發酵肉制品中凝固酶陰性葡萄球菌的應用研究進展
2020-12-23 05:29:44王松郭柯宇唐林楊勇劉書亮
食品與發酵工業 2020年23期
王松,郭柯宇,唐林,楊勇,劉書亮*
1(四川農業大學 食品學院,四川 雅安,625014) 2(宜賓學院 生命科學與食品工程學院,四川 宜賓,644000)
傳統發酵肉制品生產采用自然發酵,微生物來自于原料和環境,在特定腌制和成熟工藝條件下,通過相互競爭形成典型本地微生物群落,從而顯著影響產品的質量和安全[1]。人工發酵劑的廣泛使用極大地促進了規模化生產的發展,滿足龐大消費市場對發酵肉制品的需求。然而,千篇一律的商用發酵劑使用造成了各地區傳統發酵肉制品典型風格逐漸弱化并趨同,規模化產品在消費者心中形成了單一的、乏味的,甚至是不安全的市場形象[2]。針對這一問題,功能性肉類發酵劑引起了越來越多研究者的興趣。這種新型發酵劑是由本地自然發酵產品中精心篩分的、具有多種新型功能的菌株組成,除了傳統發酵劑的基本酸化和顯色活性外,更重要的是能夠提高發酵肉制品風味、色澤、安全、健康多方面性能[3]。凝固酶陰性葡萄球菌(coagulase-negative staphylococci, CNS)是發酵肉制品中主要微生物類群之一,大量研究已經證明其在加工生產中具有重要作用,是功能性發酵劑的重要組成部分[4]。本文綜述了發酵肉制品中CNS的多樣性及其動態變化影響因素,CNS在發酵肉制品生產中的技術性能和潛在安全隱患,提出了新型功能CNS菌株的開發策略,為開展本地CNS菌株的應用研究和促進傳統發酵肉制品產業發展提供基礎信息。
1 發酵肉制品中的CNS
1.1 發酵肉制品中CNS多樣性
傳統發酵肉制品均是在自然環境條件下經歷了長時間的脫水和成熟,不同來源的多種微生物富集于肉基質中代謝繁殖,數量可達107~109CFU/g,其中乳酸菌(lactic acid bacteria, LAB)和CNS是最主要的細菌群落[5]。LAB是研究最多的一類微生物,清酒乳桿菌(Lactobacillussakei)、彎曲乳桿菌(Lactobacilluscurvatus)、植物乳桿菌(Lactobacillusplantarum)、戊糖片球菌(Pediococcuspentosaceus)是最常見的肉制品LAB[6]。然而有研究表明[7],在發酵肉制品中CNS的多樣性比LAB豐富,其多樣性和動態變化過程越來越受到研究者的重視。如木糖葡萄球菌(Staphylococcusxylosus)、腐生葡萄球菌(Staphylococcussaprophyticus)、馬胃葡萄球菌(Staphylococcusequorum)、肉葡萄球菌(Staphylococcuscarnosus)是歐洲傳統發酵香腸中最具優勢的種類[8-9](表1),而S.xylosus、S.saprophyticus、表皮葡萄球菌(Staphylococcusepidermidis)、松鼠葡萄球菌(Staphylococcussciuri)、模仿葡萄球菌(Staphylococcussimulans)等在中式發酵肉制品中更具有優勢[10-11]。S.xylosus和S.carnosus因具有較好的硝酸鹽還原作用、產香能力和安全性能成為最普遍的工業發酵劑菌種。除上述外,小牛葡萄球菌(Staphylococcusvitulinus)、溶血葡萄球菌(Staphylococcushaemolyticus)、巴氏葡萄球菌(Staphylococcuspasteuri)、琥珀葡萄球菌(Staphylococcussuccinus)、華納葡萄球菌(Staphylococcuswarneri)等也是發酵肉制品中常見的種屬,這些充分展示了肉中CNS的多樣性。另外,即使是同種內也存在大量的表型和基因型多樣性,比如S.equorum和S.saprophyticus顯示較高的種內遺傳多樣性,而S.xylosus種內遺傳多樣性低[9]。
1.2 影響發酵肉制品CNS群落動態變化的因素
CNS群落在發酵肉制品中受到包括原輔料、加工過程、加工條件等一系列復雜因素的影響,目前仍未能完全解析其動態變化規律。比如LEROY等[18]發現S.equorum在所有環境和肉樣中均有分布且比例均勻,S.saprophyticus在環境和肉樣中均有分布,但環境樣品中含量更高,S.xylosus在部分環境和肉樣中存在,且肉樣中含量更高。溫度、pH、煙熏等加工工藝對葡萄球菌的動態變化影響比原料更大。研究顯示S.carnosus在原料肉(如豬肉)中一般很少存在,但由于其在高鹽含量和低pH (pH<5.5) 的肉基質中具有更好的競爭優勢,反而成為肉制品中優勢種群,尤其是高酸化的發酵香腸中[19]。S.xylosus更適合低溫與溫和的酸度條件,S.epidermidis更適應較高溫度的環境條件,而S.carnosus更適應較低pH的環境條件[20]。S.saprophyticus是煙熏產品中優勢種屬,但成熟過程會逐漸消耗前期代謝積累的乳酸等有機酸,引起pH值升高,導致優勢種群向S.equorum轉變,因為后者對低酸性的環境更加適應[21]。另外,不同加工類型產品中主要優勢菌群也有差異,LANDETA等[22]研究西班牙香腸和火腿中CNS發現,S.carnosus在香腸中是主要菌群,而S.equorum是火腿中的主要菌群。

表1 部分中西式發酵肉制品中CNS優勢種屬Table 1 Dominant species of CNS in parts of Chinese and Western fermented meat products
2 CNS在發酵肉制品生產中的技術性能
2.1 硝酸鹽還原

2.2 風味形成
CNS的代謝活動對風味的形成至關重要,分為4種主要的作用模式[28]:(1)碳水化合物發酵;(2)氨基酸轉化反應;(3)脂質β氧化;(4)酯酶活性。肉中主要是LAB發酵碳水化合物產生乳酸,引起肉基pH值下降。然而,CNS也能夠在一定程度上代謝碳水化合物,并將其轉化為有機酸(主要是乳酸和醋酸)以及揮發性化合物,如雙乙酰、乙醛、乙偶姻等,可以帶給干發酵香腸奶油味或奶香味[29]。除了碳水化合物代謝外,CNS代謝氨基酸產生具有低閾值的揮發性成分對發酵肉制品風味更有決定性的作用。動物死后,肌漿蛋白和肌纖維蛋白可被肌肉蛋白酶和肽酶水解。除了這些內源酶外,CNS的外源蛋白酶活性也很明顯,尤其是肽的水解及其轉化為游離氨基酸的酶活性,這可能占到總蛋白水解的40%,肽和游離氨基酸都可以通過額外的反應形成揮發性和非揮發性化合物[30]。CNS可以將支鏈氨基酸包括纈氨酸、亮氨酸和異亮氨酸通過轉胺或脫羧形成相應的支鏈醛、羧酸和醇,比如來源于亮氨酸的化合物3-甲基丁醇、3-甲基丁醛和3-甲基丁酸常與發酵香腸的香氣聯系在一起[31-32]。同樣,脂質水解主要也是內源性脂肪酶作用于脂類物質完成的,但CNS的外源脂肪酶也有一定的作用[33]。脂類水解釋放出的脂肪酸可以被CNS產生的外源性脂肪酶通過不完全β氧化降解成ketoacyl-CoA,再在硫酯酶的作用下脫去酰基形成β-酮酸、短鏈游離脂肪酸和CoA。CNS可進一步將β-酮酸經脫羧作用后生成肉制品典型風味成分甲基酮,如2-戊酮、2-己酮、2-庚酮等[34]。另外,這些甲基酮可以進一步轉化為仲醇,CNS的酯酶活性還可以將相應的醇和酸進一步反應,生成芳香酯化合物[35]。
2.3 抗氧化活性
肉類產品的氧化可使脂質和蛋白質降解,而過度氧化則導致最終產品的顏色,質地,風味和營養價值的降低。脂質過度氧化可導致形成與不良風味相關的醛,酮和羧酸等物質[36],蛋白質過度氧化可導致羰基化合物和蛋白質聚集體的形成,進而降低肉制品成熟過程中蛋白水解作用,或者直接損害蛋白水解酶如半胱氨酸蛋白酶的活性,減少游離氨基酸產生和進一步轉化,降低肉制品品質[37]。發酵肉制品中的CNS具有較強的超氧化物歧化酶(superoxide dismutase,SOD)和過氧化氫酶的活性,SOD將超氧自由基解毒成反應性較低的過氧化氫,再被過氧化氫酶轉化為水和氧氣,減緩肉制品的過度氧化過程。目前的研究表明,大多數CNS至少具有1種過氧化氫酶活性,一部分種屬如S.carnosus、S.saprophyticus、S.equorum存在2種過氧化氫酶活性[38]。VERMASSEN等[39]發現木糖鏈球菌存在多達3種過氧化氫酶活性,當在肉基質中加入亞硝酸鹽后,S.xylosus抗氧化活性的編碼基因在亞硝化脅迫下顯著上調,抗氧化活性增強。
3 潛在的安全隱患
食源性CNS菌株與臨床CNS菌株種間和種內的遺傳相似性極低,發酵肉制品中CNS的代謝活性通常不會引起安全隱患,目前商業菌株S.xylosus和S.carnosus被認為是安全的[40],我國在2016年把S.xylosus、S.carnosus和S.vitulinus列入可用于食品的菌種名單[41]。西方肉制品加工業開始逐漸認可S.equorum應用于工業生產[22],另外一些種屬如S.saprophyticus、S.epidermidis、S.simulans的特定菌株也被應用于生產實驗中,并表現出穩定的安全性能[17,42]。但是,CNS的某些菌株可能仍然表現出威脅食品安全的不良特性,在作為人工發酵劑添加時,由于人為增加菌體濃度,可能會放大這些不良特性的威脅[40]。因此,對CNS進行安全性評估是重要且必需的,尤其是在將其用作食品發酵劑時,需要采取逐案處理的方法[43]。目前對CNS菌株安全性評估工作主要包括是否能夠產生生物胺、是否具有毒力因子、是否具有可轉移的抗生素抗性等。
3.1 產生生物胺
過量攝入生物胺可對人體產生負面的生理影響,當肉制品中的生物胺含量高于 200 mg/kg 時,可能會導致食物引起的偏頭痛和高血壓。CNS菌株通常存在氨基酸脫羧酶活性,因此可能具有產生生物胺的能力,研究結果[44]也證實部分CNS菌株能夠產生低濃度的生物胺,且不同種屬和不同菌株之間的水平有所不同。如STAVROPOULOU 等[45]發現在營養豐富的培養基中孵育48 h后,不同CNS物種的生物胺濃度大多仍低于100 μmol/L,其中S.carnosus所產生的2-苯乙胺是含量最高的生物胺類,但其濃度含量仍然在安全范圍之內。目前,肉制品中受到生物胺風險的例子極少,但當有其他產生物胺細菌污染時,仍具有相當的風險,確保所用發酵劑菌株不產生物胺仍是非常必要的。
3.2 毒素因子
CNS的另一個潛在安全風險是可能產生葡萄球菌腸毒素(staphylococcal enterotoxins, SE),目前至少有23種血清學上不同的肽,并且對熱和大多數蛋白水解酶具有抗性[4]。科學家們對金黃色葡萄球菌SE進行了深入研究,但仍然對CNS腸毒素引起的食物中毒存在疑惑。TALON等[40]指出迄今為止尚無關于僅涉及CNS菌株的食物中毒的報道。PODKOWIK等[46]在32株S.epidermidis中發現8株菌有編碼SE的基因,其中1株菌的編碼基因可以進行穩定遺傳,并進一步證明該編碼基因在包括肉湯在內的多種食品中可以表達并產生金黃色葡萄球菌同源SE。因此,用于發酵肉制品的CNS菌株必需確保不產任何SE。
3.3 抗生素耐藥
抗生素耐藥的編碼基因(例如blaZ,tetK,tetM,lnuA,ermB和ermC)常常位于可轉移或可移動的質粒上,從而使抗生素耐藥成為應用于食品發酵細菌的主要安全風險[12]。發酵肉制品CNS中包含可轉移的抗生素耐藥基因通常對青霉素、紅霉素、四環素和林可霉素具有耐藥性,成為人體細菌耐藥基因的儲存庫和載體[47]。CNS的抗生素耐藥型與菌株來源無關,不同種屬、不同菌株之間可以表現出較大的差異,如一些菌株表現出完全缺乏抗藥性(56%),而另一些菌株(2%~7%)可能對多達6種抗生素具有抗藥性[13];又如S.xylosus的多重耐藥比較少,而S.epidermidis的多重耐藥比較多[14]。文獻報道最多的抗生素耐藥CNS是S.xylosus和S.carnosus,當然這與這些商用菌株研究比較多有關,但也側面反應了目前這些商用菌株耐藥基因比例逐漸升高,迫切需要找到新的商用菌株。而自然發酵肉制品中的抗生素壓力較低甚至沒有,是優良菌株的重要來源。
4 新型功能性CNS的篩選策略
合理篩選具有特定功能的CNS菌株將有助于針對性地改善發酵肉制品的質量和安全性能,滿足消費者和生產者迫切需求。針對現階段工業生產存在問題,著重篩選可以改善最終產品的風味、顏色、生物安全的CNS菌株,以及加工過程中在肉類基質具有競爭優勢的菌株,應用于生產提升產品品質與安全。
4.1 增強競爭力
一部分CNS菌株能夠在肉類基質碳水化合物水平較低時(10~20 mg/g)利用其他替代能源,比如核苷和精氨酸,從而在不同程度上增強其競爭優勢[48]。這種競爭優勢對CNS在肉類基質中實現其功能特性是極其重要的,因為只有菌體細胞生長發展到足夠的數量,表達的功能特性才能對產品產生顯著性影響。
動物機體死亡后,肌肉細胞中腺嘌呤核苷三磷酸(adenosine triphosphate,ATP)可以自動分解釋放出腺苷和肌苷,濃度可以達到300 mg/100 g肌肉,這些核苷可以通過特定的轉運蛋白吸收到細菌細胞中,被菌體細胞內核苷水解酶分解形成核糖,當肉基中其他碳源和能源不足時,這些核糖可以作為替代能源被肉中CNS代謝[49]。JANSSENS等[48]驗證了分屬于17個種的61株CNS菌株利用核苷代謝情況,證明了代謝利用核苷是CNS菌株的普遍能力。除此以外,CNS還擁有利用精氨酸作為替代能源的能力。精氨酸大量存在于肉中,可達到6 mg/100 g肌肉,且隨著肉制品加工過程可增加到原濃度的3倍。由圖1[4]可知,細菌通過精氨酸脫亞氨基酶(arginine deiminase,ADI)途徑將1 mol精氨酸轉化為1 mol鳥氨酸、1 mol CO2、2 mol NH3和1 mol ATP[50],該途徑產生的ATP和NH3可以提高細菌在發酵食品中競爭優勢[4]。ADI途徑在CNS中的表達活性在種和菌株2個水平上均存在顯著差異[48]:種水平上,S.carnosus、S.condiment、S.epidermidis、S.haemolyticus、S.hyicus、S.saccharolyticus和S.simulans普遍具有ADI活性,而S.lugdunensis、S.lutrae、S.sciuri和S.succinus中則較少出現;菌株水平差異較多出現在S.auricularis、S.hominis、S.intermedius、S.pasteuri,S.saprophyticus和S.Warneri中。JANSSENS等[48]還發現ADI途徑處于多種調控水平,表達活性在菌株水平上觀察到的差異還常常受到外界環境因素的影響,甚至可能導致其轉換為其他精氨酸代謝途徑,如當生物量較低時,ADI表達活性似乎受到了阻礙,有氧運動可能導致某些菌株向精氨酸酶活性的轉換,后者是精氨酸轉化的替代途徑,導致鳥氨酸和尿素形成。
盡管目前這些替代能源代謝機制尚未完全搞清楚,其代謝作用對CNS在肉基質中競爭優勢是否起著顯著作用,以及這種代謝能力在不同菌株間是否存在顯著性差異還需要進一步研究。但精心篩選在特定肉基質中具有較強競爭優勢的菌株,有助于新型發酵劑更好地表達功能特性,顯著改善產品質量與安全。
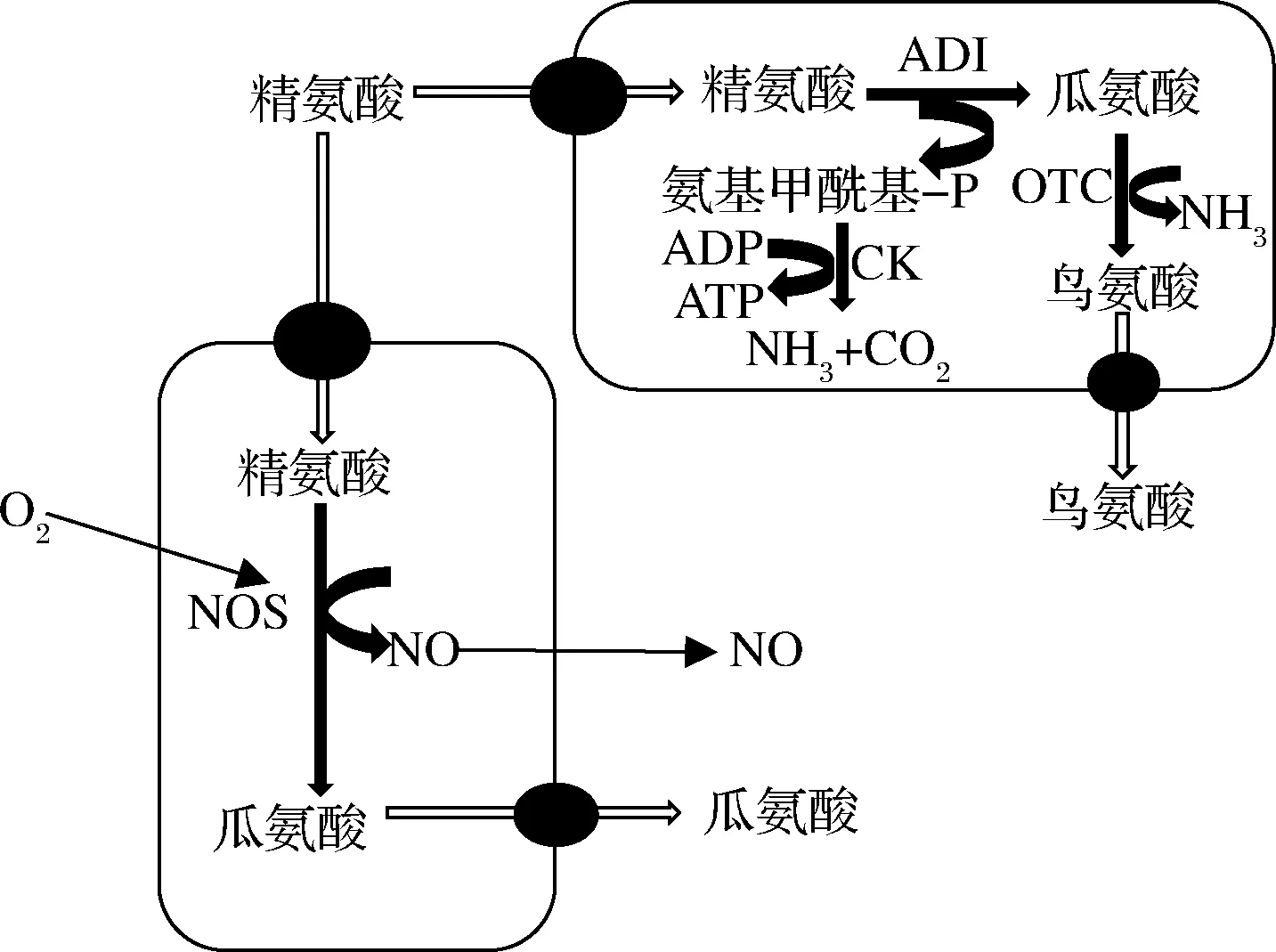
ADI-精氨酸脫亞氨基酶;OTC-鳥氨酸甲酰基轉移酶;CK-氨基甲酸激酶;NOS-一氧化氮氧合酶圖1 凝固酶陰性葡萄球菌中精氨酸代謝途徑Fig.1 Pathway of arginine metabolism in coagulase-negative staphylococci
4.2 顏色產生
如前所述,CNS普遍具有硝酸鹽還原酶活性,但硝酸鹽還原酶活性在CNS不同種之間和同一種屬不同菌株之間均存在較大差異。比如S.carnosus、S.xylosus、S.equorum、S.lentus、S.simulans、S.sciuri和S.succinus常具有較強的硝酸還原酶能力,而S.saprophyticus、S.succinus和S.warneri通常不存在硝酸鹽還原酶活性。BOSSE等[25]發現接入硝酸鹽還原酶活性較強的S.carnosus可顯著減少火腿中硝酸鹽殘留量,增加亞硝酸鹽含量;而接入酶活力較弱的S.carnosus則相反。MAINAR等[27]在添加2.96 mmol/L硝酸鹽的肉模擬培養基中評估88株CNS,發現其中78個菌株具有硝酸鹽還原酶活性,但只有35株菌表現出較高轉化活性,分屬于S.carnosus、S.epidermidis、S.equorum、S.pasteuri和S.xylosus。另一方面,添加具有硝酸還原酶活性的CNS菌株并不能確保肉制品良好色澤的形成,加工工藝條件對色澤的形成同樣具有重要的影響。研究表明CNS的硝酸鹽還原酶活性一般是在硝酸鹽存在的情況下由厭氧環境條件誘導,且在指數生長期間達到最大,只有適宜加工條件使菌株形成競爭優勢,才能最大化的表達其硝酸鹽還原酶活性[24]。GOTTERUP等[26]確認CNS的硝酸還原酶活性不能直接預測香腸制品中亞硝基肌紅蛋白的形成率,菌株良好顏色形成能力的預測還應包括生長特性和耐酸性等其他相關特性。
近年來,科學家們提出CNS還可以通過一氧化氮合成酶(nitric oxide synthase, NOS)途徑代謝精氨酸產生Nω-羥基-精氨酸中間產物,隨后形成NO和瓜氨酸(圖1)。RAS等[51]驗證了S.xylosus中確實存在nos基因,并發現大多數CNS菌株中都含有nos基因,推測其在CNS中高度保守,但是表達并產生的NO無法形成足夠的亞硝基肌紅蛋白,反而更多的是形成氧合肌紅蛋白,且在種和菌株間存在較大變異[52]。從理論上講,發酵肉制品中細菌NOS途徑可用作NO的替代產生途徑,用來減少硝酸鹽和/或亞硝酸鹽在肉制品生產中的使用。然而,細菌NOS途徑更多的是用于抵御氧化應激,其所產生的NO是否能夠用來合成足夠的亞硝基肌紅蛋白,進而在生產中替代亞硝酸鹽還缺乏足夠的研究[53]。MAINAR等[27]研究17個種的86株CNS發現,只有1種菌株顯示NOS活性且活性較弱。現有證據表明,基于NOS的肉制品良好色澤形成還有很多科學問題需要解決,但仍為尋找合適的亞硝酸鹽替代開辟了新的方向。
4.3 風味強化
風味是肉制品最重要的質量屬性,如前所述,CNS的生長代謝作用可以將肉中的碳水化合物、氨基酸和脂肪酸代謝轉化,產生一系列揮發性物質,從而改善產品風味。目前商業菌種S.xylosus和S.carmnosus在風味形成方面的作用研究很多,S.xylosus菌株更適用于發酵香腸生產,所產生的芳香族風味物質可以為產品帶來更加圓潤的風味,而S.carmnosus可能比其他CNS產生更多的甲基酮以及支鏈醛和醇類,導致成熟時間更快[54-55]。盡管如此,仍需要更多研究來弄清哪種發酵劑更適合某一特定類型的發酵肉產品,以及硝酸鹽添加量,發酵時間、發酵溫度,鹽含量和發酵劑濃度等工藝差異對風味的影響[19-20, 32, 56]。已有學者對CNS在蛋白質和脂肪降解、風味物質產生等方面的種間和種內差異性進行了大量研究,除了現有商用發酵劑外,一些CNS種屬如S.equorum、S.saprophyticus、S.sciuri、S.warneri和S.simulans的部分菌株也展示了良好的揮發性風味物質生產潛力,可為系統評估發酵肉制品的本地CNS群落和篩選優良風味功能CNS菌株奠定了重要基礎[42],如S.saprophyticus和S.warneri在一定的生產條件下,生產乙偶姻、二乙酰、乙酸等重要風味成分的能力可以達到甚至超過商業發酵菌株S.xylosus的水平,而S.carnosus則幾乎不產這類風味物質[45]。可以肯定的是,自然發酵過程利用本地野生型菌株的特定代謝作用,能夠為產品帶來誘人的獨特香氣,近年來分子技術和發酵技術的進步為我們重新評估這些本地野生型菌株提供了可能[57]。因此,篩選本地野生型風味改良菌株作為新型功能發酵劑可能是改善產品風味的有效途徑。
4.4 生物保護
篩選產細菌素的菌株作為發酵劑用于肉制品生產,提高產品對有害微生物的抵抗作用,保障發酵正常進行,減少致病菌的危害,成為肉制品功能發酵劑研究的重點[58]。近年來,肉制品發酵劑中另一種重要微生物LAB在這方面有大量的研究,對細菌素的功能、微生物產細菌素的機理和功能微生物的篩選等相關內容進行了較為深入的闡述,本文在此不再贅述。現有的研究表明,CNS菌株同樣具有產生細菌素的能力,且與LAB不同的是,部分CNS細菌素展示了對梭狀芽孢桿菌和/或金黃色葡萄球菌具有較好的抑菌活性[59]。加強CNS產細菌素方面的研究工作,篩選產細菌素CNS與產細菌素LAB(對李斯特氏菌有較好的抑制作用)聯合作為發酵劑應用于肉制品生產,可以更好地提升產品的質量和安全,在工業生產中適量減少硝酸鹽/亞硝酸鹽的用量。
5 結束語
隨著發酵工程和分子生物學的不斷發展,古老的傳統肉制品行業也在不斷進行技術創新,以適應現代化生產的需求,提高產品感官特性和食品安全性,滿足消費者需求。目前,對CNS的研究主要關注了商用菌種S.xylosus和S.carmnosus增強風味和穩定色澤等常規技術性能方面的作用,然而對CNS在不同肉制品生態中的多樣性及其功能代謝差異還研究不多。開展本地CNS的生態學研究,將工業生產需求和傳統肉制品特定風格需求結合起來,有針對性的篩選功能性CNS發酵劑菌株,例如增強特定風味、顯著改善顏色形成、提高生物安全性等,將其應用于傳統肉制品的大規模生產,滿足生產和消費兩方面的需求。
