重慶市庫蚊標本中宜昌病毒的分離鑒定
2020-07-17 06:31:28付士紅何小周何亞明殷啟凱趙潔榮馮盼盼陳曉菁馬學軍梁國棟許松濤王環宇
中國人獸共患病學報 2020年6期
李 樊,付士紅,何小周,何亞明,葉 盛,邵 楠,殷啟凱,趙潔榮,馮盼盼,祿 陽,陳曉菁,何 英,凌 華,馬學軍,梁國棟,許松濤,王環宇
重慶市位于中國內陸西南部、長江上游地區,長江橫貫其全境。面積8.24萬km2,轄38個區縣(26區、8縣、4自治縣)。地貌以丘陵、山地為主,有“山城”之稱。重慶屬亞熱帶季風性濕潤氣候,氣候溫暖濕潤、適宜媒介昆蟲孳生繁殖。 因氣候、環境、地理條件等原因,重慶市一直以來是乙型腦炎的高發地區,因此在當地開展蟲媒調查,對于了解當地蚊蟲乙腦病毒帶毒率及分子遺傳特征,了解該地區蚊蟲攜帶病毒的種類及多樣性,以及蟲媒病毒病的防控都具有重要意義[1-3]。
Yichang病毒是一種昆蟲病毒,屬于套式病毒目(Nidovirales)Mesoniviridae病毒科,最早于2014年中國湖北省宜昌市采集的蚊蟲標本中被分離出來,湖北省多個地區的蚊蟲標本中也曾檢測到該病毒的核酸[4]。目前,對于該病毒的宿主范圍、流行地理范圍尚不清晰。本研究對2017年在重慶市開展蟲媒病毒調查的過程中從庫蚊標本中分離到Yichang病毒進行報道。
1 材料和方法
1.1標本采集 2017年9月初在重慶市巴南區和豐都縣的人房和豬圈采用吸入式捕蚊器在晚間5-9點誘捕采集成蚊,當日回收成蚊放置-20 ℃冰箱冷凍30 min,剔除雄蚊并按不同種屬對雌蚊分類,30~50只/管裝至凍存管,編號后迅速保存于液氮罐中。
1.2細胞培養 實驗所用細胞為白紋伊蚊細胞(Aedes albopictus cell,C6/36)、金黃地鼠腎細胞(BHK-21)、非洲綠猴腎細胞(Vero),由中國疾病預防控制中心病毒病預防控制所保存。C6/36細胞培養液為1640培養基(Gibco)含10%胎牛血清(Fetal bovine serum, FBS, Gibco)和100 U/mL青霉素和鏈霉素。 BHK-21、Vero細胞培養液為MEM培養基含10% FBS和100 U/mL青霉素和鏈霉素。
1.3病毒分離 在生物安全柜內向每支蚊蟲標本中,加入1 mL含100 U/mL青霉素和鏈霉素的MEM培養液, 使用組織研磨機(美國QIAGEN公司)對標本進行研磨, 將研磨后的標本于4 ℃,18 000×g,離心20 min。吸取上清液150 μL用0.22 μmol/L過濾器進行過濾后,接種到已長成70%~80%的單層C6/36、Vero、BHK-21細胞,吸附1 h后,加入1 mL含2% FBS的細胞維持液,分別置于5% CO2培養箱中,逐日觀察細胞病變(Cytopathic effects,CPE)情況,當CPE達到75%時收獲培養上清液,再次接種細胞;以相同程序盲傳3代,有病變的分離物繼續進行鑒定分析。
1.4核酸提取及高通量測序 取陽性分離物上清液,用QIAamp viral RNA mini kit(QIAGEN公司)按說明書提取病毒RNA,采用Superscript III反轉錄酶(Thermo Fisher)和隨機引物合成cDNA,采用NEBNext mRNA Second Strand Synthesis Module(美國NewEnglandBiolabs公司)完成第2鏈DNA合成。再用Nextera XT DNA Library Preparation Kit (美國Illumina公司)完成文庫構建。該文庫在Qubit 4.0熒光計(Thermo Fisher)定量后,在Illumina iseq平臺上進行雙端測序(2×150堿基)。 根據平均質量值(Q30)評價測序質量并過濾數據。將過濾后數據拼接生成Contig,在美國國家生物技術信息中心(NCBI)病毒基因組數據庫中(http://www.NCBI.nlm.nih.gov)進行BLAST比對。將過濾后數據用useglaxy(https://useglaxy.org/)中的Bowtie 2比對目標基因組,并生成Consensus序列。使用Open Reading Frame Finder(https://www.ncbi.nlm.nih.gov/orffinder)預測基因組開放閱讀框。獲得病毒的全基因序列已提交NCBI基因庫,登錄號為MT070763。
1.5系統發育分析 使用MAFFT V7(https://www.ebi.ac.uk/Tools/)進行多序列比對。利用NCBI BLAST對測序結果進行核苷酸與氨基酸序列一致性分析。從GenBank中選取與陽性序列相似的參考毒株,用MEGA X構建進化樹。
2 結 果
2.1病毒分離 2017年9月,在重慶市巴南區和豐都縣共采集到732只蚊蟲標本(圖1),其中騷擾阿蚊672只(92%),致倦庫蚊30只(4%),三帶喙庫蚊30只(4%)。這些蚊蟲被分為20批次接種細胞。從重慶地區豐都縣致倦庫蚊標本分離物在接種C6/36細胞后72 h開始引起細胞CPE。CPE的主要特征是細胞變圓、皺縮、甚至脫落(圖2),120 h后細胞完全病變。而BHK-21和Vero細胞在3次盲傳過程中均未觀察到CPE。該分離物被定名為CQFD2017。
地圖中重慶市所在區域以淺灰色標識,豐都縣和巴南區以深灰色標識。圖1 蚊蟲標本采集地點Fig.1 Locations where mosquito samplings were performed
2.2CQFD2017基因組序列分析 為鑒定所分離的病毒,對病毒基因組進行高通量測序分析和系統進化分析。 CQFD2017的組裝基因組為20 893 bp,3′端含有poly(A)尾。該基因組鑒定出6個開放閱讀框:ORF1a、ORF1b、ORF2a、ORF2b、 ORF3a、ORF3b。ORF1a跨越核苷酸317-8 635 nt,并編碼2 772個氨基酸;ORF1b跨越核苷酸8 614-16 413 nt,編碼2 600 個氨基酸;ORF2a跨越核苷酸16 473-19 181 nt, 編碼902個氨基酸; ORF2b跨越核苷酸16 436-17 110 nt,編碼224個氨基酸; ORF3a跨越核苷酸19 224-19 709 nt,編碼161個氨基酸; ORF3b跨越核苷酸19 534-19 908 nt,編碼124個氨基酸; 而5′和3′非編碼區的長度分別為316 bp和529 bp。拼接組裝獲得contig經BLASTx分析表明,它與Mesoniviridae病毒科的Yichang病毒(NC_040534)最為匹配。組裝獲得的CQFD2017全基因組與Yichang病毒的核苷酸一致性有98.6%,ORF1a和ORF1b區域的氨基酸一致性分別為99.06%和99.15%,CQFD2017與Mesoniviridae病毒科其他成員的的氨基酸一致性在22%~51%之間(表1)。
2.3CQFD2017的系統發育分析 為了研究CQFD2017的系統進化地位,從NCBI 的GenBank下載了與CQFD2017相似性≥30%的RNA依賴的RNA聚合酶(RNA-dependent RNA polymerase, RdRp)氨基酸序列,并用于構建系統發育樹。分析表明,CQFD2017屬于Mesoniviridae病毒科,為Yichang病毒。CQFD2017與冠狀病毒科的形成了不同的進化分支,與Nam Dinh病毒、DaK Nong病毒、Hana病毒等構成了Mesoniviridae病毒科進化簇(圖3)。

(A)C6/36細胞對照;(B)CQFD2017感染細胞72 h; (C) CQFD2017感染細胞120 h。比例尺為200 μm圖2 CQFD2017對C6/36細胞致病變作用Fig.2 Cytopathic effects (CPEs) caused by CQFD2017 in C6/36 cells
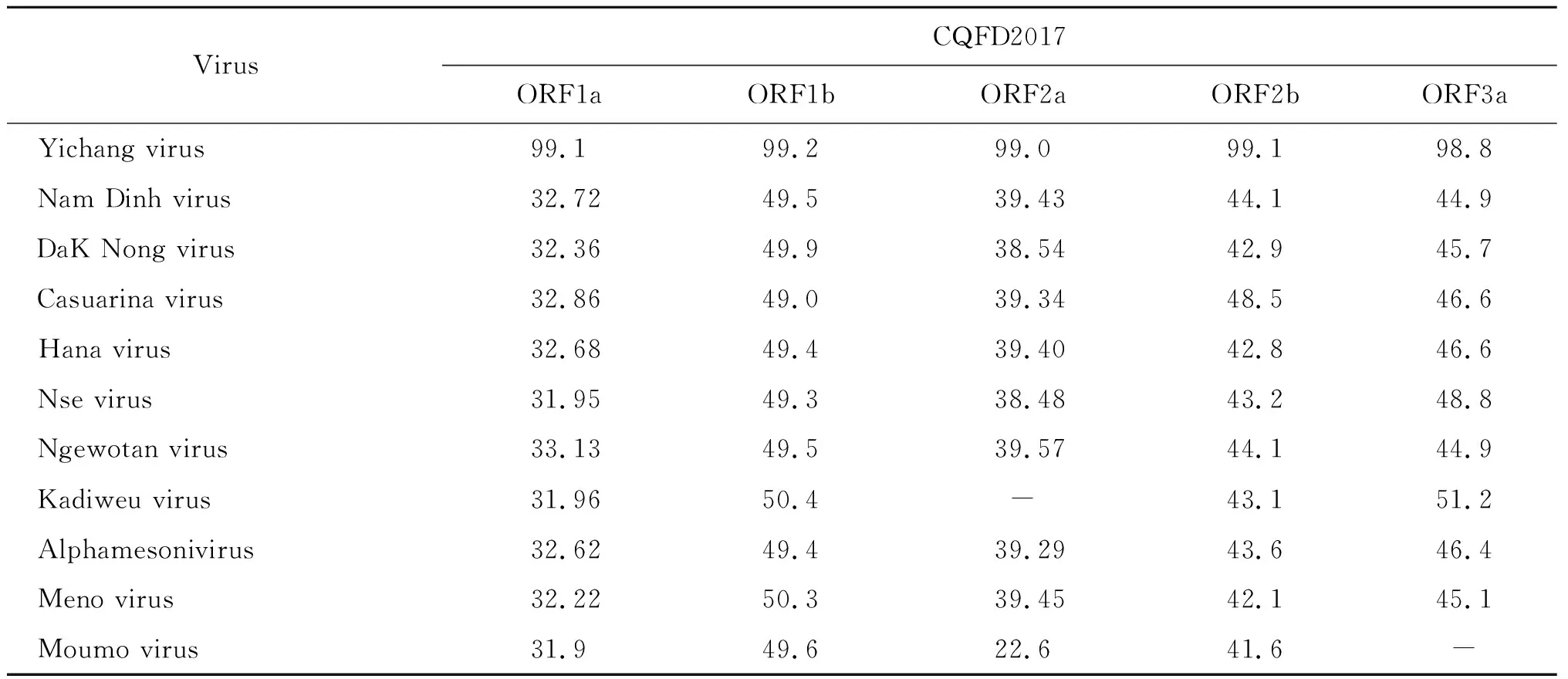
表1 CQFD2017與Mesoniviridae病毒科家族成員的氨基酸序列比對分析Tab.1 Amino acid identities between CQFD2017 and other members of family Mesoniviridae

注:實心圓表示CQFD2017分離株;分支節點上的校驗值表示經1000次重構后相應分支出現的概率;隱藏分支節點上小于60%的校驗值。圖3 基于宜昌病毒分離株CQFD2017的RdRp區氨基酸序列的系統進化分析Fig.3 Neighbor-joining tree inferred from molecular phylogenetic analysis based on the amino acid sequences of CQFD2017 RdRp segments
3 討 論
套式病毒目(Nidovirales)病毒是一類主要感染哺乳動物, 少數感染禽類及無脊椎動物的有囊膜的單股正鏈RNA病毒,也是目前已知的含有最長基因組的RNA病毒。根據2016年國際病毒分類委員會對RNA病毒的分類,目前該病毒目包含4個病毒科:動脈炎病毒科(Arteriviridae)、冠狀病毒科(Coronaviridae)、桿狀套病毒科(Roniviridae)及近些年被鑒定出來的(Mesoniviridae)[5]。 以引起藍耳病的豬繁殖與呼吸綜合征病毒(PRRSV)為代表的動脈炎病毒科基因組RNA約12.7~15.7 kb,被稱作為“小”基因組套式病毒;冠狀病毒科(包括SARS病毒、MERS病毒等)、桿狀套病毒科的基因組RNA長約26.3~31.7 kb, 被稱作“大”基因組套式病毒; 新發現的Mesoniviridae科病毒(Nam Dinh病毒、DaK Nong病毒等)基因組大小位于上述兩者之間,被稱之為 “中等”套式病毒[6-7]。隨著高通量測序技術的發展和應用,越來越多套式病毒目的成員被分離鑒定[4, 8-10]。近年來,Mesoniviridae病毒科成員被從亞洲(越南、泰國、韓國、印度尼西亞和中國)、非洲(科特迪瓦)、大洋洲(澳大利亞)和北美洲(美國)等多地的蚊蟲和蝙蝠中分離出來,表明了這類病毒地理分布的廣泛性[8, 10-16]。
2017年9月,本課題組在重慶市的巴南區和豐都縣開展了蟲媒調查,并從豐都縣采集的庫蚊標本中分離到Yichang病毒CQFD2017株。經對其細胞生物學水平及分子遺傳學水平的研究發現,該病毒與2014年袁志明教授團隊分離自中國湖北省宜昌市庫蚊標本的Yichang病毒為同種病毒,核苷酸序列高度一致達到98.6%[4]。
重慶市和宜昌市均位于長江的干流上,三峽水庫位于重慶和宜昌之間,含重慶市22個區縣市及湖北省4個縣市。而Yichang病毒在2014年和2017年在三峽地區的多次被分離或檢測到,表明該病毒在三峽地區自然界的蚊蟲中已形成穩定的流行。但前期研究結果顯示,該病毒僅能在白紋伊蚊細胞C6/36上引起CPE,而在哺乳動物細胞BHK-21和靈長類細胞Vero上并未觀察到CPE。 但與Yichang病毒同屬Mesoniviridae病毒科、分離于2002年越南腦炎患者腦脊液中的Nam Dinh病毒已被懷疑是引起人類腦炎的新型病毒。目前對于Yichang病毒在蚊蟲間的傳播機制及中間宿主的種類尚不明確,該病毒是否能引起人獸疾病尚有待進一步探討。考慮到該病毒具有引起人獸疾病的潛在風險,未來有必要在三峽地區開展的蟲媒監測中持續關注該病毒,跟蹤其流行傳播趨勢及分子遺傳變異特征。
(感謝重慶市疾病預防控制中心、巴南區疾病預防控制中心、豐都縣疾病預防控制中心的同志在蚊蟲標本采集過程中給予的配合和幫助。)
利益沖突:無
