miRNA-100低表達間充質干細胞對急性髓細胞性白血病細胞增殖、凋亡的影響及機制
2020-08-05 05:30:32繆華緯孔凡良蔣英俊王鳳云
山東醫藥 2020年20期
關鍵詞:檢測
繆華緯,孔凡良,蔣英俊,王鳳云
合肥市第二人民醫院,合肥230011
急性髓細胞性白血病(AML)是一種高度惡性的血液性疾病[1,2]。表觀學調控尤其是微小RNA(miRNA)對其靶基因的轉錄調節在白血病發生發展中起著關鍵作用[3,4]。有研究顯示,基因的異常改變可以導致miRNA功能失活,從而導致白血病干細胞增殖、分化[6,7]。miR-100被報道參與AML的發生發展[8],但是其作用機制尚未明確。近年來的研究表明,白血病的發生發展受到微環境的調控,尤其是間充質干細胞(MSC)[9]。2019年1~9月,本研究觀察了MSC中miR-100表達失調對AML細胞惡性生物學行為的影響,現報告如下。
1 材料與方法
1.1 細胞與主要實驗材料 人骨髓MSC(BFN60808567)購買于青旗生物技術發展有限公司。人白血病細胞株K562來源于安徽醫科大學。K562細胞采用完全培養基進行培養。當細胞培養至80%融合度時進行傳代。所有細胞均培養于無菌細胞培養箱中(37 ℃、5% CO2)。DMEM、胎牛血清、青霉素、鏈霉素皆購自美國Gibco公司。miRNA-100的相關引物購于Invitrogen公司。miRNA提取試劑盒購自TaKaRa公司。miR-100-3p inhibitor質粒、慢病毒載體購自上海漢恒公司。6孔細胞培養板購于康寧公司。PI染色試劑盒購于碧云天公司。Western blotting實驗所用一抗購自美國CST公司,二抗購自美國Sigma公司。
1.2 miR-100低表達MSC的構建 將MSC分為兩組,分別感染miR-100-NC和miR-100-inhibitor慢病毒。感染后觀察熒光強度,并通過嘌呤霉素篩選穩定細胞株。待所有細胞均有熒光表達時即可停止篩選,擴大培養獲得穩定表達目的基因的細胞株。采用qRT-PCR法檢測MSC中的miR-100,以GAPDH作為內參。GAPDH上游引物序列為5′-GGAGCGAGATCCCTCCAAAAT-3′,下游引物序列為5′-GGCTGTTGTCATACTTCTCATGG-3′。miR-100上游引物序列為5′-AACCCGUAGAUCCGAACUUGUG3′,下游引物序列為5′-GTCGTATCCAGTGCAGGGTCCGAGGTCACTGGAT-ACGACCACAAGT-3′。
1.3 MSC與K562細胞共培養及分組 兩組MSC培養3 d后,胰酶消化黏附生長的MSC。取1×104個細胞接種于6孔板,待過夜,加入消化好的K562細胞(懸浮生長)。共培養3 d后,吸取培養基(含有K562細胞),離心沉淀獲取K562細胞,與感染miR-100-NC慢病毒MSC共培養的細胞為對照組,與感染miR-100-inhibitor慢病毒MSC共培養的細胞為低表達組。
1.4 K562細胞增殖能力觀察 ①MTT實驗:于共培養3 d后采用MTT法檢測細胞增殖能力。將20 μL的MTT溶液分別加至每個孔并將6孔板置于37 ℃孵箱中孵育2 h。用移液槍吸凈孔內培養基及MTT溶液,加入DMSO溶液,在搖床上避光溶解30 min后,上機檢測OD值表示細胞增殖能力。②克隆形成實驗:共培養3 d后采用平板克隆實驗反映細胞增殖能力。收集細胞,在6孔板中以5×102/孔接種,加入2 mL完全培養基,放于孵箱中培養14 d。用PBS清洗6孔板,晾干后加入0.1%結晶紫染色1 h左右,清洗并拍照,觀察克隆形成情況,并計算克隆形成數。
1.5 K562細胞周期蛋白檢測 采用Western blotting法檢測細胞中的細胞周期依賴性激酶4(CDK4)、細胞周期蛋白D1(Cyclin D1),以β-actin為內參。
1.6 K562細胞凋亡情況觀察 ①調亡細胞:采用流式細胞術。取兩組K562細胞,測定總細胞數。取1×106個細胞用含10% FCS、1%疊氮化鈉的冰冷PBS重懸,再向每根離心管中添加100 μL細胞懸液。按照1∶200加入7AAD和Anexin V染料,室溫孵育30 min,離心洗滌后上機檢測凋亡細胞。②調亡相關蛋白:采用Western blotting法檢測細胞中的凋亡相關蛋白Bcl-2樣蛋白x(Bax)、cleaved Caspase-3,以β-actin為內參。
1.7 低表達miR-100的MSC中差異表達生長因子檢測 收集感染miR-100-NC和miR-100-inhibitor慢病毒的MSC,采用qRT-PCR法檢測細胞中已知參與白血病細胞增殖調節的生長因子,結果顯示低表達miR-100的MSC中骨形態發生蛋白4(BMP4)基因表達顯著上調(分別為0.12±0.02、1.62±0.31,P<0.05)。
1.8 K562細胞培養液中BMP4檢測 根據“1.3”進行細胞分組與處理,采用ELISA法檢測K562細胞培養液中的BMP4。
1.9 BMP抑制劑Noggin對K562細胞增殖、凋亡的影響觀察 將K562細胞分為對照組、低表達組和實驗組,對照組細胞與感染miR-100-NC慢病毒的MSC共培養;低表達組和實驗組細胞與感染miR-100-inhibitor慢病毒的MSC共培養,實驗組共培養1 d后加入Noggin蛋白。培養2 d后收集K562細胞,采用MTT實驗和平板克隆實驗檢測細胞增殖能力,流式細胞術檢測凋亡細胞,采用Western blotting法檢測CDK4、Cyclin D1、Bax、cleaved Caspase-3,以β-actin為內參,方法同上。
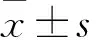
2 結果
2.1 低表達組與對照組細胞增殖能力比較 低表達組細胞共培養72 h細胞增殖能力高于對照組(P<0.05)。見表1。低表達組、對照組共培養3 d克隆形成數分別為(59±0.05)、(71±0.09)個,低表達組高于對照組(P<0.05)。
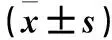
表1 兩組細胞增殖能力比較
2.2 低表達組與對照組細胞周期蛋白表達比較 低表達組細胞中Cyclin D1、CDK4相對表達量分別為6.25±0.17、8.21±0.49,對照組Cyclin D1、CDK4相對表達量均為1,低表達組Cyclin D1、CDK4相對表達量高于對照組(P均<0.05)。
2.3 低表達組與對照組細胞凋亡情況及凋亡相關蛋白表達比較 低表達組、對照組細胞凋亡率分別為15.9%±1.08%、25.13%±2.31%,低表達組細胞凋亡率低于對照組(P<0.05)。低表達組Bax、cleaved Caspase-3相對表達量分別為0.12±0.02、0.23±0.01,對照組分別為1、1,低表達組Bax、cleaved Caspase-3相對表達量低于對照組(P均<0.05)。
2.4 低表達組與對照組培養液中BMP4水平比較 低表達組、對照組培養液中BMP4水平分別為(598±0.28)、(386±0.21)pg/mL,低表達組培養液中BMP4水平高于對照組(P<0.05)。
2.5 外源性noggin對K562細胞增殖、凋亡的影響 實驗組共培養72 h后細胞增殖能力低于低表達組,低表達組增殖能力高于對照組(P均<0.05)。見表2。實驗組、低表達組、對照組克隆形成數分別為(86±0.26)、(202±0.68)、(102±0.30)個,實驗組克隆形成數少于低表達組,低表達組克隆形成數高于對照組(P均<0.05)。實驗組CDK4與Cyclin D1相對表達量低于低表達組,低表達組CDK4與Cyclin D1表達高于對照組(P均<0.05)。見表3。實驗組、低表達組、對照組細胞凋亡率分別為13.60%±0.65%、0.25%±0.32%、11.20%±0.02%,實驗組細胞凋亡率高于低表達組,低表達組凋亡率低于對照組(P均<0.05)。實驗組Bax、cleaved Caspase-3相對表達量高于低表達組,低表達組Bax、cleaved Caspase-3相對表達量低于對照組(P均<0.05)。見表3。
表2 三組細胞增殖能力比較
表3 三組細胞中CDK4、Cyclin D1、Bax、cleaved Caspase-3相對表達量比較
3 討論
miRNA的異常調節與AML的發生發展密切相關[10]。前期有研究報道miRNA在白血病中呈現不同的表達趨勢,其中miR-100低表達[11,12]。然而miR-100是如何調節AML的進程,當前知之甚少。
MSC是白血病微環境中重要的支持細胞。在獲得特定的基因修飾后,MSC能夠被轉化為對維持造血干細胞或白血病干細胞增殖和穩態有利的支持細胞[13]。Lena等[14]報道在MSC中敲除IKBα或PARγ小鼠皆能發生骨髓增殖性腫瘤。MSC作為微環境中的一員對骨髓增殖性腫瘤甚至是白血病的發生發展至關重要。有報道表明,MSC中的miR-100可通過外泌體的形式參與mTOR/Hif1α信號調控,進而影響乳腺癌的惡性進程[15]。本研究結果顯示,miR-100敲低的MSC能夠促進白血病細胞的增殖,減少細胞凋亡,細胞周期蛋白與凋亡相關蛋白表達也相應發生變化。這表明miR-100敲低后,MSC可能通過某種機制影響白血病細胞的增殖和凋亡。細胞周期的調控主要由細胞周期調控蛋白決定[16]。本研究顯示,與miR-100低表達的MSC共培養后,CDK4在K562細胞中呈現高表達,提示通過靶向CDK4可能會在miR-100低表達的AML中具有轉化治療前景。
MSC對白血病細胞的影響至少有兩種,一是分泌細胞因子直接影響白血病細胞的功能[17],二是通過分泌趨化因子募集其他白細胞(如單核細胞)對白血病細胞的功能產生影響[9]。本研究采用體外實驗模擬兩種細胞之間的作用,細胞因子的篩選成為本研究的重點。我們對報道過參與白血病細胞增殖調節的細胞因子進行篩選,結果顯示BMP4蛋白上升水平最高。BMP信號通路調控干細胞穩態,同時也參與了腫瘤的發生發展[17]。但是,BMP信號通路是否調控白血病細胞的增殖,目前仍然知之甚少。本研究結果顯示,與miR-100敲低的MSC共培養的K562細胞培養液中BMP4水平升高,提示MSC可能參與BMP4的分泌;而抑制BMP后,K562細胞增殖受到抑制、凋亡增多,提示miR-100敲低后K562細胞增殖、凋亡的變化可能與BMP4水平變化有關。因此,BMP4很有可能參與形成白血病干細胞的維持、自我更新,并調節白血病細胞的代謝。通過靶向BMP信號通路可能有助于改善miR-100低表達白血病患者的預后。
結合上述研究結果,我們認為,miR-100低表達的MSC可促進AML細胞增殖、抑制其凋亡,可能是通過上調BMP4水平實現的。這一發現將有助于靶向BMP信號通路的特異性藥物的研發和應用。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
