基于微衛(wèi)星分子標(biāo)記的凡納濱對(duì)蝦商業(yè)苗種遺傳多樣性分析*
2020-09-27 06:40:32方振朋孟憲紅李旭鵬曹家旺陳寶龍閆茂倉胡利華
漁業(yè)科學(xué)進(jìn)展 2020年5期
關(guān)鍵詞:分析
方振朋 孟憲紅 李旭鵬 欒 生 曹家旺 陳寶龍 孔 杰 閆茂倉 胡利華
基于微衛(wèi)星分子標(biāo)記的凡納濱對(duì)蝦商業(yè)苗種遺傳多樣性分析*
方振朋1,2孟憲紅2①李旭鵬2欒 生2曹家旺2陳寶龍2孔 杰2閆茂倉3胡利華3
(1. 上海海洋大學(xué)水產(chǎn)與生命學(xué)院 上海 201306;2. 中國水產(chǎn)科學(xué)研究院黃海水產(chǎn)研究所 農(nóng)業(yè)農(nóng)村部海洋漁業(yè)可持續(xù)發(fā)展重點(diǎn)實(shí)驗(yàn)室 青島海洋科學(xué)與技術(shù)試點(diǎn)國家實(shí)驗(yàn)室海洋漁業(yè)科學(xué)與食物產(chǎn)出過程功能實(shí)驗(yàn)室 青島 266071; 3. 浙江省近岸水域生物資源開發(fā)與保護(hù)重點(diǎn)實(shí)驗(yàn)室 溫州 325005)
為研究我國凡納濱對(duì)蝦()商業(yè)苗種的遺傳多樣性特征,于河北、山東、廣東、海南等地采集了6個(gè)有代表性的凡納濱對(duì)蝦品牌苗種,分別命名為黃驊R、東營M、廣州P、廣州Z、海南S和海南Z,以8個(gè)微衛(wèi)星標(biāo)記檢測(cè)其遺傳多樣性。結(jié)果顯示,6個(gè)品牌的凡納濱對(duì)蝦在8個(gè)位點(diǎn)呈現(xiàn)不同程度的多態(tài)性,其平均等位基因數(shù)(a)、期望雜合度(e)、觀測(cè)雜合度(o)和多態(tài)信息含量(PIC)分別為4.5~9.5、0.516~0.733、0.346~0.550和0.472~0.700,各品牌遺傳多樣性豐富程度從高到低分別為黃驊R>廣州Z>廣州P>海南Z>東營M>海南S。哈迪–溫伯格平衡(HWE)檢驗(yàn)顯示,4.17% (2/48)的檢測(cè)結(jié)果表現(xiàn)為顯著的偏離(0.01<<0.05),58.33% (28/48)表現(xiàn)為極顯著偏離(<0.01)。分子變異方差分析(AMOVA)發(fā)現(xiàn),12%的變異來自品牌間,24%的變異來自品牌內(nèi)個(gè)體間,其余64%的變異均來自所有品牌個(gè)體。UPGMA聚類分析結(jié)果顯示,6個(gè)品牌的凡納濱對(duì)蝦聚為2個(gè)明顯的分支,廣州P和廣州Z聚為一支,東營M、黃驊R、海南Z和海南S聚為一支。主成分分析(PCA)結(jié)果顯示,各品牌凡納濱對(duì)蝦無法單獨(dú)進(jìn)行聚類。本研究初步分析了當(dāng)前國內(nèi)養(yǎng)殖凡納濱對(duì)蝦的遺傳背景,實(shí)驗(yàn)結(jié)果可為凡納濱對(duì)蝦良種選育提供數(shù)據(jù)支撐。
凡納濱對(duì)蝦;商業(yè)苗種;微衛(wèi)星;遺傳多樣性
凡納濱對(duì)蝦()原產(chǎn)地主要為中、南美的秘魯北部至墨西哥太平洋沿岸,尤其以厄瓜多爾沿岸分布最為密集(王興強(qiáng)等, 2004)。凡納濱對(duì)蝦為世界三大養(yǎng)殖對(duì)蝦之一(張偉權(quán), 1990),其產(chǎn)量約占全球?qū)ξr產(chǎn)量的70%(張龍等, 2019),具有成活率高、食性廣、生長(zhǎng)快、適應(yīng)能力強(qiáng)等優(yōu)點(diǎn)。凡納濱對(duì)蝦于1988年從美國引入,之后迅速在全國范圍內(nèi)普及(于洋, 2014; 唐揚(yáng)等, 2018),成為我國海水養(yǎng)殖動(dòng)物中養(yǎng)殖發(fā)展最快的一個(gè)種類(馬春艷等, 2011)。目前,我國已成為世界上凡納濱對(duì)蝦養(yǎng)殖產(chǎn)量最高的國家(頡曉勇等, 2008; 童馨等, 2009)。
隨著我國凡納濱對(duì)蝦市場(chǎng)需求量的增大,對(duì)蝦苗種培育方式也出現(xiàn)了轉(zhuǎn)變。目前,在集約化苗種培育技術(shù)已達(dá)峰值的背景下,有些育苗場(chǎng)為了進(jìn)一步降低成本,種蝦不經(jīng)選育,長(zhǎng)期近交繁殖,隨著養(yǎng)殖產(chǎn)量的增大,隨之而來的問題也越來越多。最大的問題莫過于養(yǎng)殖對(duì)蝦遺傳多樣性下降,加之外來親本得不到更新,國內(nèi)養(yǎng)殖對(duì)蝦出現(xiàn)了種質(zhì)退化、病害暴發(fā)頻繁的現(xiàn)象。從長(zhǎng)遠(yuǎn)角度來看,查清我國凡納濱對(duì)蝦遺傳背景,對(duì)進(jìn)一步培育優(yōu)良品種和促進(jìn)可持續(xù)健康養(yǎng)殖具有重要指導(dǎo)意義。
分子標(biāo)記目前已廣泛運(yùn)用于水產(chǎn)動(dòng)物育種中(張瓊等, 2011; 孫苗苗等, 2017; 王軍等, 2018),其中,又以微衛(wèi)星(Microsatellite或Simple Sequence Repeats)分子標(biāo)記應(yīng)用最為普遍。微衛(wèi)星標(biāo)記是2~6個(gè)堿基為核心的短串聯(lián)重復(fù)序列,重復(fù)單位的重復(fù)次數(shù)在個(gè)體間呈高度變異性且數(shù)量豐富,分布于生物體整個(gè)基因組中(Tautz, 1989; Schl?tterer, 1992)。微衛(wèi)星標(biāo)記具有多態(tài)性高、重復(fù)性好、操作簡(jiǎn)單、共顯性遺傳等優(yōu)點(diǎn),被廣泛用于對(duì)蝦群體遺傳多樣性分析(Bringmann, 1996; Postlethwait, 1998; 張?zhí)鞎r(shí)等, 2005; 曾地剛等, 2008)。本研究利用8個(gè)擴(kuò)增穩(wěn)定的微衛(wèi)星位點(diǎn),對(duì)國內(nèi)6個(gè)商業(yè)品牌凡納濱對(duì)蝦苗種進(jìn)行遺傳多樣性分析,初步分析當(dāng)前國內(nèi)養(yǎng)殖凡納濱對(duì)蝦的遺傳背景,為凡納濱對(duì)蝦優(yōu)良品種的選育提供基礎(chǔ)數(shù)據(jù)。
1 材料與方法
1.1 實(shí)驗(yàn)材料
選用我國有代表性的6個(gè)品牌凡納濱對(duì)蝦商業(yè)苗種[平均體長(zhǎng)為(49.73±1.53) mm,平均體重為(1.41± 0.11) g],分別命名為黃驊R、海南Z、海南S、廣州Z、廣州P和東營M。每個(gè)品牌取30尾個(gè)體進(jìn)行肌肉組織DNA提取,共計(jì)180尾凡納濱對(duì)蝦個(gè)體。
1.2 實(shí)驗(yàn)方法
采用天根海洋動(dòng)物組織DNA提取試劑盒進(jìn)行肌肉組織DNA提取,利用紫外分光光度計(jì)(BioImaging ystems, UVP)進(jìn)行DNA濃度測(cè)量,并根據(jù)測(cè)量結(jié)果將DNA稀釋至50 ng/μl。將稀釋得到的DNA利用 8個(gè)微衛(wèi)星位點(diǎn)進(jìn)行PCR擴(kuò)增。PCR反應(yīng)體系:總體積為20 μl,其中,模板DNA 1.5 μl,(Vazyme) 2 ×Master Mix (Dye Plus) 10 μl,正向和反向引物 (10 mmol/L)各0.8 μl,滅菌超純水6.9 μl。PCR反應(yīng)程序:95℃預(yù)變性3 min;95℃變性30 s;72℃退火30 s,72℃延伸30 s,共30個(gè)循環(huán);72℃延伸5 min;4℃保存。PCR產(chǎn)物微衛(wèi)星分型于生工生物工程(上海)股份有限公司,利用ABI 3730XL測(cè)序儀完成,所用微衛(wèi)星位點(diǎn)及其引物信息見表1。
1.3 數(shù)據(jù)處理
對(duì)6個(gè)品牌凡納濱對(duì)蝦商業(yè)苗種進(jìn)行哈迪–溫伯格平衡(Hardy-Weinberg equilibrium, HWE)檢測(cè)和遺傳多樣性分析。利用GENEPOP version 3.4軟件進(jìn)行HWE檢驗(yàn),得到精確值(H-W)。通過Cervus 3.0計(jì)算各位點(diǎn)的等位基因數(shù)目(Number of alleles,a),觀測(cè)雜合度(Observed heterozygosities,o)、期望雜合度(Expected heterozygosities,e)、8個(gè)微衛(wèi)星位點(diǎn)的多態(tài)信息含量(Polymorphism information content, PIC)和無效等位基因頻率[Null alleles frequency,(Null)]。利用GenAlEx 6.51軟件計(jì)算6個(gè)品牌凡納濱對(duì)蝦在各位點(diǎn)的香農(nóng)多樣性指數(shù)(Shannon's diversity index)、8個(gè)位點(diǎn)每個(gè)品牌特有等位基因(Private alleles)平均值、各品牌間的遺傳分化程度和基因流(包括總的ST值以及兩兩品牌間的ST值);分子變異方差分析(Analysis of molecular variance, AMOVA),估測(cè)遺傳變異在品牌內(nèi)和品牌間的分配情況;6個(gè)品牌間Nei’s無偏遺傳距離、遺傳相似性系數(shù)、品牌間遺傳距離矩陣、個(gè)體間遺傳距離矩陣;利用品牌間和個(gè)體間遺傳距離矩陣進(jìn)行主成分分析(Principal coordinate analysis, PCA)。利用NTSYSpc 2.1軟件按照遺傳一致度進(jìn)行6個(gè)品牌凡納濱對(duì)蝦UPGMA聚類。
2 結(jié)果
2.1 6個(gè)商業(yè)品牌凡納濱對(duì)蝦苗種遺傳多樣性分析
6個(gè)品牌凡納濱對(duì)蝦商業(yè)苗種的遺傳多樣性參數(shù)見表2和圖1。在8個(gè)微衛(wèi)星位點(diǎn)中,6個(gè)品牌凡納濱對(duì)蝦總等位基因數(shù)為36~76個(gè),最大為黃驊R,最小為東營M。各品牌平均等位基因數(shù)為4.5~9.5個(gè)。黃驊R平均每個(gè)微衛(wèi)星位點(diǎn)存在1.625個(gè)特有等位基因,在所有品牌中最多。東營M平均每個(gè)位點(diǎn)存在0.125個(gè)特有等位基因,為各品牌中最少。8個(gè)微衛(wèi)星位點(diǎn)的多態(tài)信息含量(PIC)值為0.343~0.925,最高為TuMXLv7.56位點(diǎn),最低為M1103位點(diǎn),除M1103位點(diǎn)外,其他7個(gè)微衛(wèi)星位點(diǎn)PIC都大于0.5。黃驊R、廣州Z和海南Z的香農(nóng)多樣性指數(shù)較高,其他3個(gè)品牌較低。8個(gè)微衛(wèi)星位點(diǎn)的無效等位基因頻率范圍為0.031~0.403,5個(gè)位點(diǎn)存在無效等位基因。8個(gè)微衛(wèi)星位點(diǎn)分別對(duì)6個(gè)品牌凡納濱對(duì)蝦進(jìn)行了48次HWE檢驗(yàn)。其中,4.17%(2/48)的檢測(cè)結(jié)果表現(xiàn)為顯著的偏離(0.01<<0.05),58.33%(28/48)表現(xiàn)為極顯著偏離(<0.01),其余的37.5%(18/48)符合HWE (>0.05)。
表1 8個(gè)微衛(wèi)星位點(diǎn)信息

Tab.1 The information of 8 microsatellites loci
2.2 6個(gè)商業(yè)品牌凡納濱對(duì)蝦的遺傳分化
分子變異方差分析(AMOVA)發(fā)現(xiàn),僅有12%的遺傳變異來自品牌間,24%來自品牌內(nèi)個(gè)體間,其余64%的變異均來自所有品牌個(gè)體,這表明遺傳變異主要存在于個(gè)體間(表3)。
6個(gè)品牌凡納濱對(duì)蝦商業(yè)苗種的統(tǒng)計(jì)量分別為0.359 (IT),0.273 (IS),0.118 (ST)。兩兩品牌間的值ST在0.034和0.111之間,均<0.15(表4),且明顯的分成兩部分。其中,小于0.05的為4組,占全部15組的26.67%,沒有出現(xiàn)遺傳分化(ST<0.05);介于0.05和0.15之間的為11組,占全部15組的73.33%,為中等程度分化(0.05
2.3 6個(gè)商業(yè)品牌凡納濱對(duì)蝦的遺傳聚類
從表5可以看出,東營M和海南Z品牌間的遺傳距離最近(0.128),遺傳相似性系數(shù)最大(0.880),親緣關(guān)系最近;廣州P和海南S品牌間的遺傳距離最遠(yuǎn)(0.549),遺傳相似性系數(shù)最小(0.578),親緣關(guān)系最遠(yuǎn)。由圖2可以看出,6個(gè)品牌的凡納濱對(duì)蝦聚為2個(gè)明顯的分支,廣州P和廣州Z聚為一支,東營M、黃驊R、海南Z和海南S聚為一支。
2.4 6個(gè)商業(yè)品牌凡納濱對(duì)蝦遺傳關(guān)系主成分分析
主成分分析(PCA)結(jié)果顯示(圖3),6個(gè)品牌凡納濱對(duì)蝦商業(yè)苗種產(chǎn)生了明顯的遺傳歧化。其中,廣州P和廣州Z最為聚集,其次為東營M和海南Z,之后為黃驊R和海南S,主成分1 (Coord.1)和主成分2 (Coord.2)共解釋了總遺傳變異的71.22%。在個(gè)體水平上,主成分1 (Coord.1)和主成分2 (Coord.2)共解釋了總遺傳變異的20.05%。從圖3可知,6個(gè)品牌的180尾凡納濱對(duì)蝦個(gè)體聚集較為集中,其中,廣州P、廣州Z和黃驊R 3個(gè)品牌的凡納濱對(duì)蝦交集最大。同時(shí),每個(gè)品牌的凡納濱對(duì)蝦個(gè)體無法單獨(dú)聚為一類。
表2 6個(gè)品牌凡納濱對(duì)蝦在8個(gè)微衛(wèi)星位點(diǎn)中的遺傳多樣性參數(shù)

Tab.2 Genetic diversity indices of 8 microsatellite loci of L. vannamei from 6 brands
3 討論
微衛(wèi)星分子標(biāo)記在種內(nèi)有高度的遺傳變異,是群體遺傳分化分析的有效標(biāo)記(孫效文等, 2008)。微衛(wèi)星分子標(biāo)記廣泛分布于基因組中,數(shù)量眾多,容易檢測(cè),在分子輔助育種和物種多樣性檢測(cè)等方面被廣泛應(yīng)用(張麗娟等, 2014)。群體的遺傳多樣性來源于物種適應(yīng)復(fù)雜環(huán)境和生存進(jìn)化(Li, 2016),其主要表現(xiàn)在等位基因數(shù)的豐富和均勻程度、遺傳雜合度的大小、多態(tài)信息含量的高低3個(gè)方面(Beardmore, 1997; 王鶴等, 2016)。研究發(fā)現(xiàn),等位基因越豐富,遺傳雜合度數(shù)值越大,多態(tài)信息含量越高,則群體遺傳變異更高,更能應(yīng)對(duì)環(huán)境變化,也更能產(chǎn)生優(yōu)質(zhì)的種質(zhì)資源(Eschenroeder, 2016)。

圖1 6個(gè)品牌凡納濱對(duì)蝦商業(yè)苗種遺傳多樣性參數(shù)均值(8個(gè)微衛(wèi)星位點(diǎn))
表3 6個(gè)品牌凡納濱對(duì)蝦分子變異方差分析
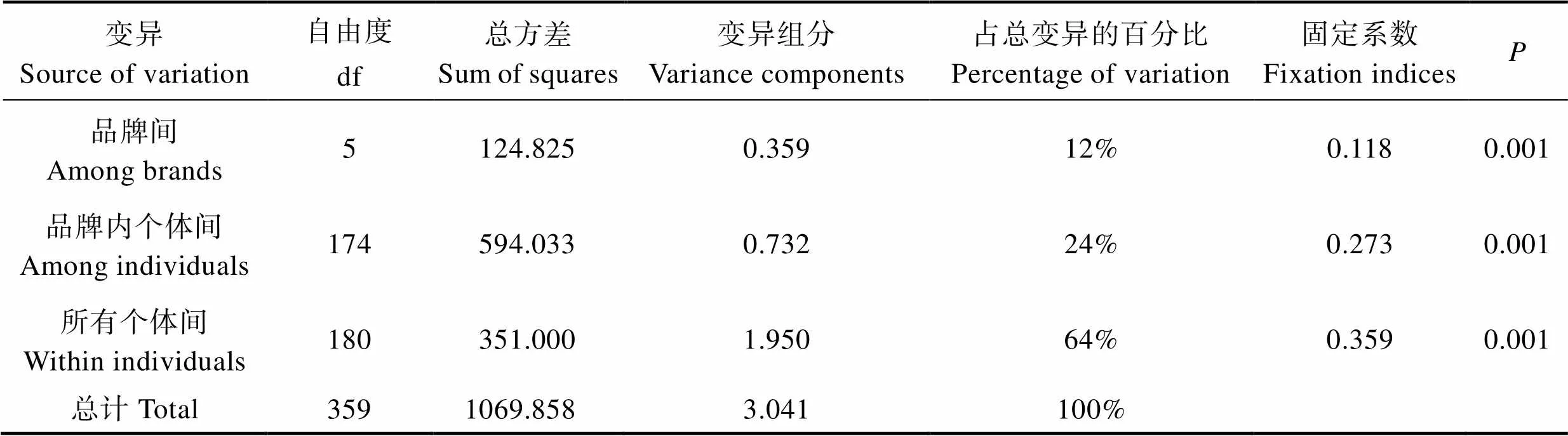
Tab.3 Analysis of molecular variances (AMOVA) of microsatellites in 6 brands of L. vannamei
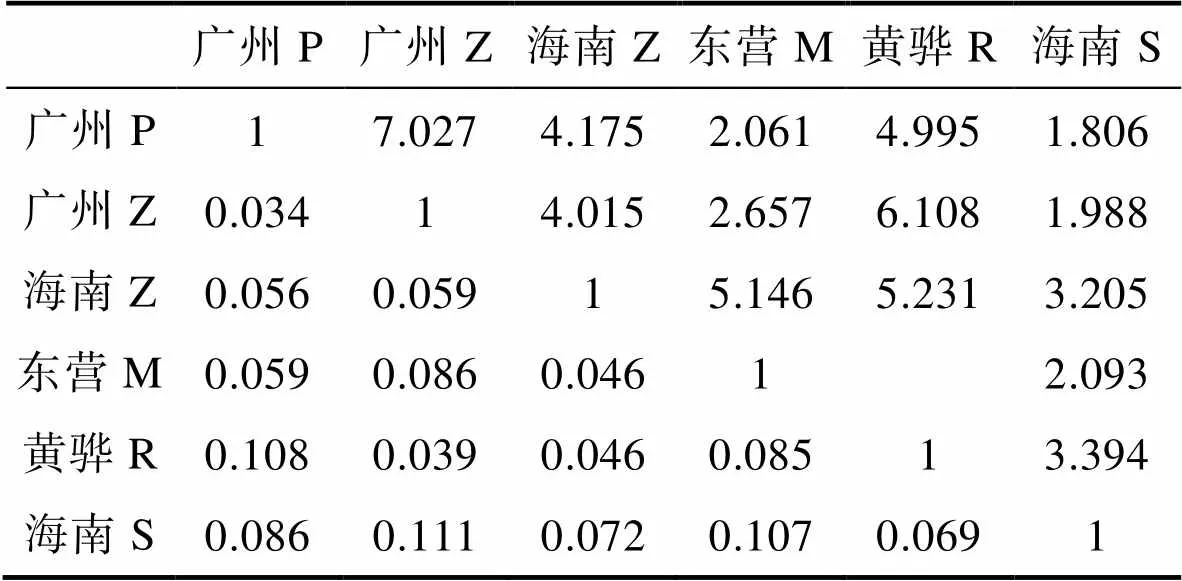
表4 6個(gè)品牌凡納濱對(duì)蝦FST值(下三角)與Nm值(上三角)
表5 6個(gè)品牌凡納濱對(duì)蝦遺傳相似性系數(shù)(下三角)與遺傳距離(上三角)

Tab.5 Genetic similarity coefficient (below the diagonal) and genetic distance (above the diagonal) of six brands’ L. vannamei

圖2 根據(jù)遺傳一致度構(gòu)建的UPGMA樹
關(guān)于凡納濱對(duì)蝦微衛(wèi)星標(biāo)記開發(fā)和篩選的工作已有大量報(bào)道(Meehan,, 2003; 馬寧等, 2013; 楊銘等, 2017)。本研究從實(shí)驗(yàn)室發(fā)表過的文章(李東宇等, 2016)中篩選出8個(gè)擴(kuò)增穩(wěn)定的微衛(wèi)星位點(diǎn),分析了6個(gè)國內(nèi)品牌的凡納濱對(duì)蝦商業(yè)苗種的遺傳多樣性、遺傳結(jié)構(gòu)和親緣關(guān)系。結(jié)果顯示,87.5% (7/8)的位點(diǎn)平均多態(tài)信息含量(PIC)都在0.5以上。根據(jù)Botstein等(1980)提出的衡量標(biāo)準(zhǔn),當(dāng)PIC>0.5時(shí),意味著該位點(diǎn)為高度多態(tài)位點(diǎn),這也說明本研究所引用的微衛(wèi)星標(biāo)記的多態(tài)性較高。基因雜合度是衡量群體遺傳變異水平的理想?yún)?shù)(王日芳等, 2017),8個(gè)微衛(wèi)星位點(diǎn)中的觀測(cè)雜合度(o)平均為0.486,期望雜合度(e)平均為0.745,觀測(cè)雜合度小于期望雜合度,也從一個(gè)側(cè)面說明了國內(nèi)商業(yè)苗種凡納濱對(duì)蝦出現(xiàn)了雜合子缺失、純合子過剩的情況。在進(jìn)行微衛(wèi)星分型的8個(gè)位點(diǎn)中,有5個(gè)(62.5%)位點(diǎn)顯著偏離HWE (<0.05)。產(chǎn)生該結(jié)果的原因:一方面為實(shí)驗(yàn)對(duì)蝦純合子過剩、雜合子缺失;另一方面為水產(chǎn)動(dòng)物生物結(jié)構(gòu)較為簡(jiǎn)單,從而導(dǎo)致的遺傳變異度也較高。因此,造成微衛(wèi)星位點(diǎn)在同一物種不同個(gè)體中發(fā)生堿基形態(tài)和數(shù)目的變異,無法擴(kuò)增正確的等位基因,擴(kuò)增的無效等位基因會(huì)導(dǎo)致分型結(jié)果顯著偏離HWE (舒妙安等, 2011; 宋忠魁等, 2013)。親緣關(guān)系的遠(yuǎn)近可以用遺傳距離來反映(陳子桂等, 2016; 劉洪濤等, 2018)。在本研究中,遺傳距離最大的2個(gè)品牌是廣州P和海南S,遺傳距離最小的2個(gè)品牌是東營M和海南Z,并且針對(duì)個(gè)體的遺傳關(guān)系主成分分析(PCA)表明,6個(gè)品牌的凡納濱對(duì)蝦無法依照每個(gè)品牌單獨(dú)聚集,品牌間親緣關(guān)系較為接近。這種結(jié)果表明,當(dāng)前國內(nèi)不同品牌的凡納濱對(duì)蝦來源存在一定的相似性,也可能是二代、三代苗種導(dǎo)致部分品牌對(duì)蝦出現(xiàn)近交情況,使其親緣關(guān)系逐漸接近。

圖3 6個(gè)品牌凡納濱對(duì)蝦主成分分析
種苗處于對(duì)蝦產(chǎn)業(yè)鏈上游,其質(zhì)量對(duì)養(yǎng)殖成敗起關(guān)鍵作用(黃小帥等, 2019)。本研究所分析的6個(gè)品牌凡納濱對(duì)蝦商業(yè)苗種中,黃驊R與廣州Z具有較高的遺傳多樣性,不同品牌的商業(yè)苗種間的遺傳特征存在一定差異。在當(dāng)前凡納濱對(duì)蝦養(yǎng)殖規(guī)模不斷擴(kuò)大的情況下,研究遺傳因素與養(yǎng)殖生產(chǎn)性能之間的關(guān)聯(lián)尤為重要,只有充分掌握對(duì)蝦群體的遺傳背景,才能合理利用雜交優(yōu)勢(shì)進(jìn)行良種選育。
Beardmore JA, Mair GC, Lewis RI. Biodiversity in aquatic systems in relation to aquaculture. Aquaculture Research, 1997, 28(10): 829–839
Botstein D, White RL, Skolnick M,. Construction of a genetic linkage map in man using restriction fragment lengthpolymorphisms. American Journal of Human Genetics, 1980, 32(3): 314–331
Bringmann P, Rinke J, Appel B,. Microsatellite polymorphism and the population structure of Atlantic cod () in the northwest Atlantic. Canadian Journal of Fisheries and Aquatic Sciences, 1996, 53(4): 2706–2721
Chen ZG, Wang PP, Liao Y,. Genetic diversity analysis of F2generation families of carp hybrid lines population using microsatellite. Fisheries Science and Technology Information, 2016, 43(2): 83–87 [陳子桂, 王培培, 廖愚, 等. 微衛(wèi)星分析鯉魚雜交系F2代家系群體遺傳多樣性. 水產(chǎn)科技情報(bào), 2016, 43(2): 83–87]
Eschenroeder JC, Roberts JH. Novel polymorphic microsatellite loci for distinguishing rock bass (), Roanoke bass (), and their hybrids. Molecular Biology Reports, 2016, 43(10): 1035–1039
Huang XS, Xu Y, Hu XJ,. Genetic diversity analysis of first filial generation of seven introducedpopulations using microsatellite DNA markers. South China Fisheries Science, 2019, 15(1): 54–62 [黃小帥, 徐煜, 胡曉娟, 等. 利用微衛(wèi)星標(biāo)記分析7個(gè)凡納濱對(duì)蝦引進(jìn)群體子一代的遺傳多樣性. 南方水產(chǎn)科學(xué), 2019, 15(1): 54–62]
Jie XY, Su TF, Chen W,. Analysis on genetic diversity of six cultured stocks of. South China Fisheries Science, 2008, 4(6): 42–49 [頡曉勇, 蘇天鳳, 陳文, 等. 凡納濱對(duì)蝦6個(gè)養(yǎng)殖群體遺傳多樣性的比較分析. 南方水產(chǎn), 2008, 4(6): 42–49]
Li DY, Kong J, Meng XH,. Development of multiplex PCR systems of microsatellite markers for Pacific white shrimp () and its application for parentage identification. Progress in Fishery Sciences, 2016, 37(3): 58–67 [李東宇, 孔杰, 孟憲紅, 等. 凡納濱對(duì)蝦()微衛(wèi)星多重PCR體系的建立及其在家系親權(quán)鑒定中的應(yīng)用. 漁業(yè)科學(xué)進(jìn)展, 2016, 37(3): 58–67]
Li Y, Zhao H, Peng Z,. Isolation and characterization of 21 polymorphic microsatellite markers for a new labeonine fish (Zhao. 2014) using illumina paired-end sequencing. Journal of Applied Ichthyology, 2016, 32(1): 126–128
Liu HT, Yang MQ, He YG,. Microsatellite analysis of genetic diversity in eight geographical populations of. Natural Science Journal of Hainan University, 2018, 36(2): 146–152 [劉洪濤, 楊明秋, 何玉貴, 等. 凡納濱對(duì)蝦八個(gè)地理群體遺傳多樣性的微衛(wèi)星分析. 海南大學(xué)學(xué)報(bào)自然科學(xué)版, 2018, 36(2): 146–152]
Ma CY, Ma HY, Ma LB,. Microsatellite analysis on genetic variation of importedpopulanon and culture stocks. Marine Fisheries, 2011, 33(1): 1–8 [馬春艷, 馬洪雨, 馬凌波, 等. 凡納濱對(duì)蝦引進(jìn)群體和2個(gè)養(yǎng)殖群體遺傳變異的微衛(wèi)星分析. 海洋漁業(yè), 2011, 33(1): 1–8]
Ma N, Zeng DG. Isolation of microsatellite sequences from. Southwest China Journal of Agricultural Sciences, 2013, 26(6): 2629–2633 [馬寧, 曾地剛. 凡納濱對(duì)蝦微衛(wèi)星序列的篩選. 西南農(nóng)業(yè)學(xué)報(bào), 2013, 26(6): 2629–2633]
Meehan D, Xu Z, Zuniga G,. High frequency and large number of polymorphic microsatellites in cultured shrimp,()[Crustacea: Decapoda]. Marine Biotechnology, 2003, 5(4): 311–330
Postlethwait JH, Yan YL, Gates MA,. Vertebrate genome evolution and the zebrafish gene map. Nature Genetics, 1998, 18(4): 345–349
Schl?tterer C, Tautz D. Slippage synthesis of simple sequence DNA. Nucleic Acids Research, 1992, 20(2): 211–215
Shu MA, Zhou YF, Zhu XY,. Microsatellite analysis on genetic diversity of seven wild populations ofin China. Journal of Fisheries of China, 2011, 35(7): 977–984 [舒妙安, 周宇芳, 朱曉宇, 等. 中國沿海擬穴青蟹群體遺傳多樣性的微衛(wèi)星分析. 水產(chǎn)學(xué)報(bào), 2011, 35(7): 977–984]
Song ZK, Nie ZP, Wang YF. Screening for polymorphic primer pairs of (CA)microsatellite from(mud crab) genome. Marine Sciences, 2013, 37(10): 98–103 [宋忠魁, 聶振平, 王芳宇. 擬穴青蟹(CA)微衛(wèi)星DNA多態(tài)性引物篩選. 海洋科學(xué), 2013, 37(10): 98–103]
Sun MM, Yang QB, Chen BY,. Genetic diversity in four populations ofassessed by AFLP makers. Marine Sciences, 2017, 41(3): 41–47 [孫苗苗, 楊其彬, 陳百堯, 等. 斑節(jié)對(duì)蝦4個(gè)地理群體遺傳多樣性的AFLP分析. 海洋科學(xué), 2017, 41(3): 41–47]
Sun XW, Zhang XF, Zhao YY,. Development and application of microsatellite markers in aquatic species. Journal of Fishery Sciences of China, 2008, 15(4): 689–703 [孫效文, 張曉鋒, 趙瑩瑩, 等. 水產(chǎn)生物微衛(wèi)星標(biāo)記技術(shù)研究進(jìn)展及其應(yīng)用. 中國水產(chǎn)科學(xué), 2008, 15(4): 689–703]
Tang Y, Meng XF, Shen RF,. Research and application of family selective breeding in culture of Pacific white shrimp. Fisheries Science, 2018, 37(4): 555– 563 [唐揚(yáng), 孟小菲, 沈瑞福, 等. 凡納濱對(duì)蝦家系選育的研究與應(yīng)用. 水產(chǎn)科學(xué), 2018, 37(4): 555–563]
Tautz D. Hypervariabflity of simple sequences as a general source for polymorphic DNA markers. Nucleic Acids Research, 1989, 17(16): 6463–6471
Tong X, Gong SY, Yu DH,. Genetic diversity of cultured Pacific white shrimp () stocks of different generations in China. Oceanologia et Limnologia Sinica, 2009, 40(2): 214–220 [童馨, 龔世圓, 喻達(dá)輝, 等. 凡納濱對(duì)蝦()不同世代養(yǎng)殖群體的遺傳多樣性分析. 海洋與湖沼, 2009, 40(2): 214–220]
Wang H, Wang TT, Hu LP,. Microsatellite analysis of gentic characteristics in crosses between Korea population and China population of sea cucumber. Fisheries Science, 2016, 35(1): 60–66 [王鶴, 王田田, 胡麗萍, 等. 仿刺參中國群體與韓國群體雜交子代微衛(wèi)星標(biāo)記分析. 水產(chǎn)科學(xué), 2016, 35(1): 60–66]
Wang J, Wang QY, Kong J,. SSR analysis on genetic diversity in breeding and wild populations ofProgress in Fishery Sciences, 2018, 39(2): 104– 111 [王軍, 王清印, 孔杰, 等. 中國明對(duì)蝦人工選育群體與野生群體遺傳多樣性的SSR分析. 漁業(yè)科學(xué)進(jìn)展, 2018, 39(2): 104–111]
Wang RF, Li J, Li JT,. Genetic analysis of 33 microsatellite loci in 3 families ofinbred line. Progress in Fishery Sciences, 2017, 38(4): 78–86 [王日芳, 李健, 李吉濤, 等. 脊尾白蝦()近交系3個(gè)家系33個(gè)微衛(wèi)星座位的遺傳分析. 漁業(yè)科學(xué)進(jìn)展, 2017, 38(4): 78–86]
Wang XQ, Ma S, Dong SL. Studies on the biology and cultural ecology of: A review. Transactions of Oceanology and Limnology, 2004(4): 94–100 [王興強(qiáng), 馬甡, 董雙林. 凡納濱對(duì)蝦生物學(xué)及養(yǎng)殖生態(tài)學(xué)研究進(jìn)展. 海洋湖沼通報(bào), 2004(4): 94–100]
Yang M, Yu Y, Zhang XJ,. Development of microsatellite markers from the transcriptome sequences of Pacific white shrimp (). Marine Sciences, 2017, 41(2): 96–102 [楊銘, 于洋, 張曉軍, 等. 基于轉(zhuǎn)錄組數(shù)據(jù)的凡納濱對(duì)蝦微衛(wèi)星標(biāo)記開發(fā). 海洋科學(xué), 2017, 41(2): 96–102]
Yu Y. Development of molecular markers and their applications in selective breeding of the Pacific white shrimp,. Doctoral Dissertation of Institute of Oceanology, Chinese Academy of Sciences, 2014 [于洋. 凡納濱對(duì)蝦分子標(biāo)記的開發(fā)及其在遺傳育種中的應(yīng)用. 中國科學(xué)院研究生院(海洋研究所)博士研究生學(xué)位論文, 2014]
Zeng DG, Chen XH, Li YM,. SSR Analysis of genetic diversity of. Reservoir Fisheries, 2008, 28(1): 31–34 [曾地剛, 陳曉漢, 李詠梅, 等. 凡納濱對(duì)蝦遺傳多樣性的SSR分析. 水利漁業(yè), 2008, 28(1): 31–34]
Zhang L, Chen Z, Wang L,. Effect of stocking density on intermediate cultivation ofpostlarvae. Progress in Fishery Sciences, 2019, 40(1): 76–83 [張龍, 陳釗, 汪魯, 等. 放養(yǎng)密度對(duì)凡納濱對(duì)蝦苗種中間培育效果的影響. 漁業(yè)科學(xué)進(jìn)展, 2019, 40(1): 76–83]
Zhang LJ, Zhu F, Yin SW,. Microsatellite analysis of,and their hybrid progeny. Freshwater Fisheries, 2014, 44(5): 9–14 [張麗娟, 祝斐, 尹紹武, 等. 河川沙塘鱧、鴨綠沙塘鱧及其雜交子代的微衛(wèi)星分析. 淡水漁業(yè), 2014, 44(5): 9–14]
Zhang Q, Li XL, Xue SW,. Screening and polymorphism of EST-microsatellite markers in. Acta Oceanologica Sinica, 2011, 33(1): 121–126 [張瓊, 李喜蓮, 薛淑雯, 等. 凡納濱對(duì)蝦EST-SSR標(biāo)記的篩選及遺傳多態(tài)性檢測(cè). 海洋學(xué)報(bào), 2011, 33(1): 121–126]
Zhang TS, Liu P, Li J,. Genetic diversity of cultured populations ofshrimp using microsatellites. Journal of Fisheries of China, 2005, 29(1): 6–12 [張?zhí)鞎r(shí), 劉萍, 李健, 等. 用微衛(wèi)星DNA技術(shù)對(duì)中國對(duì)蝦人工選育群體遺傳多樣性的研究. 水產(chǎn)學(xué)報(bào), 2005, 29(1): 6–12]
Zhang WQ. An important breeding species in the world?Biology of. Marine Sciences, 1990, 14: 69–73 [張偉權(quán). 世界重要養(yǎng)殖品種——南美白對(duì)蝦生物學(xué)簡(jiǎn)介. 海洋科學(xué), 1990, 14: 69–73]
Genetic Diversity Analysis of Domestic Commercial Brands Seedlings ofBased on Microsatellite Molecular Markers
FANG Zhenpeng1,2, MENG Xianhong2①, LI Xupeng2, LUAN Sheng2, CAO Jiawang2, CHEN Baolong2, KONG Jie2, YAN Maocang3, HU Lihua3
(1.College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306; 2. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, Laboratory of Marine Fisheries Science and Food Production Processes, Pilot National Laboratory of Marine Science and Technology (Qingdao),Qingdao 266071; 3. Zhejiang Key Laboratory of Exploitation and Preservation of Coastal Bio-Resource, Wenzhou 325005)
To clarify the genetic diversity characteristics of domestic commercial brand seedlings of, six domestic commercial brands ofnamely, Huanghua R, Dongying M, Guangzhou P, Guangzhou Z, Hainan S, and Hainan Z, were collected from Hebei, Shandong, Guangdong, and Hainan Province. Their genetic diversity was detected using eight microsatellite markers. The results showed that the average allele number (a), expected heterozygosity (e), observed heterozygosity (o), and polymorphic information content (PIC) were 4.5~9.5, 0.516~0.733, 0.346~0.550, and 0.472~0.700, respectively. The genetic diversity of the six brands ranged from high to low: Huanghua R > Guangzhou Z > Guangzhou P > Hainan Z >Dongying M > Hainan S, and 48 Hardy-Weinberg equilibrium tests were conducted on the 6 brands ofusing 8 microsatellite loci. It was observed that, 4.17% (2/48) showed significant deviation (0.01<<0.05) and 58.33% (28/48) showed extremely significant deviation (<0.01). Molecular variance analysis (AMOVA) showed that 12% of the variance was from the brand, 24% was from the individual within the brand, and the remaining 64% was from all individual brands. UPGMA clustering map showed that each brand ofcould not cluster independently. The analysis showed that the six brands ofclustered into two distinct branches, Guangzhou P and Guangzhou Z clustered into one branch, and Dongying M, Huanghua R, Hainan Z, and Hainan S clustered into another branch. PCA results showed that each brand ofcould not cluster independently. This study preliminarily analyzed the genetic background ofcultured in China. The experimental results can provide data to support the breeding of improved varieties of.
Pacific white shrimp; Commercial seedling; Microsatellite; Genetic diversity
MENG Xianhong, E-mail: mengxianhong@ysfri.ac.cn
S917.4
A
2095-9869(2020)05-0012-09
10.19663/j.issn2095-9869.20190527004
http://www.yykxjz.cn/
方振朋, 孟憲紅,李旭鵬, 欒生, 曹家旺, 陳寶龍, 孔杰, 閆茂倉, 胡利華. 基于微衛(wèi)星分子標(biāo)記的凡納濱對(duì)蝦商業(yè)苗種遺傳多樣性分析. 漁業(yè)科學(xué)進(jìn)展, 2020, 41(5): 101–109
Fang ZP, Meng XH, Li XP, Luan S, Cao JW, Chen BL, Kong J, Yan MC, Hu LH. Genetic diversity analysis of domestic commercial brands seedlings ofbased on microsatellite molecular markers. Progress in Fishery Sciences, 2020, 41(5): 101–109
* 浙江省近岸水域生物資源開發(fā)與保護(hù)重點(diǎn)實(shí)驗(yàn)室開放基金項(xiàng)目(J2018003)、中國水產(chǎn)科學(xué)研究院基本科研業(yè)務(wù)費(fèi)(2018GH10)、山東省農(nóng)業(yè)良種工程(2017LZN011)、泰山學(xué)者良種工程項(xiàng)目和現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系專項(xiàng)資金(CARS-48)共同資助[This work was supported by Open Research Fund from Zhejiang Key Laboratory of Exploitation and Preservation of Coastal Bio-Resource (J2018003), Central Public-Interest Scientific Institution Basal Research Fund, CAFS (2018GH10), Shandong Province Agricultural Seed Improvement Project (2017LZN011), Taishan Scholar Program for Seed Industry, and China Agriculture Research System(CARS-48)]. 方振朋,E-mail: fangzhenpeng@126.com
孟憲紅,研究員,E-mail: mengxianhong@ysfri.ac.cn
2019-05-27,
2019-06-15
(編輯 馬璀艷)
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
