白細胞介素34對大鼠根尖牙乳頭干細胞增殖的影響
2021-01-29 08:46:44王旋宇朱永娜武慶華姜麗娜朱晨晨張曉東
蚌埠醫學院學報 2020年12期
王旋宇,朱永娜,薛 魁,武慶華,姜麗娜,朱晨晨,張曉東
年輕恒牙在發育過程中,常由于各種有害刺激、解剖變異等導致牙髓暴露,牙根發育受限[1]。相對于牙髓治療,以替代或修復病損區細胞為主的干細胞療法成為一種理想的治療方案[2]。根尖牙乳頭干細胞(stem cells from the apical papilla,SCAP)是生長在恒牙牙乳頭內的新型干細胞群體,該細胞具有集落形成、自我更新、多分化及抗感染的優越性,是牙根牙本質再生的種子細胞[3]。當牙髓炎及根尖炎發生時,炎性環境及炎性因子的存在致使牙根發育停止,SCAP介導的牙本質再生功能受限,表明炎性因子可以影響SCAP的特性[4]。
白細胞介素-34(interleukin-34,IL-34)是一種具有復雜生物學特性的炎性因子,它可以刺激破骨細胞的產生,導致炎癥區域的骨吸收[5]。研究[6]發現IL-34在牙周炎和種植體周圍炎中高表達,參與牙周組織破壞過程;另有研究[7]發現IL-34在慢性根尖周病損組織中高表達,參與根尖炎癥反應過程,但炎性環境中的IL-34是否作用于SCAP,尚鮮見研究報道。本研究通過不同濃度的IL-34作用于SCAP,探究IL-34對SCAP增殖能力的影響,從而進一步了解炎性壞境中SCAP特性的改變,為臨床治療年輕恒牙牙髓炎、根尖炎提供理論依據。
1 材料與方法
1.1 試劑與材料 青鏈霉素混合液,高糖培養基DMEM、胎牛血清FBS(美國Gibco);IL-34(上海滬震);CD24單克隆抗體(美國BD);茜素紅染液(美國sigma);MTT(德國BioFROXX);流式細胞儀(美國BD-FACSVerse);細胞培養箱(美國Thermo Scientific 8000);光學顯微鏡、倒置拍照顯微鏡(德國萊卡),低速離心機(上海盧湘儀),酶標檢測儀(美國MDC)。
1.2 方法
1.2.1 SCAP的分離培養 經蚌埠醫學院倫理委員會批準,取2只2~3周齡的SD大鼠,水合氯醛麻醉致死,將根尖牙乳頭組織從根尖分離,PBS清洗后,無菌剪刀剪碎,置于3 mg/mL的膠原酶和4 mg/mL的dispase酶中,37℃的環境下消化30 min,在含有20%FBS的DMEM培養基中生長到70%的融合率,按1∶3的比例用酶消化法傳代。
1.2.2 SCAP的鑒定
1.2.2.1 流式細胞儀鑒定SCAP表面標志物 CD24是SCAP的特征性標志物,在SCAP中優先表達,而在其他間充質干細胞中不表達[8]。故本研究收集第3代SCAP,計數并調整細胞密度為107個/毫升,離心,沉淀后,加入含CD24的100 μL的染色緩沖液,對應孔加入含IgM的100 μL的染色緩沖液,放置4 ℃冰箱避光孵育30 min。洗滌、重懸細胞后,流式細胞術定量檢測。
1.2.2.2 茜素紅染色鑒定SCAP分化潛能 取第3代SCAP,以5×104個/孔接種到12孔板中,以FBS為10%的DMEM培養細胞。待細胞融合率達到70%時,更換成骨誘導液。成骨誘導2周后,茜素紅染色。
1.2.3 四甲基偶氮唑藍(MTT)實驗檢測SCAP增殖能力 調整SCAP密度為2×103個/孔,隨機分為對照組和實驗組6組,分別接種至96孔細胞培養板,每孔液體體積200 μL,待細胞貼壁后饑餓24 h,實驗組加入含IL-34的培養液,濃度分別為25、50、100、200、400、600 ng/mL,每個濃度梯度3個復孔。在細胞培養的第1、3、5、7、9天,酶標儀492 nm波長測出同一時間點吸光度值,實驗重復3次。
1.3 統計學方法 采用方差分析和q檢驗。
2 結果
2.1 SCAP的形態學特征 分離出的大鼠根尖牙乳頭呈條索狀,原發性SCAP在培養5~7 d后呈現典型的細胞集落,顯微鏡下大部分SCAP呈梭形(見圖1)。
2.2 SCAP的鑒定 流式細胞儀結果顯示,第3代SCAP表達CD24為90.7%,呈陽性表達(見圖2)。成骨誘導液誘導2周后,茜素紅染色,鏡下見鈣鹽沉積(見圖3)。
2.3 不同濃度IL-34對SCAP增殖的影響 通過MTT實驗測得各組不同時間細胞吸光度值,結果顯示,第1天600 ng/mL組低于對照組;第3天50 ng/mL組和100 ng/mL組高于對照組,400 ng/mL組和600 ng/mL組低于對照組;第5天和第7天25 ng/mL組、50 ng/mL組和100 ng/mL組高于對照組,400 ng/mL組和600 ng/mL組低于對照組;第9天25 ng/mL組、50 ng/mL組和100 ng/mL組高于對照組,200 ng/mL組、400 ng/mL組和600 ng/mL組低于對照組,差異均有統計學意義(P<0.05)。說明25 ng/mL、50 ng/mL、100 ng/mL 3組濃度的IL-34對SCAP增殖能力具有促進作用,其中50 ng/mL實驗組對SCAP增殖作用最顯著,達到峰值;200 ng/mL、400 ng/mL、600 ng/mL 3組濃度的IL-34抑制SCAP的增殖能力,尤以600 ng/m L實驗組對SCAP的抑制作用最顯著(見表1)。
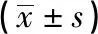
表1 不同濃度IL-34各組吸光度值比較
3 討論
在未發育完全的牙齒中,根尖牙乳頭代表著一個豐富的干細胞池,其中存在的SCAP作為一種外間充質細胞,在牙髓完全喪失的情況下,可以遷移到髓腔,促進牙髓再生[9],且該細胞來源于早期成牙本質細胞,有效促進牙根再生[10]。研究[3,11]發現,與其他干細胞相比,SCAP具有快速增殖、誘導生物遷移和礦化的能力,然而SCAP的特性卻受到炎性環境等因素影響,TOBIAS DUARTE等[12]研究了牙髓感染后根尖牙乳頭的組織學狀況,發現牙髓暴露后,炎性因子的釋放導致根尖牙乳頭組織紊亂,SCAP活力降低。IL-34是2008年發現的新型炎性因子,該因子在牙周膜細胞、牙齦成纖維細胞中均有表達[13-14],并在根尖周炎癥組織中高表達,因此探討IL-34對SCAP增殖能力的影響對臨床具有重要的指導意義。
本研究通過MTT實驗發現低濃度的IL-34對SCAP增殖能力具有顯著的促進作用,其中50 ng/mL的IL-34對SCAP增殖的促進作用最顯著,SéGALINY等[15]研究了IL-34對集落形成樣內皮前體細胞(ECFCs)的影響,發現IL-34可有效促進ECFCs增殖,最佳增殖濃度為50 ng/mL。這與本實驗研究結果一致,提示50 ng/mL的IL-34可能為細胞增殖的最佳濃度。
IL-34在病理生理狀態下具有潛在致病性[16],BATRA等[17]研究發現IL-34在慢性牙周炎和侵襲性牙周炎病人的齦溝液中高表達,HWANG等[18]也發現IL-34在類風濕關節炎病人的滑膜組織和滑液中高表達,并與滑膜炎的嚴重程度有關。本研究發現隨著IL-34濃度的增加,其對SCAP的增殖作用逐漸減弱,并在濃度達到200 ng/mL時抑制SACP的增殖,且濃度越高,抑制作用越強。
IL-34作為根尖炎性環境中的促炎因子之一,與炎性因子腫瘤壞死因子α(TNF-α)關系密切。在類風濕性關節炎病人的滑膜組織中,IL-34作為TNF-α的下游效應因子調節巨噬細胞的增殖和分化,導致炎癥和骨侵蝕[18]。在干燥綜合征中,IL-34與TNF-α的表達呈正相關[19]。LIU等[20]曾研究TNF-α對SCAP增殖能力的影響,發現10 ng/mL的TNF-α對SCAP的增殖作用顯著增強。YANG等[21]也發現10 ng/mL的TNF-α可以提高SCAP的增殖潛能。本研究結果與前述研究相似,發現低濃度的炎性因子IL-34促進SCAP增殖,提示在炎癥的早期階段,IL-34可能是干細胞抵御外界刺激的屏障之一,然而IL-34在低濃度時對SCAP增殖能力的促進作用是否與TNF-α有關,還需進一步研究。
綜上所述,本研究表明炎性因子IL-34對SCAP的增殖能力有一定的影響,為臨床治療年輕恒牙牙髓炎、根尖周炎提供一定的指導意義,但其發生的具體機制,還需后續實驗進一步研究。
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
