硫酸亞鐵浸種對黃山欒樹種子發芽及發芽特性的影響研究
2020-10-30 01:54:02鄭清周一濤周瑞明鄧麗娜姜文
安徽農學通報 2020年16期
關鍵詞:植物
鄭清 周一濤 周瑞明 鄧麗娜 姜文


摘要:以硫酸亞鐵(FeS04)為外源浸種劑,研究0.1%、0.2%、0.3%FeS04對黃山欒樹種子浸泡后發芽效果的影響。結果表明:FeS04作為外源浸種劑,改變了黃山欒樹種子的SOD、POD,可溶性蛋白含量和水分含量變化不大,其中以0.1 % FeS〇4對黃山欒樹發芽效果較好。
關鍵詞:黃山欒樹;種子;催芽;硫酸亞鐵;超氧化物歧化酶(SOD);過氧化物酶(POD)
中圖分類號 S792.99 文獻標識碼 A 文章編號 1007-7731(2020)16-0088-03
黃山欒樹(Koelreuteria bipinnata)屬無患子科(Sapin-claceae)欒樹屬(Koelreuteria)木本植物,樹形挺拔,美麗端莊,枝葉秀麗茂密,春天葉子呈紫紅色,夏天開花樹葉金黃,到秋天之后鮮紅的果子閃爍在葉間十分美麗。對生境土壤的要求不高,在微酸性、中性、鹽堿土均能生長,在江淮以南地區廣泛種植,是行道樹及庭院綠化的常用樹種[1],市場需求量大[2]。播種是黃山欒樹常用的育苗方法,但黃山欒樹種子的發芽率非常低,在沒有人為干預的情況下,黃山欒樹的發芽率僅僅只有10%~?20%[3]。
目前,對黃山欒樹研究多集中在生物學特性方面,而關于外源浸種劑對黃山欒樹種子萌發影響的研究則較少。為此,本研究開展了不同用量FeS04對黃山欒樹種子的影響試驗,測定種子的發芽率,發芽指數以及幼苗的超氧化物歧化酶(SOD)、過氧化物酶(POD)、水分含量等相關指標,探討外源浸種劑及其濃度與黃山欒樹種子萌發的關系,為提高黃山欒樹種子的發芽率以及完善黃山欒樹的育苗技術提供理論依據。
1 材料與方法
1.1 供試材料 種子采集于鹽城市區綠化用黃山欒樹,果實飽滿、整齊一致、無病蟲害。育苗土壤為水稻土。試驗在鹽城工學院植物資源開發實驗室進行。
1.2 試驗方法 將FeS04分別配置成質量分數為0.1%、0.2%、0.3%,并配置好的溶液放入水浴鍋中水浴加熱至45℃。將一定量的種子分別放入配置好的溶液中催芽12h,再將種子清洗干凈后擦干備用。將經過浸種劑處理的黃山欒樹種子在培養皿中發芽,每個培養皿中放入30顆種子,種子之間有間隔,再覆蓋上砂石和土,每個濃度2個重復組。最后把培養皿都放入光照培養箱中,溫度調至24℃,24h光照培養,并且每天澆水,每天記錄發芽的數量至15d,最后計算發芽率。15d時,將各個培養皿內已經發芽的幼苗取出,用直尺測量每株黃山欒樹幼苗的長度,對照試驗組采用只加清水,每個濃度重復2次,取平均值。
1.3 測定指標 (1)形態指標:發芽率、發芽指數:
發芽率(%)=發芽種子數/發芽種子總數×100[4];
發芽指數=∑(第t天發芽數/發芽試驗的天數)[4]
生長指標的測定:在出苗后15d時用測量幼芽的根莖葉長度[4]。株高用直尺,根莖用游標卡尺測量。
(2)生理生化指標:
可溶性蛋白質含量:采用考馬斯亮藍G250法[5];
超氧化歧化酶(SOD)酶活:NBT核黃素比色法[6],總SOD活力(U/g) = (對照組0D值-測定0D值)/對照組0D值×2×反應體系的稀釋倍數/待測樣本的蛋白濃度(mgprot/mL);
過氧化物酶(POD)酶活:愈創木酚比色法[7],POD活力(U/mg)=(測定0D-對照0D值)/12×反應液總體積(mL)/樣本量/反應時間(30min)/勻漿蛋白濃度(mgprot/mL) ×1000含水量:烘干法[8]。
1.4 數據處理 數據處理分析采用Excel 2017軟件處理完成。
2 結果與分析
2.1 不同濃度FeS〇4處理對黃山欒樹發芽率的影響 發芽率是研究植物生長狀況等指標十分重要的因素之一[9]。由圖1可知,FeS04處理后對黃山欒樹種子發芽有一定的促進作用,0.1%FeSO4處理對黃山欒樹發芽率提高的效果最大,15d的發芽率達44%,0.2%FeSO4處理,15d的發芽率達34%,硫酸亞鐵12h浸泡后發芽效果由高到低為0.1% FeS04〉0.3% FeS04〉0.2% FeS04。
2.2 不同濃度FeS04處理對黃山欒樹的發芽指數的影響 發芽指數體現了種子的萌發活性,發芽指數越高說明種子活性越高。由圖2可知,0.1%FeS04處理下發芽指數最高,其次是0.3%,最低的是0.2%處理。
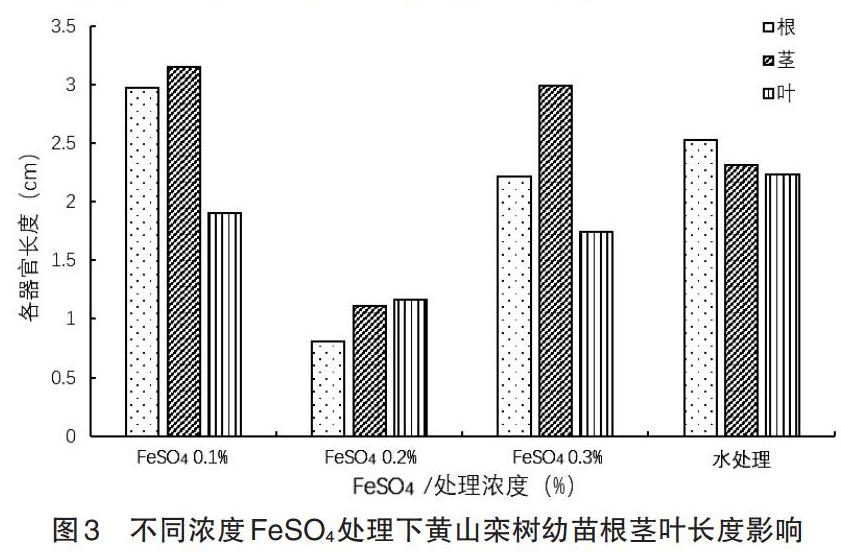
2.3 不同濃度FeS04處理對黃山欒樹幼苗根莖葉長度的影響 幼芽的根莖葉長度是非常重要的指標,可在一定程度上直觀地表現出浸種劑對黃山欒樹中子的催芽效果。在接種后15d時,測量幼芽的根莖葉長度,結果如圖3所示[10]。由圖3可知,各個濃度的處理之中,0.1% FeS04處理的黃山欒樹種子萌發的幼苗平均根長最長,但葉的長度不如對照組,0.2%FeS〇4和0.3%FeS04處理黃山欒樹種子萌發的幼苗根,葉都不如對照組,0.2%FeS04的莖不如對照組,說明雖然0.2%濃度的FeS04處理能夠促進黃山欒樹種子的萌發,但是萌發以后,幼苗的生長較慢,長勢較差,0.3% FeS04的莖比對照組的長度長。
2.4 不同濃度FeS04處理條件對黃山欒樹可溶性蛋白的影響 植物可溶性蛋白的作用十分巨大,他們中的很大一部分都調控著植物的生長與代謝,對于研究黃山欒樹的總代謝指標來說是一個不能忽視的指標[11]。可溶性蛋白含量的提高會使植物種的水分含量增高,因為它能夠使黃山欒樹植物細胞對水分的吸收提高,是一種對植物具有重要意義的滲透調節蛋白。可溶性蛋白的含量作為植物體內最為重要的生理生化指標之一。由圖4可知,在各個濃度的處理之中,0.2% FeS04處理的黃山欒樹種子的可溶性蛋白含量最高。但是總體來看,每一個處理的可溶性蛋白含量都差不多,推測是這幾種方法處理都沒有很有效地提高黃山欒樹種子的可溶性蛋白。
2.5 不同濃度FeS04處理對黃山欒樹S OD酶活的影響SOD(Superoxide dismutase)是超氧化物歧化酶的簡稱,它可以催化超氧陰離子,使其中的自由基被分解成過氧化氫和氧氣,是存在于生物體內細胞十分重要的一種抗氧化金屬酶,它使機體的氧化與抗氧化達成一個平衡,其含量相對穩定,也是植物抗氧化的評價指標之一[12]。它在需氧生物中含量豐富,是第一個在活性氧清除系統中起作用的抗氧化酶[13]。此外,植物中細胞的SOD與很多東西都密切相關,他讓植物能夠抵抗大氣的污染,在冬天氣溫降低的時候讓植物能夠不被凍死,還有干旱時不至于枯萎,以及對于強光照射和鹽度過高滲透壓較高的地方時能夠具有抗性[14]。在植物體內超氧化物歧化酶的活性高時,植物的抗逆性較強,酚代謝也會更加旺盛,并且超氧化物歧化酶的活性也顯示了植物在進化的階段過程中的表現。更重要的是它可以提高植物對于特定環境比如過旱過潮的抵抗能力[15]。SOD酶的活性越高,說明種子的抗逆性越強,越利于黃山欒樹種子的儲存,0.1% FeS04處理的黃山欒樹種子的SOD濃度最低,說明0.1% FeS04處理的黃山欒樹種子萌發最好。
2.6 不同濃度FeS04處理對黃山欒樹POD酶活的影響POD(Peroxidase)是過氧化物酶的英文縮寫,是一種十分重要的氧化還原酶類,是細胞內消除活性氧的保護酶,也是植物進行光合作用的一種十分重要的酶類,能夠消除酚類和過氧化氫、醛和胺類以及苯毒性雙重作用[16],在黃山欒樹中,POD的含量也比較高,它與植物生理活動中的高耗能過程密切相關,比如植物的光合作用以及呼吸作用,隨著植物的不斷生長,POD的含量也會不斷提高,是一種與組織老化密切相關的生理指標[17]。由圖6可知,0.2% FeS04處理的黃山欒樹的POD的酶活最高,其次是0.3% FeS04處理的黃山欒樹種子。
2.7 不同濃度FeS04浸種劑處理對黃山欒樹的含水量的影響 植物組織中的水通常以2種形式存在,一種是結合水,一種是自由水,自由水與結合水的比值的大小對植物組織的影響很大。當自由水和結合水的比值較高時,說明植物的生長較為旺盛,生命代謝活動較快,說明植物長勢較好。而自由水和結合水的比值較低時,說明植物的生長較為緩慢,生命代謝緩慢,但這種情況下,植物的抗逆性會大幅提升。所以研究植物體內水分含量就可以了解植物的生長以及代謝情況,是一種十分重要的指標。由圖7可知,浸種劑對黃山欒樹種子的含水量并無太大的影響,0.1%FeS04處理的黃山欒樹種子的含水量最高。
3 結論
(1)總體上看,在不同濃度FeS04處理中,0.1%濃度的FeS04處理黃山欒樹種子發芽率是最高的,可能是因為0.1%FeS04相較于其他濃度的FeS04更能夠提高種子萌發的蛋白質含量。
(2)FeS04處理的黃山欒樹種子可降低SOD含量,可能是由于FeS04處理降低了黃山欒樹種子的抗逆性,提高了呼吸作用,從而提高了黃山欒樹種子的發芽率。
綜合以上發芽和生理生化指標看出,FeS04可作為外源浸種劑改變黃山欒樹種子的SOD、P0D酶活,可溶性蛋白含量和水分含量變化不大,對黃山欒樹種子具有促進發芽的作用,其中以0.1 %FeS04的催芽作用較好。
參考文獻
[1] 賈玉林.苗圃薦優,優秀的季相樹種,欒樹[J].園林,2007(2):41-42.
[2] 呂潤生,呂敏生,郭松飛,等.欒樹的繁殖與苗木培育[J].中國果菜,2010,23(1):23-24.
[3] 郭紅娜.欒樹的培育與應用[J].花木盆景(花卉園藝),2000(12):24-25.
[4] 金山紅.國槐大樹移植及管理[J].中國林業,2000( 19) :55-55.
[5] 袁良玉,江莉,周揚,等.牛血清白蛋內殘留量檢測試劑盒內部參考品的制備及標定[J].國際生物制品學雜志,2019,42(4):189-l92.
[6] 瞿濤.栽培基質及肥料對盆栽參生長影響的研究[D].吉林:吉林農業大學,2017:33-34.
[7] 裴斌,張光燦,張淑勇,等.土壤干旱脅迫對沙棘葉片光合作用和抗氧化酶活性的影響[J].生態學報,2013,33 (5): 1386-1396.
[8] 項恒.熱風循環隧道烘箱的三段結構和原理[J].科技視界,2017(020:210-211.
[9] 楊淑琴,周瑞蓮,梁慧敏,等.沙漠植物抗氧化酶活性及滲透調節物質含量與光合作用的關系[J].中國沙漠,2015(6):1557-1564.
[10] 俞揚鳳,王鳳珍,張麗林.藥劑浸種提高水稻種子發芽率的試驗[J].作物雜志,2000(4) :30.
[11] 林阿妹.不同催芽方法對黃山欒樹發芽率的影響[J].北京農業,2013(27):52.
[12] Wafa'a A.Al-Taisan, Abclulaziz A.Al-Qarawi, Moodi S.Alsuhiee.Effect of water stress by Polyethylene Glycol 8000 and SodiumChloride on germination of Ephedra alata Decne seeds [J].SaudiJournal of Biological Sciences, 17(3) :253-257.
[13] 朱秀敏.超氧化物歧化酶的生理活性[J].當代醫學,2011,17(15):26-27.
[14] 王峰.水稻鐵超氧化物歧化酶基因及其啟動子的克隆和功能研究[D].中山:中山大學,2005.
[15] 劉西平,胥耀平,王姝清,等.低溫脅迫下欒樹幼苗衰老與膜脂質過氧化關系[J].西北林學院學報,1995(4) :22-23.
[16] 韓鳳,李霞,李海峰,等.過氧化物酶的應用研究進展[J].天然產物研究與開發,2013 (12): 223-228.
[17] 楊淑琴,周瑞蓮,梁慧敏,等.沙漠植物抗氧化酶活性及滲透調節物質含量與光合作用的關系[J].中閏沙漠,2015(6): 1557-1564.
(責編:張宏民)
基金項目:江蘇蘇北科技發展計劃一科技項目SZ-YC2018023。
作者簡介:鄭清(1971-),女,江蘇蘇州人,研究方向:植物資源利用。 收稿日期:2020-07-01
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
