南極紅藻Iridaea cordata和Curdiea racovitzae轉錄組分析及其極端光環境適應相關基因的挖掘
2020-11-16 03:41:08劉晨臨王秀良林學政
海洋學報 2020年10期
劉晨臨,王秀良,林學政
( 1. 自然資源部第一海洋研究所,山東 青島 266061;2. 中國科學院海洋研究所,山東 青島 266071;3. 青島海洋科學與技術試點國家實驗室 海洋生物學與生物技術實驗室,山東 青島 266237)
1 引言
生活在南極的大型海藻約有120~130種,其中有三分之一的是南極特有的[1]。在西南極半島,大型海藻廣泛生長在潮間帶或淺岸海底的礁石上。喬治王島(King George Island)的梅爾維爾角(Melville Cape),90%以上的巖石底層被海藻覆蓋。而在將軍灣(Admiralty Bay),夏季大型海藻帶幾乎環繞著整個海灣沿岸,覆蓋了約37%海灣面積。褐藻和紅藻在潮下帶巖石基底上呈垂直分布,最深可達100 m水深處[2–3]。大型海藻是近岸海洋系統重要的初級生產者,為其他海洋生物提供庇護和食物來源[4]。除生態學意義,許多大型海藻也具有較高的經濟價值,目前對南大洋海藻資源的開發利用日益成為熱點[5]。產膠紅藻Iridaea cordata(Turner)Bory和Curdiea racovitzaeHariot是南極和亞南極海岸潮間帶和潮下帶的常見物種[6]。I. cordata產卡拉膠[7],其硫酸多糖具有抗癌活性[8],C. racovitzae則是產瓊膠海藻[9]。
許多來自南極的物種只在低于5℃的溫度條件下生長和繁殖。低溫需求決定了它們在南極的北部分布邊界。I. cordata的最適生長溫度在0~5℃,生存溫度的上限為11~16℃[10]。除冰凍和低溫外,海藻的生長發育也被南極地區多種極端的光環境影響。夏季,潮間帶海藻暴露在強烈的紫外線輻射下,需要有效的紫外線傷害的防御和修復能力[11–12]。在I. cordata中發現了能夠吸收紫外線,降低紫外輻射傷害的類菌胞素氨基酸[13]。而冬季由于極夜和海冰覆蓋,黑暗時間較長,海藻生長必須適應弱光環境,降低其光合作用和生長對光照的需求量[14–15]。在南極漫長黑暗的冬季,因為具有較高的光合效率和極低的光補償點和光飽和點,I. cordata仍然能夠持續生長[10]。
目前,對南極大型海藻的研究主要集中在生態分布、生物量和生物多樣性[2–3,6,15–16]、功能活性物質的發現[17–18],以 及 光 合 生 理 活 性[10–12,19–20]等 方 面。迄 今 為止,除了南極綠藻Prasiola crispa的轉錄組數據[21],未見其他南極海藻功能基因或轉錄組學研究的報道。南極藻類因其特有的耐受極端環境的能力,成為發現新基因和新代謝機制的理想材料。而轉錄組測序和分析是發現這些基因資源最經濟、有效的手段。我們從兩種南極紅藻的轉錄組序列中,挖掘了一些獨特的可能與其南極極端光環境,包括弱光和強紫外線輻射等適應相關基因,為后續開展南極紅藻極端環境適應機制研究提供了依據。同時從南極紅藻中發現的光修復酶基因,在化妝品領域也有開發利用的前景。
2 材料與方法
2.1 海藻樣品
2018年2 月,在第34次中國南極科學考察中,依托“向陽紅01”號科學考察船,在西南極南設得蘭群島喬治王島的長城站附近(62.216°S,58.960°W)潮間帶采集紅藻樣品。采樣時陰天有小雨,氣溫約2℃。用無菌海水清洗藻體后,置于船上?150℃冰箱保存。保持冷凍狀態至回國后,在實驗室提取藻樣的總RNA。
2.2 總RNA提取和cDNA文庫的構建
首先將冷凍的海藻樣品浸入液氮中,用冷卻的研缽研磨成細粉。使用天根植物RNAprep試劑盒提取總RNA。采用微量核酸定量儀檢測RNA的質量和濃度。測序文庫使用TruSeq RNA樣品制備試劑盒(Illumina, USA)構建。用Illumina PCR引物進行15個循環的PCR反應,選擇性富集兩端連接接頭分子的DNA片段,并用AMPure XP系統純化(Beckman Coulter,USA)。采用安捷倫(Agilent)生物分析儀2100系統中的高靈敏度DNA分析技術對產物進行定量。Illumina Hiseq測序委托上海派森諾生物技術有限公司完成。
2.3 轉錄組數據組裝和功能注釋
在數據組裝前先過濾原始數據。使用Cutadapt(v1.15)軟件刪除引物或接頭序列。過濾掉超過10%堿基質量低于Q20的或未知核苷酸(Ns)堿基超過5%的序列。使用Trinity (v2.5.1)軟件對轉錄本進行拼接[22]。選擇每個基因最長的轉錄本作為該基因的代表性序列稱為非冗余基因。BlastX同源搜索NCBI非冗余蛋白數據庫NR (http://www.ncbi.nlm.nih.gov),E-value<10–5。利用真核生物直系同源蛋白質聚類EggNOG(Evolutionary Genealogy of Genes: Non-supervised Orthologous Groups,http://eggnog.embl.de/version_3.0/)對非冗余基因進行功能分類注釋[23]。利用京都基因與基因組百科全書KEGG(Kyoto Encyclopedia of Genes and Genomes,http://www.kegg.jp/)進行代謝通路注釋,獲得物種內分子間相互作用和反應的網絡。KEGG代謝通路注釋以雙向最佳匹配(BBH)的方法,采用KAAS軟件完成[24]。使用PrimerPro(http://webdocs.cs.ualberta.ca/~yifeng/PrimerPro/)的微衛星搜索模塊(MISA)篩選簡單重復序列(SSRs)。
2.4 系統進化樹的構建
基因家族系統進化樹的構建用MEGA X軟件完成[25],將選擇的序列經MUSCLE比對后,采用默認參數,用鄰接法構建系統樹。經過1000次自展值的檢驗,顯示一致性的系統樹。
3 結果與討論
3.1 南極紅藻Iridaea cordata和Curdiea racovitzae轉錄組組裝和注釋
南極紅藻I. cordata和C. racovitzae的形態如圖1所示。紅藻的鑒定采用克隆的18S rRNA基因與Gen-Bank已知序列進行比對,相似性為100%。
每種紅藻的轉錄組測序量約為6 G,I. cordata和C. racovitzae的轉錄組序列分別拼接成了14055條和12006條非冗余基因(表1)。非冗余基因的平均長度分別為1473 bp和1448 bp。I. cordata的鳥嘌呤和胞嘧啶(GC)含量約為53.82%,C. racovitzae的GC含量約為47.28%。GC含量在一定程度上反映了基因組的熱穩定性。角叉菜Chondrus crispus的GC含量約為55.6%[26],而龍須菜Gracilariopsis lemaneiformisGC含量為48.13%[27]。南極紅藻的GC含量與常溫紅藻的GC含量差別不大。
I. cordata和C. racovitzae轉錄組序列在各個數據庫的注釋情況見表2。在NR數據庫,I. cordata和C.racovitzae分別有4788條(34.07%)和4468條(37.21%)非冗余基因得到注釋。在Swissprot數量庫中I. cordata和C. racovitzae則分別有4864(34.61%)和4938(41.13%)條非冗余基因得到注釋。紫菜Pyropia tenera轉錄組序列在UniProt數據庫(不含其他紫菜屬物種的基因組和轉錄組序列)中也僅有33.6%的重疊群匹配到同源序列[28]。這說明了紅藻基因與公共數據庫中已知基因序列間的同源性較低,繼續豐富不同種類紅藻基因數據資源是很有必要的[29]。

圖1 南極紅藻Iridaea cordata (a)和Curdiea racovitzae (b)的形態Fig. 1 The morphology of Iridaea cordata (a) and Curdiea racovitzae (b)
I. cordata和C. racovitzae中被EggNOG注釋的非冗余基因分別有4523和4238條(圖2)。除了一般功能預測和功能未知的分類,翻譯后修飾、蛋白反轉和分子伴侶類別的非冗余基因(579,581)最多,其次依次是翻譯、核糖體結構和起源(485,485)、信號傳導機制(389,321)、碳水化合物轉運和代謝(301,280)、能量的產生和轉換(259,279)、RNA加工和修飾(237,221)、復制、重組和修復(209,176)、細胞內轉移、分泌和囊泡轉運(209,220)、氨基酸轉運和代謝(201,209)以及轉錄(198,193)等分類。
3.2 南極紅藻轉錄組序列與常溫近緣紅藻基因組序列的比較
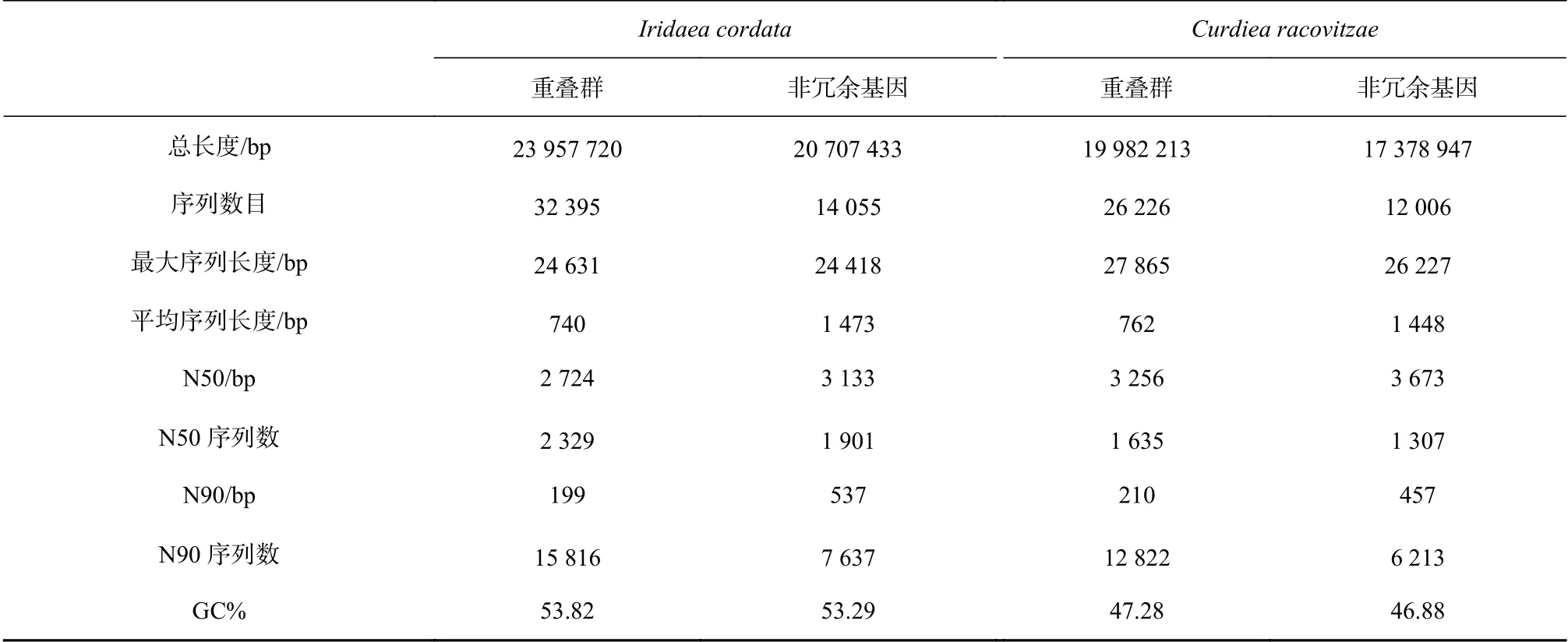
表1 南極紅藻Iridaea cordata和Curdiea racovitzae轉錄本和非冗余基因Table 1 The contigs and unigenes of Iridaea cordata and Curdiea racovitzae
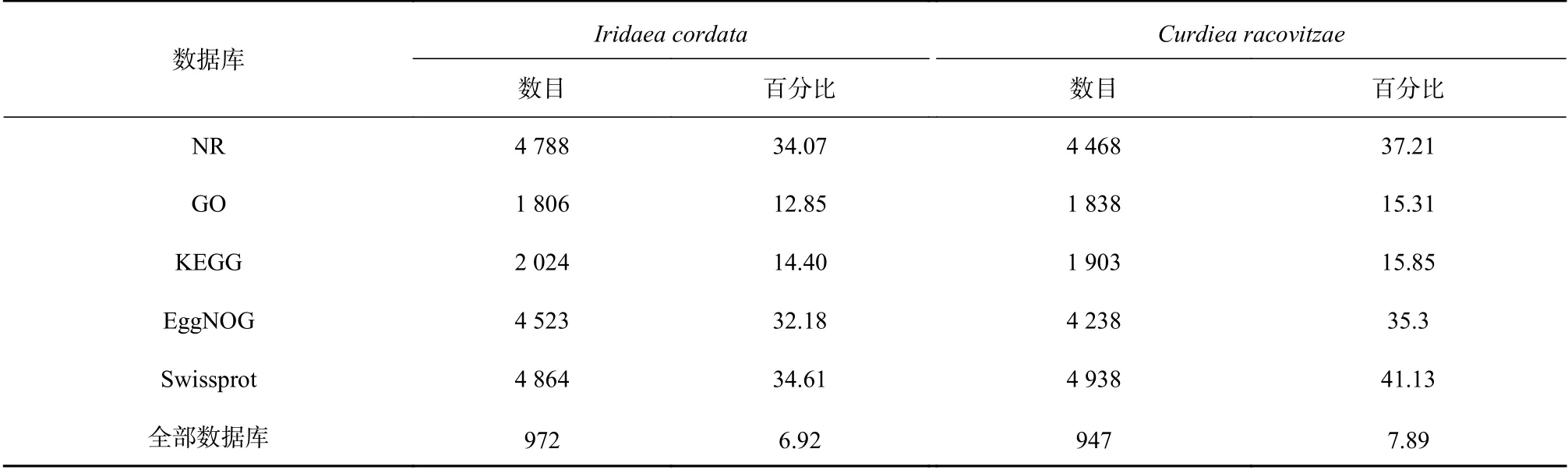
表2 Iridaea cordata和Curdiea racovitzae轉錄組序列在各個數據庫的注釋結果Table 2 The annotation results of Iridaea cordata and Curdiea racovitzae transcriptome
在已完成全基因組測序的紅藻中,角叉菜與I. cordata都屬于杉菜目,親緣關系最近。龍須菜與C.racovitzae同屬于江蘺目,有較近的親緣關系。我們將所測的兩種南極紅藻的轉錄組序列分別與角叉菜(基因組GenBank序列收錄號:PRJEA78309、PRJNA 193762)和龍須菜 (PRJNA361418) 的基因組的蛋白編碼序列用本地BlastX(E-value<10?5)進行比較,發現有5955條I. cordata的非冗余基因與4059條角叉菜的序列匹配。C. racovitzae的4964條非冗余基因匹配到了3596條龍須菜基因序列(圖3),但是兩兩物種間,互相不匹配的序列占有更大比例,說明同目的紅藻間遺傳差異較大。
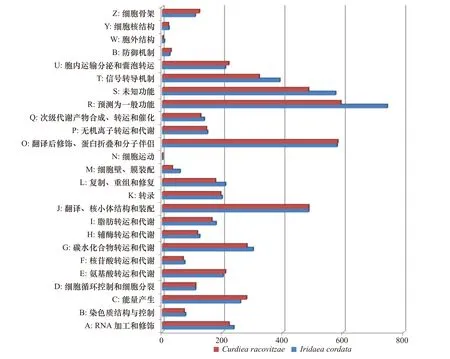
圖2 Iridaea cordata和Curdiea racovitzae轉錄組序列的EggNOG功能分類Fig. 2 EggNOG function classification of Iridaea cordata and Curdiea racovitzae
3.3 捕光復合物基因家族
在光系統中,捕光復合物LHC (Light-Harvesting Complex) 負責捕獲光能并將能量轉移到反應中心。除了吸收光,這些蛋白質還參與了光保護反應,以清除光合作用過程中產生的活性氧[30]。我們從紅藻角叉菜、龍須菜和耐受酸熱環境的單細胞紅藻Galdieria sulphuraria(PRJNA13023) 的基因組序列,以及兩種南極紅藻轉錄組序列中,通過BlastP比對、KEGG注釋分析和Pfam注釋查找有CB (Chloro a/b-bind) 結構域的序列[31],篩選了假定的LHC家族基因,利用這些基因編碼的蛋白序列構建了系統樹(圖4)。之前的研究認為紅藻中僅有光系統I(PSI)相關的,與葉綠素a和類胡蘿卜素結合的LHC天線蛋白,稱為LHCR[32–33]。LHCR的數目在不同紅藻間存在差異,蛋白分離純化的結果表明紫球藻有6種不同的LHCR蛋白,G. sulphuraria有5種,而Cyanidioschyzon merolae只有3種[33]。
從角叉菜、龍須菜和G. sulphuraria的基因組序列中各找到了5、13和10條Lhc序列。角叉菜的5條Lhcr基因均編碼紅藻型LHC蛋白,龍須菜有7條Lhcr基因,6條其他類型的Lhc基因。G. sulphuraria中有9條Lhcr基因和1條其他類型的Lhc基因。從I. cordata和C. racovitzae轉錄組中分別找到了8條和5條Lhc基因。分別包括4條Lhcr和1條Lhcf基因。Lhcf編碼巖藻黃質和Chla/c結合蛋白,屬于PSI天線復合體。LHCF是硅藻和褐藻中的主要天線蛋白,但還未見在紅藻中的報道[34–35]。此外,I. cordata轉錄組中還包含多條與綠藻LHC蛋白同源的序列,參照萊茵衣藻LHC的分類,分別為Lhca2,Lhca6和Lhcb基因(圖5)。在萊茵衣藻中,LHCa2和LHCa6屬于PSI,而LHCb屬于PSII。
目前關于LHC蛋白的進化還缺少切實的證據,一種觀點認為,紅藻、綠藻和高等植物Lhc基因家族是共同起源,獨立進化的[32]。在進化過程中,由于藻膽體的丟失,有些Lhca(LhcI) 的基因功能進一步分化,成為PSII的天線蛋白[33]。綠藻中編碼Chla/b-和Chla/c結合蛋白的基因在LHCI / LHCII分化之前就已經彼此分離[36]。LHCb是植物生長和光保護反應的關鍵蛋白。目前對PSII中LHC蛋白的研究主要在高等植物和綠藻中,而其他物種中則少見報道[36]。對于南極紅藻I. cordata特有的Lhc基因的蛋白產物還需要蛋白分離純化等生物化學的方法進行驗證。但是Lhc基因家族的擴張能夠增強I. cordata在光限制環境中的光吸收和能量轉移能力,可能是適應南極冬季弱光環境生存的關鍵。

圖3 角叉菜Chondrus crispus與Iridaea cordata,龍須菜Gracilariopsis chorda與Curdiea racovitzae匹配基因數目的韋恩圖Fig. 3 Diagram of the matching gene numbers between Chondrus crispus and Iridaea cordata, Gracilariopsis chorda and Curdiea racovitzae
3.4 光裂解酶基因家族
環境非生物脅迫,如干旱或紫外線,可誘導藻類DNA損傷。目前發現的兩種光裂解酶,環丁烷嘧啶二聚體(CPD)光裂解酶和6?4光裂解酶,可以特異性地修復不同類型的UV誘導的DNA損傷,CPD和6?4嘧啶酮光產物 (6?4 PP)[37]。光裂解酶屬于紫外線/藍光激活蛋白家族—光裂解酶/隱花色素(CPF)家族,這個超家族包括光裂解酶、隱花色素(Cry)和Cry-DASH。這三類成員有很高的同源性,都含有能結合兩個生色團的PHR(Photolyase Homology Region)功能區[38],但是Cry-DASH和隱花色素的PHR無光裂解酶活性[39]。
從紅藻角叉菜、龍須菜和G. sulphuraria的基因組序列,以及南極紅藻的轉錄組序列中通過Pfam注釋,選取了所有含PHR功能區的序列,作為假定的光裂解酶/隱花色素CPF家族蛋白序列,同時參考Fortunato等[40]構建的CPF超級家族系統樹,選擇其中的綠藻、硅藻、褐藻和擬南芥的序列,與我們篩選的紅藻CPF家族蛋白序列一起構建了的系統進化樹,并對進化樹的分支進行了功能劃分(圖6)。I. cordata的轉錄組序列中有4條CPF家族基因,分別屬于6?4光裂解酶,光裂解酶CPD I和CPD II家族。C. racovitzae轉錄組序列中有6條CPF家族基因,分別編碼隱花色素Cry、Cry-DASH、Cry-DASH II和光裂解酶CPD II。6?4光裂解酶分支的物種,包括紅藻、硅藻、褐藻、綠藻和高等植物。Fortunato等[40]未在角叉菜中發現6?4光裂解酶基因,認為只有單細胞紅藻中有6?4光裂解酶,但是我們在除C. racovitzae之外的所有紅藻中,都找到了6?4光裂解酶同源序列,說明其在紅藻中是廣泛存在的。光裂解酶CPD,分為2個分支,CPD II廣泛分布于各類生物物種[40],與我們的結果一致。而CPD I則主要存在于細菌、古細菌、藍細菌和真菌中[37,40]。Fortunato等[40]未在灰色藻、甲藻和紅藻角叉菜中找到CPD I家族基因,但是在我們構建的系統進化樹中,在紅藻I. cordata,龍須菜以及硅藻中都找到了CPD I的同源序列。我們認為在少數種類的硅藻和紅藻中存在的CPD I基因可能是通過水平基因轉移從原核生物中獲得的,用于修復其因生活環境嚴酷而造成的DNA損傷。在I. cordata轉錄組中未找到紅藻中普遍存在的Cry和Cry-DASH序列,同時在C. racovitzae轉錄組中未發現6?4光裂解酶,可能是轉錄組測序未能完全覆蓋基因組中所有編碼序列導致的。
3.5 簡單重復序列SSR
從轉錄組序列中篩選的SSR序列,因為其豐富性且與編碼基因緊密相關,成為在物種適應性、生物地理分布和進化研究中的重要分子標記。利用MISA軟件分別從I. cordata的3014條非冗余基因中鑒定出5560個SSR位點,從C. racovitzae的1636條非冗余基因中鑒定出2473個SSR位點。其中單核苷酸重復最多。紫菜P. seriata中三核苷酸的數目最多[29]。SSR的頻率和分布被認為取決于各種因素,如數據集的大小、工具和使用的標準[41]。
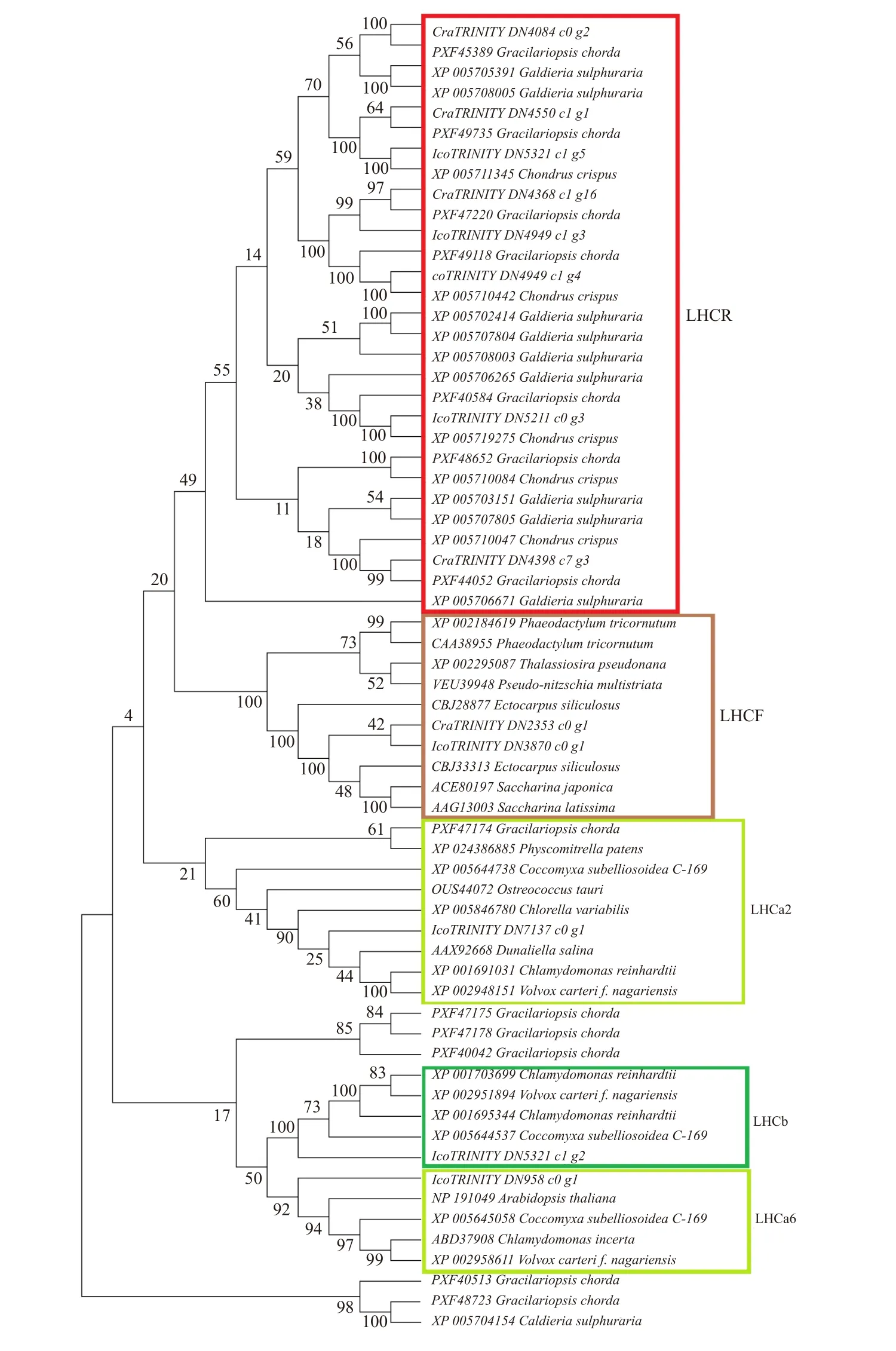
圖4 捕光復合物的系統進化樹Fig. 4 Phylogenetic tree of the light harvesting complex superfamily

圖5 Iridaea cordata的Lhca2、Lhca6和Lhcb基因編碼的蛋白序列與綠藻LHC序列的比較Fig. 5 Alignment of Lhca2, Lhca6 and Lhcb of Iridaea cordata with homologous LHCs in green algae
南極歷史上的冰川期對潮間帶物種造成了毀滅性的破壞,但是南極大陸架邊緣仍有零散的冰川未影響到的區域,成為潮間帶海藻的庇護所。在過去的500萬年間,南極地區至少發生了38個冰期?間冰期循環[42]。由此可能會導致南極潮間帶生物生殖隔離和遺傳分化的產生[43]。利用單一基因的DNA條形碼序列研究冰川期棲息地減少對南極半島和南設得蘭群島常見的6種大型紅藻遺傳多樣性的影響,由于得到的遺傳多樣性水平極低,無法有效劃分海藻的種群,難以確定南極海藻起源的冰川庇護所的位置[6]。采用SSR標記獲得海藻種內的更精細的遺傳學分化,則有希望解決這一問題。同時SSR標記還可以應用于研究南極的地質演變以及預測氣候變化對南極生態系統的影響。
4 結論
南極紅藻是南極地區重要的生物資源,作為極端環境生物,對其轉錄組序列的研究有助于新基因和代謝途徑的發現。由于實驗材料獲取和運輸保藏的困難,很難獲取新鮮的材料提取RNA用于轉錄組測序,這可能是目前南極大型海藻轉錄組學研究較少的原因。我們依托“向陽紅01”號的船載超低溫冰箱,保證了從樣品采集到實驗室RNA提取的過程中,材料保存完好。提取到了符合建庫要求的RNA,順利完成了兩種產膠紅藻的轉錄組測序工作。通過分析轉錄組序列,我們發現了一些可能與南極紅藻光限制和紫外線輻射等極端光環境適應相關的特有基因,南極紅藻更加多樣化的Lhc基因可能是增強I. cordata在光限制環境中的光吸收和能量轉移能力,適應南極冬季弱光環境生存的關鍵。而光裂解酶基因與其適應極地的強紫外線輻射密切相關。這些基因的功能仍有待于進一步的驗證。通過轉錄組測序獲得的SSR標記也為南極紅藻的適應性和遺傳多樣性的研究開展奠定了基礎。
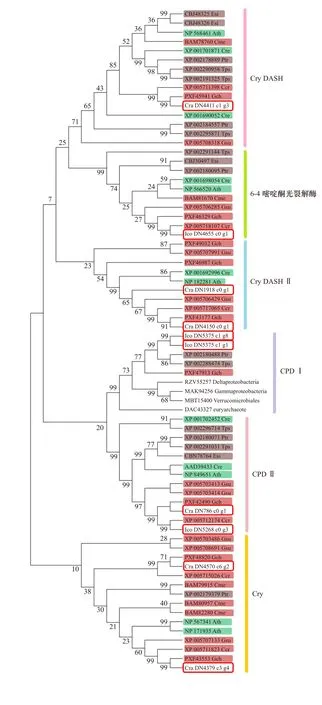
圖6 光裂解酶/隱花色素超家族的系統進化樹Fig. 6 Phylogenetic tree of the cryptochrome/photolyase family
