosis analysis of leaf phenotype and photosynthetic physiological characters in cross F1 generation of Castanea
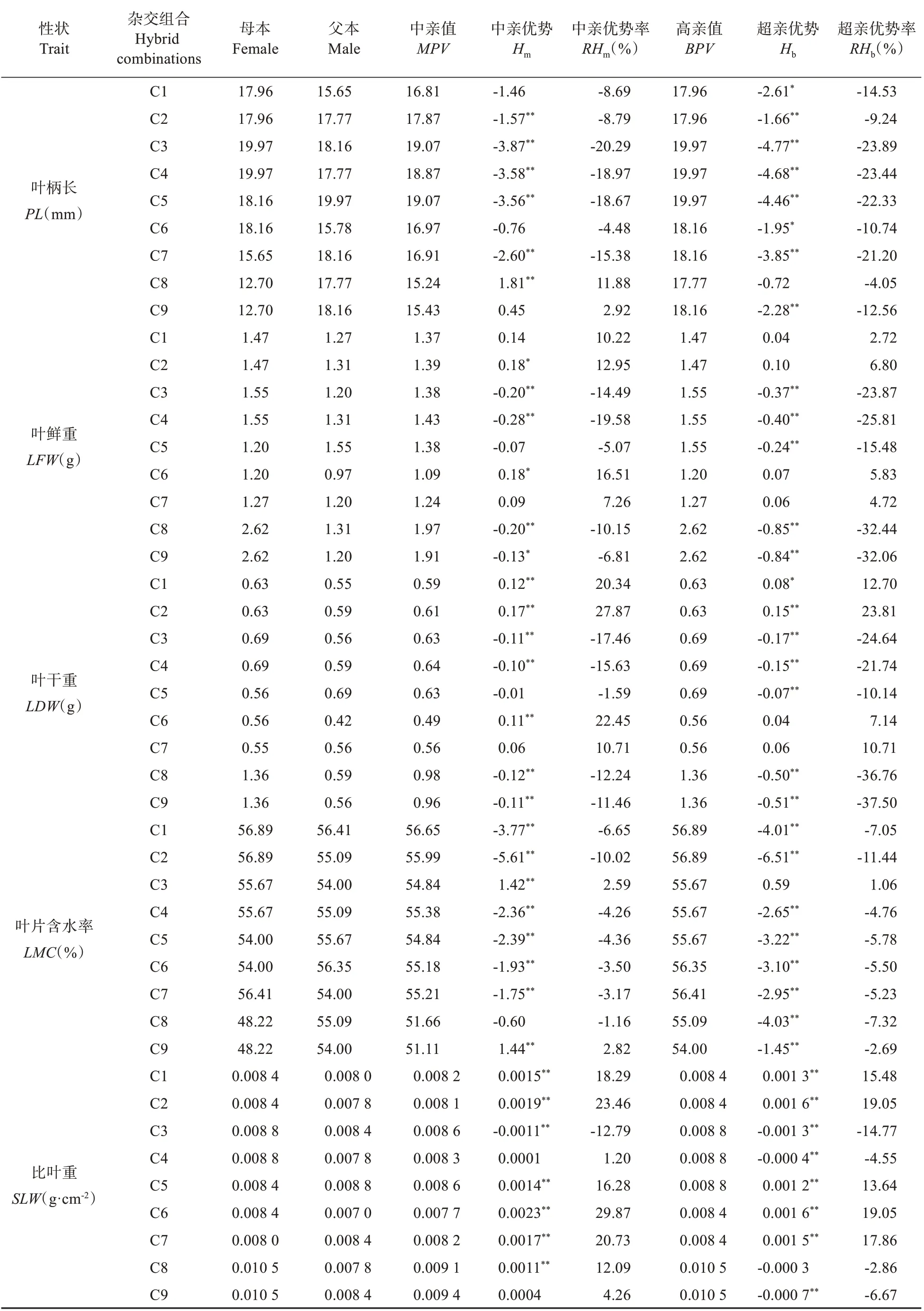
續表4 Continued table 4
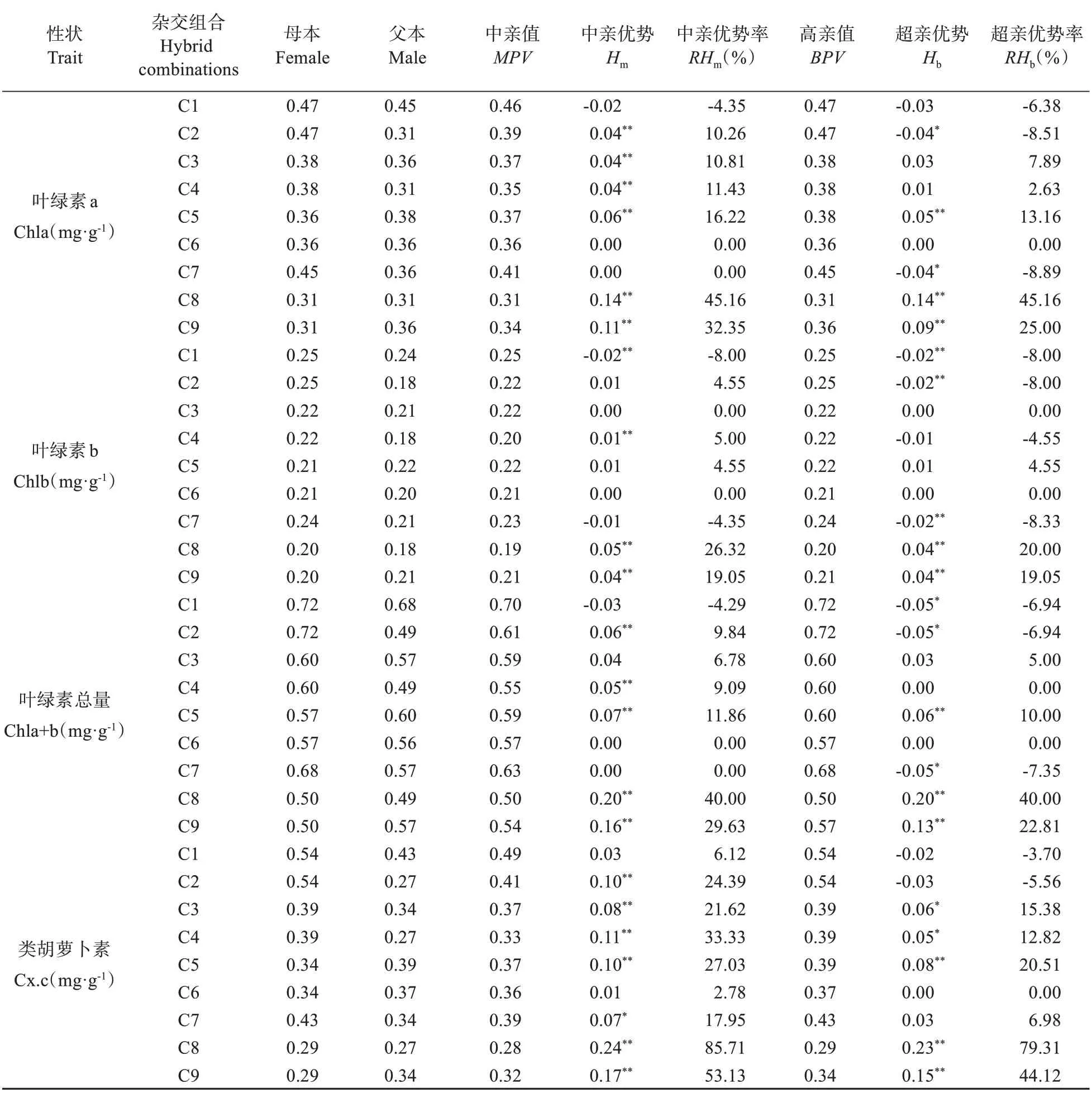
續表4 Continued table 4
2.3 雜交F1代葉片性狀雜種優勢分析
由表4可知,栗屬不同雜交組合F1代葉表型及光合生理14 個性狀的中親優勢率為-27.16%~90.53%。t檢驗表明,除葉長、葉綠素b和葉綠素總量外,其余11 個性狀在絕大多數組合中的中親優勢值均達到極顯著(P<0.01)或顯著(P<0.05)的水平。各性狀中,葉形指數和類胡蘿卜素含量均表現為正向中親優勢,而葉寬卻表現為負向中親優勢,其余性狀均表現出正負2 個方向的中親優勢。各組合中C3 和C4 的葉厚,C2 和C6 的葉干重、比葉重,以及C8 和C9 的葉綠素a 含量、葉綠素b 含量、葉綠素總量及類胡蘿卜素含量的中親優勢率均在19%以上,中親優勢明顯。結合表2 分析,14個性狀中除葉形指數外其他13個性狀超親分離現象均不明顯,且不同組合間性狀的超親優勢存在明顯差異;其中,組合C2 的葉干重、C3、C4 的葉厚和C8、C9 的葉綠素a 含量、葉綠素b 含量、葉綠素總量及類胡蘿卜素含量的均值均明顯高于雙親,超親優勢率均在19%以上,且其相對應的超親優勢值均也達到極顯著水平(P<0.01),表現出明顯的超親優勢;而各組合的葉寬、葉柄長的均值均明顯低于雙親,其超親優勢率為-36.99%~-4.05%,表現為明顯的負向超親優勢。
2.4 栗屬雜交F1代葉片性狀相關性分析
栗屬雜交F1代葉表型和光合生理14個性狀之間的相關性分析見表5,91 對相關性中分別有46對和13對相關性達到極顯著(P<0.01)或顯著水平(P<0.05)。其中,4 個光合生理性狀間的相關性系數均在0.85 以上,相關性程度極高,達極顯著水平;而10 個葉表型性狀中,除含水率外,絕大多數性狀相互間的相關性達到極顯著或顯著水平。10個葉表型性狀與4 個葉片光合生理性狀的相關性分析顯示,葉長、葉面積和葉干重與光合生理4 個性狀間的相關性均達極顯著或顯著水平;葉寬和葉鮮重僅與葉綠素a、葉綠素b、葉綠素總量間存在極顯著或顯著的相關性;而葉形指數、葉柄長、比葉重及葉厚與光合生理4 個性狀間不存在顯著的相關性;由此可見,栗屬雜交F1代葉長、葉寬等5個葉表型性狀及葉綠素a 等4 個光合生理指標相互間影響較大。
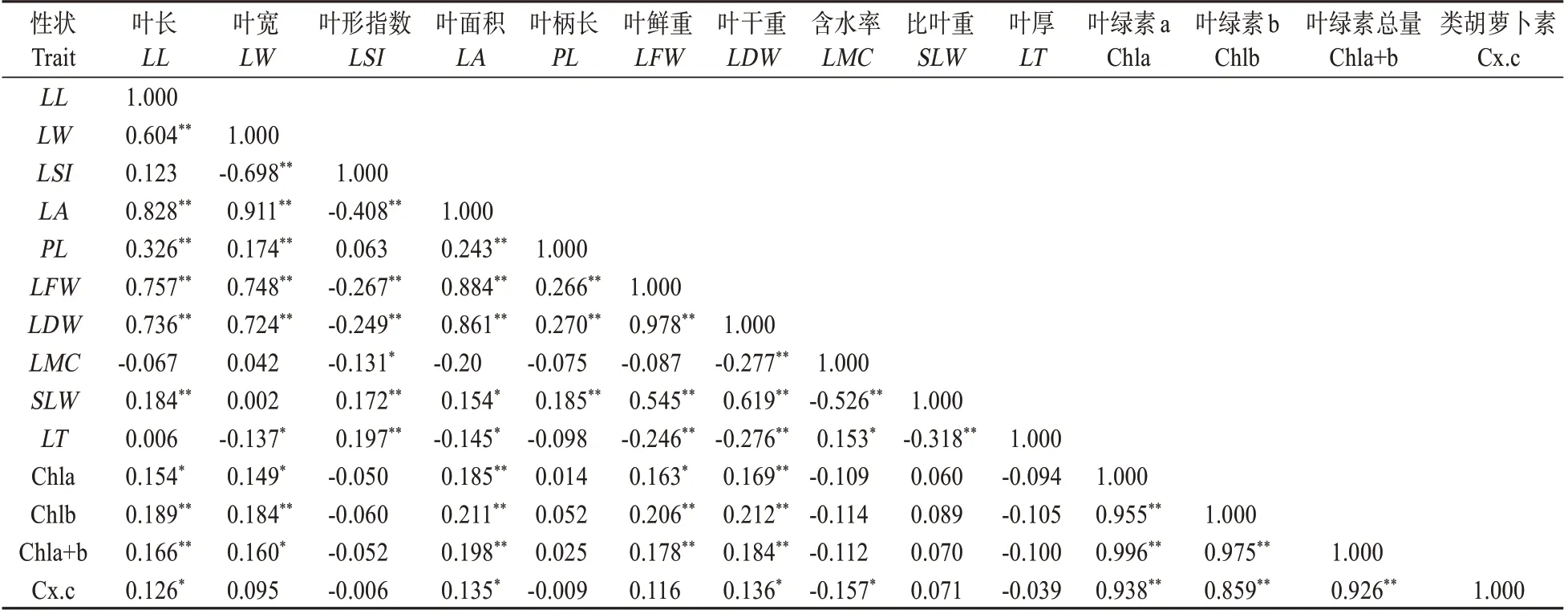
表5 栗屬雜交F1代葉表型及光合生理指標的Pearson相關性分析Table 5 Leaf phenotypic traits and photosynthetic physiological indices in F1 generation of Castanea by Pearson correlation analysis
2.5 栗屬雜交F1代葉片性狀主成分分析
利用F1代葉表型及光合生理14 個性狀對9 個組合235 個單株進行主成分分析(見表6),前4 個主成分的特征值均大于1,累計貢獻率達83.88%,表明這4 個主成分可以反映出235 個單株14 個性狀83.88%的信息。其中,對第一主成分貢獻率較大的性狀是葉長、葉寬、葉面積、葉鮮重及葉干重,即表征葉片大小和重量的綜合因子,累計貢獻率為37.06%,占4 個主成分貢獻率的44.18%。對第二主成分貢獻率較大的性狀為葉綠素a、葉綠素b、葉綠素總量及類胡蘿卜素,即表征光合生理的綜合因子。此外,第三主成分主要反映葉片含水率及比葉重的信息,第四主成分主要反映葉片形態及葉片厚度的信息。
利用主成分分析法對235 個雜交子代進行綜合評價,即通過各主成分得分值與相應特征值百分率的積構建優株綜合評價指數模型,進而利用各性狀標準化數據計算235 個雜交子代綜合主成分值[28]。通過比較分析最終篩選出排名前40的優良雜交子代,其性狀綜合主成分值均在1以上。由表7 可知,綜合評價值前40 名優良雜交子代中,組合C8子代的入選數最高,占入選總數的32.5%,C9次之,占入選總數的27.5%,而組合C3子代的入選數為0,表明組合C8 和C9 子代葉片發育狀況較為優良,可為進一步篩選優良雜交子代的提供主要素材。進一步分析發現,組合C8和C9均為栗屬種間雜交組合,母本均為板栗(魁栗),父本均為錐栗(不同品系錐栗),且其產生F1代的葉長、葉寬、葉面積、葉鮮重、葉干重5 個葉表型性狀值及葉綠素a等4個光合生理指標值均高于而其他7個錐栗種間雜交組合的子代。由此可見,錐栗種間雜交產生的子代葉片性狀多優于種內雜交。

表6 栗屬雜交F1代各性狀相關矩陣的特征向量Table 6 Eigenvectors of correlation matrix of traits in F1 generation of Castanea
3 討論
經濟林果樹雜交后代早期性狀的預先選擇研究一直是國內外研究的熱點問題[22,29~30]。根據果樹早期性狀對其雜交后代進行有效的篩選,可以提高育種效率,加快育種進程[31]。葉片是果樹極其重要的營養器官,也是光合作用的物質基礎和制造營養物質的“綠色工廠”,葉片性狀表現是果樹農藝栽培性狀和生物學特征的重要反映[18,24],研究雜交子代葉片表型等性狀的遺傳變異規律和雜種優勢是果樹遺傳育種的重要研究內容,可以為雜交實生苗的早期鑒定和選擇提供理論依據。
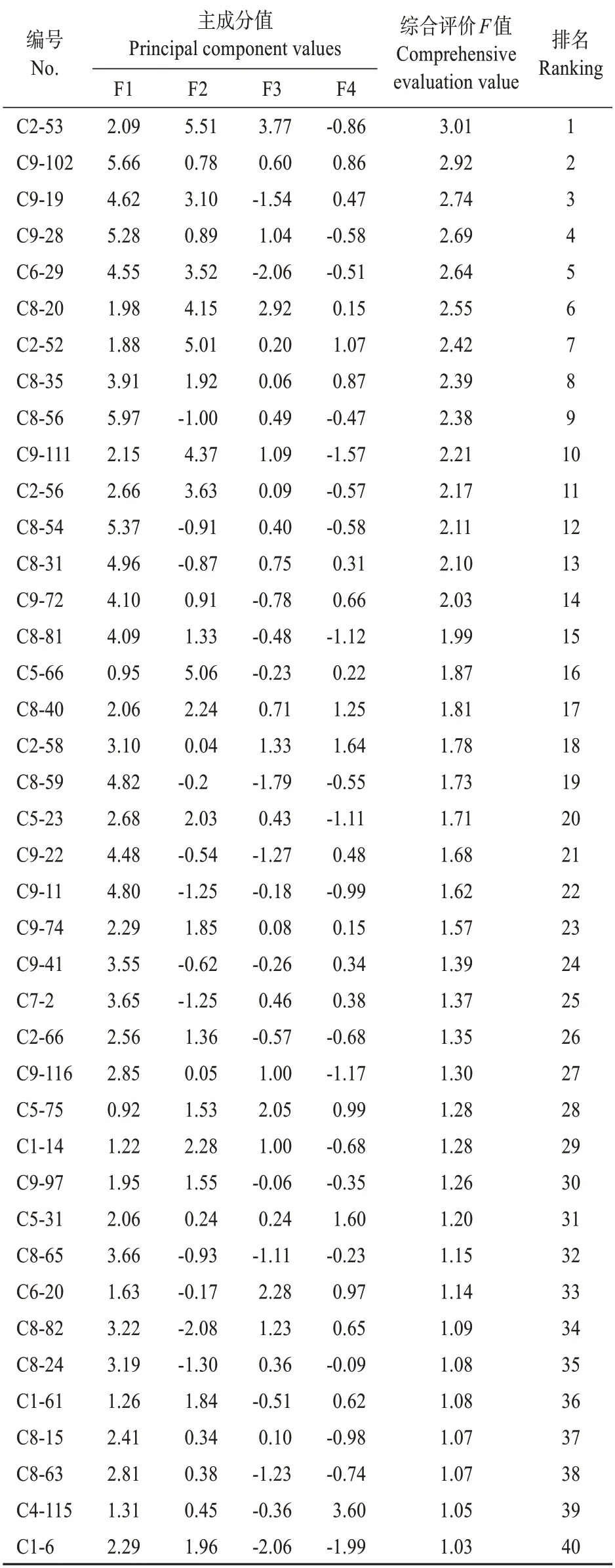
表7 栗屬雜交F1代優樹綜合評價結果排名前40概況Table 7 The top 40 of the comprehensive evaluation results of superiortrees in F1 generation of Castanea
本研究中,栗屬雜交F1代葉表型及光合生理性狀存在較高的遺傳多樣性,14 個性狀在組合間和組合內的差異均達到極顯著或顯著水平,變異系數范圍為4.40%~27.76%,變異程度較大,這與前人關于板栗雜交后代表型性狀的研究結果較為一致[2,8,16]。其中,葉面積、葉柄長、葉鮮重、葉干重、葉綠素a 和葉綠素b 的平均變異系數均在17%以上,表明在F1代中該6個葉片性狀的離散程度較高,存在豐富的表型多樣性及變異水平。同時,F1代14 個性狀組合間平均表型分化系數較低(32.32%),組合內變異遠高于組合間變異,組合內變異是表型變異的主要來源。究其原因可能是由于本次試驗所選擇材料均為板栗、錐栗種內、種間雜交子代,且其生長環境基本相同,因而不存在地理、生殖隔離上的影響;此外,有研究表明栗屬植物葉片性狀是遺傳上較為穩定、不易受環境影響的性狀,主要受自身遺傳因素控制[26]。
雜種優勢是指通過雜交方式產生的F1代,在生長勢、抗逆性、繁殖力、適應性、產量以及品質等性狀上的表現優于雙親的現象[8]。有效利用雜種優勢不但可以提高植物產量,還可改良品質以及增強植物的抗逆性、抗病性,因而其自提出以來,已在糧食作物和大宗果樹中得到了廣泛應用,并取得了巨大的成功。然而,多年來栗屬雜交育種雖已開展了大量的研究,且獲得了一批優良雜交品系,但至今為止關于栗屬植物雜交子代實生植株性狀尤其是幼林期葉表型及光合生理性狀的雜種優勢研究卻鮮有報道。本研究中,栗屬不同雜交組合F1代葉表型及光合生理14個性狀的中親優勢率為-27.16%~90.53%,其中,葉寬在各組合中的中親優勢值和中親優勢率均為負值,表現出極顯著的負向中親優勢,表明葉寬存在具有顯性效應的雜種優勢,此現象在柿(Diospyros kaki)中也有發現[18]。相關研究表明植物葉片的厚度與其抗旱性能間存在一定的聯系,較厚的葉片有利于水分的保存,因此抗旱性越強[32~33]。結合本試驗發現,組合C4 的葉厚中親及超親優勢率分別為90.53%、88.30%,且中親和超親優勢值均達極顯著水平(P<0.01),受基因的加性效應及非加性效應的雙重影響表現出較強的超親優勢[34],因而該組合子代耐旱性較強,具有篩選抗旱品種的潛力。同時,本研究還發現雜交后代葉表型性狀和光合生理指標的平均值大都低于親中值,表現出明顯的性狀衰退現象,此結果與趙爽等[28]對核桃(Juglans regia L.)雜交后代表型性狀的研究結果相似。F1代的雜種優勢源于基因的異質性[18,35],栗屬為雜合基因型,其雜交F1代必然存在廣泛的分離和雜種優勢衰退的現象,本研究也證明了這一結論。雜種優勢衰退的現象可能會導致雜種優勢的利用受到限制,但并不妨礙從F1代中選擇葉面積、葉片質量、比葉重、光和色素含量等顯著高于雙親的、具有速生特點的優良單株,從而培育成新品種。因此,在栗屬雜交育種中,應對上述性狀在F1代中的衰退表現給予充分估計,以達到選育理想雜交品種的目的。
栗屬植物生物學性狀較多,且其相互之間的關系錯綜復雜。Pearson相關性分析能夠揭示各性狀間的相互關系,而主成分分析法可以將許多性狀指標整合壓縮成少量的反映較多信息的綜合性評價指標[36],將二者結合使用,往往能夠更好地把握各性狀的綜合表現。本研究表明栗屬雜交F1代葉面積、葉鮮重及葉干重均與大多數葉片性狀間存在極顯著或顯著相關關系,且主成分分析也表明該3 個性狀屬于反映235 個單株的37.06%信息的第一主成分,結合性狀遺傳變異分析,筆者認為在早期初步篩選優良雜交子代時,可將葉面積、葉鮮重、葉干重作為預先選擇的指標,如可選擇葉面積大、物質含量高的單株作為優良子代。此外,本研究中也利用主成分分析法對栗屬235 個雜交子代的14個葉表型及光合生理性狀進行綜合分析評價,通過主成分分析提取了能夠反映原性狀83.88%的信息的4 個主成分,并用其構建了栗屬雜交子代優株葉片性狀綜合評價指數,綜合分析表明種間雜交組合C8 和C9 多數子代具有葉面積大、干物質含量及光合有關色素含量高等特性,可為進一步篩選優良雜交子代提供主要素材。
本研究從形態學及生理水平上,對板栗、錐栗種間、種內9 個雜交組合235 個雜交子代的葉表型及光合生理性狀的遺傳變異規律及雜種優勢進行了分析,為栗屬植物雜交F1代預先選擇及下一步改良利用提供了一定的理論依據。然而,栗屬為多年生木本植物,生長周期長,導致其雜交育種必將是個漫長的過程,通過對雜交子代幼林期葉片相關性狀遺傳變異及雜種優勢規律的研究只可用于指導雜交子代進行初步篩選,后期還需對其果實性狀(產量、品質等)進行反復測定,并結合分子生物學、基因組學等技術手段進行鑒定和驗證,篩選出真正可直接用于栗屬植物早期選擇的指標,以此指導栗屬植物雜交子代的早期選擇。
