基于SSR分子標記的78份核桃種質資源遺傳多樣性分析
2020-07-23 06:38:28陳利娜李好先唐麗穎曹尚銀
江西農業學報 2020年6期
關鍵詞:資源
敬 丹,駱 翔,陳利娜,李好先,唐麗穎,曹尚銀
(中國農業科學院 鄭州果樹研究所,河南 鄭州 450009)
核桃(JuglansregiaL.)屬于胡桃科、核桃屬的木本樹種,是世界著名四大干果之一,也是世界上重要的油料樹種,與油桐、油茶、烏桕并稱我國四大木本油料樹種[1]。核桃具有較高的經濟價值,其中葉、青皮、核仁、木材以及核桃林本身的經濟價值都很高[2]。核桃還具有重要的營養藥用價值,核桃仁是上好的滋補品,含有多種營養成分,對人體健康及飲食健康起到了重要作用。我國是核桃屬植物的起源和分布中心之一,栽培歷史悠久,種質資源豐富,分布廣泛[3]。加強對種質資源的收集和保護,既是對優良基因的一種保護,又是種質資源創新的前提。一般來說農家品種對自生境有較強的適應性,含有更多優良基因。然而核桃優良的農家品種資源分布較散,往往分布在山地、丘陵地區,收集存在著一定的障礙和困難,國內也尚未有專門單位對其進行收集。此外,將收集來的農家品種進行鑒定和分類保存不僅需要專門的種質資源圃,也需要耗費大量的人力、物力。再者由于傳統的形態學鑒定方法存在一定的短板之處,隨著核桃品種的多樣化,一些親緣關系較為接近的品種僅靠形態學鑒定方法難以鑒別品種間的差異,導致研究者對農家品種的重視程度不高[4]。因此,加強核桃優良農家品種的收集,建立一種有效的苗木品種資源鑒定方法,對于核桃種質資源的創新及核桃產業的發展尤為重要和迫切。
分子標記技術是在形態標記、細胞標記和生化標記后出現的一種新興的技術手段,它以DNA多態性為基礎,直接反映DNA水平上的遺傳變異[4]。目前常用的分子標記有限制性片段長度多態性(RFLP)、隨機擴增多態性DNA(RAPD)、擴增片段多態性(AFLP)、單核苷酸多態性(SNP)、簡單重復序列(SSR)等[1]。在果樹的育種工作中,分子標記技術可用于研究果樹種質資源及親緣關系的遠近、分析其遺傳多樣性,這對于果樹種質資源的評價和保存具有重要意義,同時還可利用分子標記技術輔助育種,加快育種效率。目前,分子標記技術已被廣泛應用于植物遺傳研究和育種實踐中,如桃、蘋果、梨、核桃、茶樹等[4-5]。Freeman等利用SSR技術對不同地域茶樹品種的親緣關系及居群的分化程度進行了分析[6]。張鵬等對42份云南玉米自交系進行了遺傳多樣性分析,為云南地區玉米育種提供了參考依據[7]。在核桃方面的應用主要集中在核桃種群遺傳結構分析、遺傳標記開發等方面[8-9]。目前核桃中已有數百對特異性SSR引物,它們已被證明在核桃的多種遺傳分析中具有實用價值[10]。但是大部分是針對核桃的優良品種之間的遺傳多樣性及親緣關系的研究,而對于核桃農家品種資源的相關研究尚未見報道。本研究旨在收集分布在全國各地的核桃農家品種資源,利用SSR標記對所收集到的核桃農家品種及部分栽培種進行分子標記遺傳多樣性分析,以期為核桃農家品種資源的保存和利用以及核桃種質資源的創新奠定一定的工作基礎。
1 材料與方法
1.1 試驗材料
試驗材料為收集來的78份核桃資源的幼嫩葉片,采后用錫箔紙包裹,在液氮中速凍,然后帶回置于-80 ℃超低溫冰箱保存,所有材料均采自中國農業科學院鄭州果樹研究所品種資源圃(表1)。
1.2 試驗方法
1.2.1 DNA 提取 采用CTAB法提取78份核桃葉片的基因組DNA。利用1%的瓊脂糖凝膠以及Bio-Photometer核酸檢測儀(Eppendorf)檢測DNA的質量、濃度與純度,并將DNA樣品的濃度稀釋至50 ng/μL,置于-20 ℃冰箱保存備用。
1.2.2 SSR分析 以78份核桃的基因組DNA為模板,根據已發表的NCBI公共數據庫中核桃的SSR引物,在各材料中進行擴增,所有引物均由尚亞生物技術有限公司合成。所用反應體系為20 μL的PCR反應體系,其中10×PCR buffer 2 μL,2.5 mmol/L的MgCl21.6 μL,2.5 mmol/L 的4×dNTP 1.2 μL,4 μmol/L的上、下游引物各0.8 μL,5 U/μL的Taq DNA聚合酶0.1 μL,30 ng/μL的DNA 2 μL,用ddH2O補至體積為20 μL。PCR反應程序為:94 ℃ 4 min;94 ℃ 40 s,45~60 ℃ 40 s,72 ℃ 1 min,35個循環;72 ℃ 10 min;4 ℃保存。PCR 擴增產物用6%非變性聚丙烯酰胺凝膠在DYY-Ⅱ型垂直板電泳儀及 DYC-30 型電泳槽中檢測。吸取PCR產物2 μL進行加樣,上樣完畢后在20 mA電流條件下電泳40 min。電泳結束后進行銀染。
1.2.3 譜帶的記錄及數據統計與分析 根據各分子標記在相同電泳遷移率 (相同分子量片段) 的有無對銀染后的凝膠圖像進行條帶統計,在相同位置上,有DNA擴增條帶記為“1”,無帶記為“0”。利用PowerMarker V3.25軟件計算等位基因數(NA)、多態信息含量(PIC)及遺傳距離(GD)等信息。利用NTsys 2.1軟件采用SAHN功能進行UPGMA(類平均法)聚類分析。利用Structure 2.2軟件進行78份核桃資源群體結構分析,其中“Length of Burnin Period”參數設為10000,“Number of MCMC Reps after Burnin”設為50000,K設置為1~10,每個K值重復20次。最佳K值根據L(K)和ΔK隨K值的變化確定[11],其中L(K)及ΔK的計算參照曾可為等[12]的方法進行。
2 結果與分析
2.1 78份核桃農家品種資源SSR標記多態性分析
在78份核桃農家品種資源中,一共從25對SSR引物中篩選出8對具有一定多態性的SSR引物,如表2所示。

表2 SSR標記引物信息
對這8對引物進行分析發現,等位基因(NA)數量變幅為1~16,共擴增出了55個多態性位點,平均每對引物擴增出的等位基因數為6.875。其中引物M02的擴增產物最多,擴增出16條帶;最少的為引物W08,僅有1條帶。此外,主等位基因頻率(MAF)、基因多樣性(Gene Diversity)、多態信息含量(PIC)之間也存在一定的差異。每個SSR位點的PIC在0.075~0.228,平均為0.151。MAF范圍為0.823~0.959,平均為0.893。基因多樣性范圍為0.078~0.271,平均為0.173(表3)。
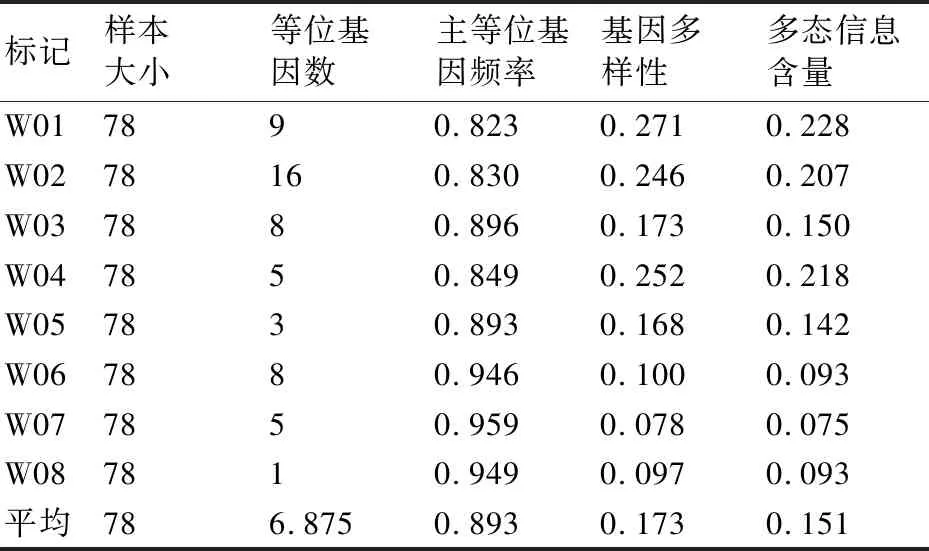
表3 8對SSR引物擴增結果及多態性信息
2.2 78份核桃品種間遺傳多樣性分析
基于這8對引物,采用UPGMA方法對78份資源進行聚類分析。78份核桃品種資源之間的遺傳關系較為接近,遺傳相似系數在0.605和1.000之間發生變化,其中隴南L(H1)和高嶺4號(H18)、宋南3東4(H17)和中短05(H3)、濟西11號(H21)和壽長核桃1號(H37)、響水湖懷柔5號(H22)和汾陽綿核桃10號(H28),安家灘4號(H25)、抗晚霜核桃(H26)和王河核桃2號(H27)、宋南2東5(H5)和中短22(H8)之間的遺傳相似系數較大,遺傳距離較小,說明這些品種間的差異較小,遺傳關系較為接近。由圖1可知,當Neis遺傳距離為0.704時,可將78份核桃資源分為5部分,分別為Ⅰ、Ⅱ、Ⅲ、Ⅳ和Ⅴ。其中Ⅰ和Ⅱ品種較多,Ⅰ包括65個品種,占供試材料的83.33%。Ⅱ包括9個品種,占供試材料的11.54%。Ⅲ和Ⅳ分別僅包含GL4(H72)、西藏核桃(H69)一個品種,分別來自北京和西藏。Ⅴ包含兩個品種,即宋南5東6(H76)和宋南2東9(H74),兩者均來源于河南鞏義。此外,該群體內的平均遺傳距離為0.23,遺傳距離0.2~0.3所占比例最大,為26.86%;遺傳距離0.5以內的占96.38%(圖2)。
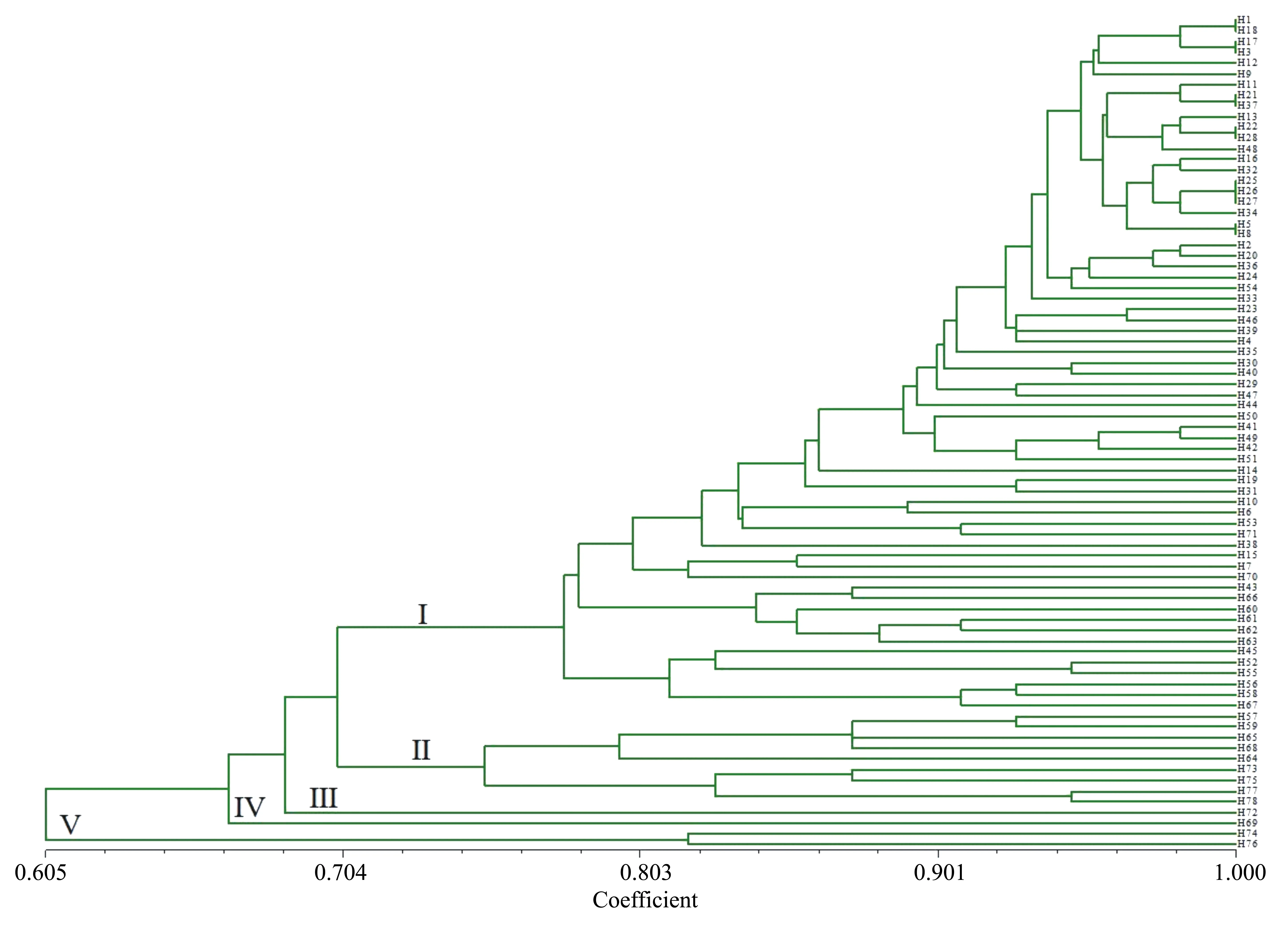
圖1 78份核桃資源的UPGMA聚類結果
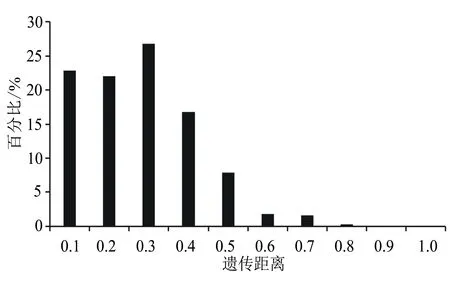
圖2 78份核桃資源的遺傳距離分布
2.3 群體結構分析
利用Structure 2.2軟件對78份核桃資源進行群體結構分析,結果發現隨著K值的增加,對數極大似然值L(K)值呈現遞增的趨勢,無明顯的轉折點(圖3A);但ΔK值隨K值增加呈先增加后下降的趨勢。根據最大K值確定最佳亞群數[13],當K=2時,ΔK值出現最大峰值(圖3B),由此說明該群體最適可劃分為2個亞群(圖4),分別為Q1(紅色)和Q2(綠色)。其中Q1包含25份核桃資源,主要來自河南(10份),其余來源于北京、甘肅、新疆及河北等地;Q2包含53份核桃資源。78份資源未明顯按照地理來源進行群體劃分,該結果與UPGMA聚類結果相似,說明群體的劃分與地理來源不完全相關。此外有31份種質既含有紅色部分,又包含綠色部分,這些資源可能是兩個祖先亞群雜交而來。
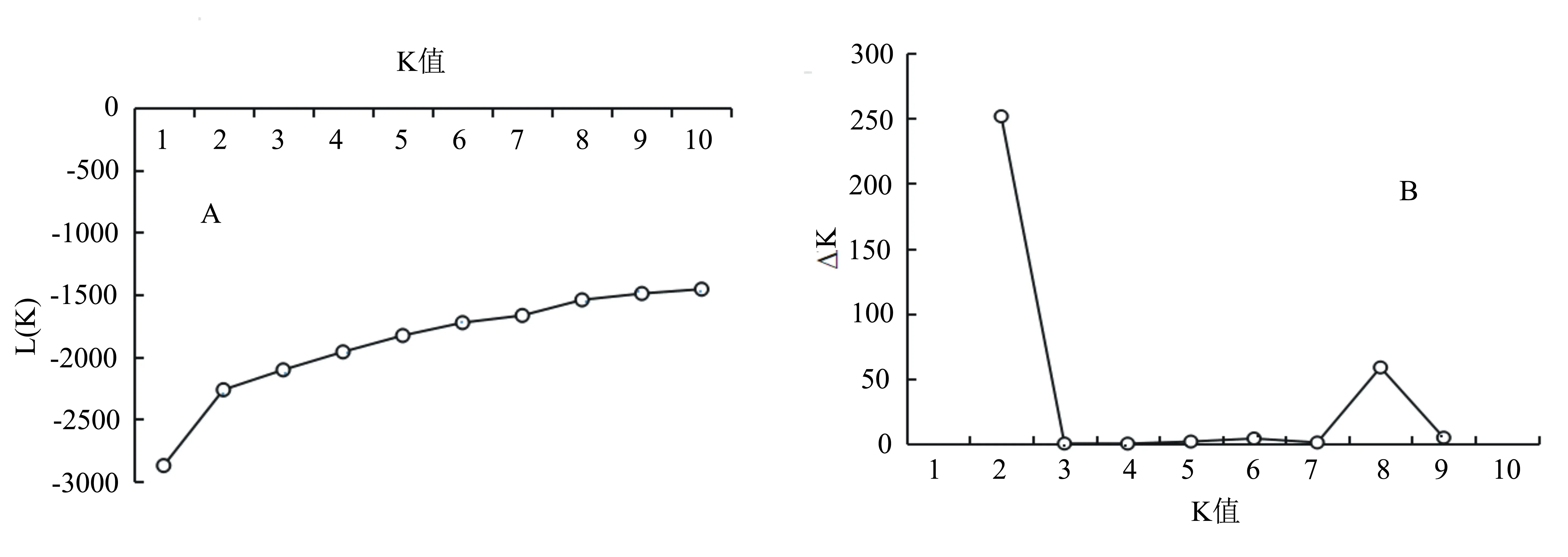
A為78份核桃資源的L(K)與K值關系圖;B為ΔK與K值關系圖。圖3 78份核桃資源的L(K)、ΔK與K值關系圖

圖4 當K=2時78份核桃資源的群體結構
3 結論與討論
不同物種適應新環境的基礎是具備一定的遺傳多樣性,遺傳多樣性水平越高,該物種或種群對新環境的適應能力就越強[14-15]。目前遺傳多樣性的研究已從傳統的形態學標記研究向分子標記研究轉變,利用分子標記技術對多種核桃資源進行品種間的鑒定與分類不斷取得進展。中國是核桃起源地之一,栽培歷史悠久,種質資源豐富,核桃因其較好的食用價值、經濟價值被推薦為世界各地人類健康飲食的重要資源,受到了廣泛的歡迎,但中國核桃產業良種化程度不高,品種、類型混雜,難以區分。利用分子標記技術對一定群體內的核桃種質資源進行遺傳特性分析,對于種質資源的區分、評價和保存及優良種質的選育具有重要的意義[16]。Wang H等[10]利用SSR標記揭示了西藏地區核桃及泡核桃的遺傳多樣性和雜種優勢,發現西藏核桃群體具有平均水平的遺傳多樣性和大量的稀有等位基因。周于波等利用SSR技術,對四川29份核桃良種資源進行了遺傳多樣性分析和聚類分析,增強了品種間的區分能力[1]。與前人研究相比,本研究所選78份試材大部分為農家品種資源,一般來說,地方性品種對自生境適應性較強,對分散于全國各地的核桃農家品種資源進行遺傳多樣性分析,了解農家品種現有資源的多樣性及其結構,可為遺傳資源的保護和優良種質的篩選提供重要信息。
本研究利用25對SSR引物對78份核桃資源進行遺傳多樣性研究,最終篩選出8對引物,可加強對78份資源的鑒別。從78份供試材料共擴增出55個多態性條帶,平均每對引物擴增出6.875個多態性位點。楊本蕓等[17]利用10對引物對21份核桃品種進行擴增,每對引物平均擴增出6.7個多態性位點,多態性位點的相同與否可能與引物本身的多態性有關。78個不同品種間的遺傳相似系數變化范圍在0.605~1.000之間,而遺傳相似系數可作為判斷品種間親緣關系遠近的標準[1],本研究品種間遺傳相似系數較大,遺傳距離較小,平均遺傳距離為0.23,由此說明該研究所采用的78份資源之間親緣關系較為接近,遺傳背景較為相似。這與陳良華等[18]對四川核桃遺傳多樣性的研究中野生核桃群體親緣關系相近的結果相似。基于遺傳相似系數,對78份材料進行UPGMA聚類分析,在遺傳系數為0.704時,將群體分為5部分,分析發現栽培種與農家品種以及不同地理來源的核桃品種界限不明顯,存在相互滲透的現象,也說明遺傳聚類組的劃分與地理來源不完全相關。姚明哲等[19]對江北地區茶樹資源進行研究發現也存在該現象,四川的茶樹品種被聚類到重慶茶樹群體中。此外利用Structure 2.2軟件對核桃群體進行遺傳結構分析,發現最佳亞群體數為2,群體結構并不復雜,但存在相互滲透現象,可能存在種質的遺傳變異或者基因交流[20]。
種質資源的研究是育種工作的一個重要基礎,正確評價核桃種質資源的遺傳多樣性對其種質資源的創新具有重要意義。利用分子標記技術,同時結合形態學標記等方法對不同核桃品種的遺傳多樣性進行系統的研究與評價,可為培育核桃優良新品種、提高核桃現有種質資源的品質及抗性提供重要的科學理論依據。本研究78份核桃資源間的遺傳關系相似,大部分品種間親緣關系較為接近,因此應加強品種資源的有效利用,加強材料共享與交流,利用分子標記技術輔助育種,提高育種效率。
猜你喜歡
江蘇安全生產(2023年1期)2023-02-08 05:58:38
資源節約與環保(2022年8期)2022-09-20 02:25:22
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
藝術品鑒(2020年7期)2020-09-11 08:04:44
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
當代貴州(2018年28期)2018-09-19 06:39:04
資源再生(2017年3期)2017-06-01 12:20:59
決策(2015年9期)2015-09-10 07:22:44
