基于核磁共振代謝組學分析糖尿病大鼠腦區特異性代謝變化
2020-06-11 11:49:02倪志濤季慧高紅昌鄭宏張華杰
溫州醫科大學學報 2020年5期
關鍵詞:糖尿病
倪志濤,季慧,高紅昌,鄭宏,張華杰
(溫州醫科大學藥學院,浙江溫州325035)
糖尿病是一種因體內胰島素分泌缺陷或生物功能受損而導致的全身代謝性疾病[1]。糖尿病往往會出現涉及全身多個器官的嚴重并發癥,其發病率逐年增加[2]。早在1922年就發現了糖尿病能引起認知功能障礙[3],其表現為智力減退和靈活性降低等[4]。糖尿病腦病作為糖尿病的并發癥之一會嚴重降低患者的生活質量[5],但是其發病機制尚不明確。
代謝組學是繼基因組學和蛋白質組學之后新發展起來的一門學科,是系統生物學的重要組成部分[6]。基于核磁共振代謝組學技術能夠系統地分析機體受到疾病干預情況下代謝物變化[7],其憑借樣品制備簡單、分析速度快、重現性高等優勢廣泛應用于腦代謝研究[8]。現階段的腦代謝研究主要集中在與學習記憶密切相關的海馬組織,而對其他腦區的關注較少,因此本研究運用基于核磁共振代謝組學技術分析糖尿病大鼠大腦皮層、紋狀體、下丘腦和中腦等腦區的代謝變化,為探索糖尿病腦病的潛在代謝機制提供新線索。
1 材料和方法
1.1 儀器、試劑與動物 BrukerAVANCEIII600核磁共振譜儀(德國Bruker公司);冷凍干燥機(ALPHA-4,德國Christ公司)。血糖儀及血糖試紙(德國貝朗醫療國際貿易有限公司);重水(D2O,99.9%)購于美國劍橋同位素實驗室;鏈脲佐菌素(streptozocin,STZ)購自美國Sigma-Aldrich公司;枸櫞酸、枸櫞酸鈉、甲醇、氯仿購于上海國藥集團化學試劑有限公司。雄性SD大鼠20 只,體質量180~200g,購于上海斯萊克實驗動物有限公司,飼養于溫州醫科大學實驗動物中心SPF級動物房,室溫(22±2)℃,相對濕度45%±3%,動物許可證號:SYXK(滬)2017-0008。本研究通過實驗動物倫理學委員會批準。
1.2 方法
1.2.1 建立1 型糖尿病大鼠模型:雄性SD大鼠適應性飼養1周后隨機分成糖尿病組和對照組,每組8只。糖尿病組大鼠禁食12h后分別按體質量單次腹腔注射65mg/kg的STZ枸櫞酸鈉混懸液,對照組給予相同體積的枸櫞酸鈉溶液,72h后分別測其空腹血糖水平,以大鼠血糖值大于16.7mmol/L代表造模成功。
1.2.2 大鼠腦組織收集與處理:大鼠經造模成功
10周后斷頭處死,迅速分離出大腦皮層、紋狀體、下丘腦和中腦,并將分離出的組織放入液氮中急凍,之后儲存于-80℃。取樣本稱重后置于勻漿管內,分別加入冰甲醇4mL/g,蒸餾水0.85mL/g,使用組織勻漿機勻漿后渦旋15s;再分別加入2mL/g冰氯仿和冰蒸餾水,渦漩后于冰上靜置15min[9]。4℃、12000×g離心15min后取上層溶液置于凍干機內凍干24h。最后將凍干粉末重新溶解于500μLD2O中,4℃、12000×g 離心10min后將上清液轉入NMR樣品管中進行測試。
1.2.31HNMR圖譜采集與分析:使用BrukerAVANCEIII600MHz超導高分辨核磁共振譜儀采集1HNMR圖譜。采用zgpr標準脈沖序列,僅用預飽和方式壓制水峰。具體參數如下:采樣標準溫度298K,譜寬12000Hz,數據采集點數64K,弛豫延遲10s,累加次數256次,每次掃描時間2.65s。采集到的圖譜均采用BrukerTopspin2.1軟件進行相位及基線調整,并以乳酸甲基峰的化學位移(1.33ppm)進行定標。將同組樣本導出的數據一起導入到MATLABR2010軟件(美國MathWorks公司)中進行相位調整,然后對譜峰段(0.5~9.0ppm)以0.01ppm的間隔進行積分切割。為了消除負值和水峰的影響,將各組樣本數據中的負值和水峰(4.60~5.20ppm)置零。對每一段積分值都相對于該譜的所有積分值進行歸一化[10-11],然后將數據導入SIMCA-P+12.0軟件(瑞典Umetrics公司)進行主成分分析(principalcomponentanalysis,PCA),所得譜圖的X軸和Y軸分別代表第一主成分(firstprincipalcomponent,PC1)和第二主成分(secondprincipalcomponent,PC2)。
1.3 統計學處理方法 應用SPSS13.0統計軟件進行統計學處理。計量資料以表示,2組間比較采用獨立樣本t 檢驗。P <0.05 為差異有統計學意義。
2 結果
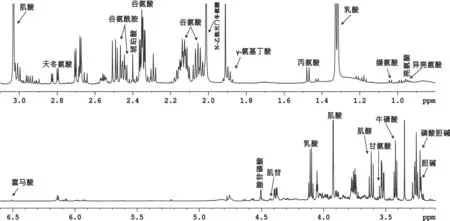
圖1 糖尿病大鼠大腦皮層組織典型1HNMR代謝指紋圖譜
2.1 糖尿病大鼠腦區代謝模式分析 圖1為大鼠大腦皮層的典型1D1HNMR譜,紋狀體、下丘腦、中腦的譜圖與大腦皮層譜圖極為相似,故不做展示。通過相關文獻,并結合NMRSuite7.0軟件和人類代謝數據庫(www.hmdb.ca)進行分析鑒定,歸屬出的代謝物有亮氨酸、異亮氨酸、纈氨酸、乳酸、丙氨酸、γ-氨基丁酸、N-乙酰天門冬氨酸、肌酸、膽堿、磷酸膽堿、牛磺酸、甘氨酸、腺苷磷酸、肌苷、富馬酸、天冬氨酸、琥珀酸、谷氨酸、谷氨酰胺、肌醇。2組大鼠4個腦區代謝物的PCA分析結果表明,糖尿病組和對照組大鼠4個腦區代謝物在PC1方向上均有明顯區分,代謝模式差異明顯(見圖2)。從相對應的載荷圖中可以發現乳酸、肌酸、谷氨酸、N-乙酰天門冬氨酸等在大腦皮層的代謝模式區分中貢獻較大。谷氨酸、谷氨酰胺、γ-氨基丁酸、肌醇、牛磺酸等在紋狀體的代謝模式區分中貢獻較大。乳酸、肌醇、磷酸膽堿、牛磺酸、γ-氨基丁酸等在下丘腦的代謝模式區分中貢獻較大。另外,乳酸、谷氨酸、谷氨酰胺、亮氨酸、磷酸膽堿等在中腦的代謝模式區分中貢獻較大。
2.2 腦區代謝物定量分析結果 對代謝物進行定量分析發現,與對照組大鼠相比,糖尿病組大鼠大腦皮層中乳酸、牛磺酸、肌酸、磷酸腺苷、谷氨酸、纈氨酸、天冬氨酸等代謝物含量顯著上升,而甘氨酸、磷酸膽堿、肌醇、富馬酸、谷氨酰胺、γ-氨基丁酸等代謝物出現下降(P <0.05),見圖3。與對照組大鼠相比,糖尿病組大鼠紋狀體中乳酸、牛磺酸、磷酸膽堿、磷酸腺苷的含量顯著升高,而丙氨酸、甘氨酸、肌酸、膽堿、肌苷、亮氨酸、異亮氨酸、纈氨酸、N-乙酰天門冬氨酸、γ-氨基丁酸等代謝物出現下降(P<0.05),見圖4。與對照組大鼠相比,糖尿病組大鼠下丘腦中肌醇、牛磺酸、磷酸膽堿含量顯著上升,而膽堿、N-乙酰天門冬氨酸、琥珀酸、亮氨酸、谷氨酸、谷氨酰胺、γ-氨基丁酸等代謝物出現下降(P<0.05),見圖5。與對照組大鼠相比,糖尿病組大鼠中腦中乳酸、肌醇、磷酸膽堿的含量顯著上升,而丙氨酸、磷酸腺苷、N-乙酰天門冬氨酸、肌醇、琥珀酸、亮氨酸、異亮氨酸、谷氨酸、谷氨酰胺、γ-氨基丁酸、丙氨酸、富馬酸等代謝物出現顯著下降(P<0.05),見圖6。
3 討論
糖尿病患者出現認知功能損傷的確切機制至今不明,研究人員更多關注于持續高血糖造成的神經元功能紊亂、氧化應激、糖基化終產物產生增多等機制。而本研究從基于1HNMR的代謝組學角度出發,分析糖尿病對不同腦區代謝所帶來的影響,有助于更加深入了解糖尿病腦病的發病機制。
3.1 糖尿病大鼠腦區特異性能量代謝紊亂 大腦正常生理活動所需的能量占人體能量總消耗的20%以上,腦病的發生與腦內能量代謝變化也有著密切聯系[12-13]。本研究發現,與對照組相比,糖尿病大鼠的大腦皮層、紋狀體、中腦三個腦區中乳酸的含量異常升高。這一結果與報道[14]的糖尿病大鼠海馬組織乳酸含量異常升高相類似,提示糖尿病大鼠腦內會出現廣泛的乳酸代謝異常。腦內乳酸由葡萄糖通過無氧酵解的方式生成,其含量的升高通常被認為與線粒體功能受損和丙酮酸氧化能力下降有關。此外,三羧酸循環(tricarboxylicacidcycle,TCA)中間產物富馬酸、琥珀酸在大腦皮層、下丘腦和中腦中出現不同程度下降,這說明在這三個腦區中三羧酸循環活性減弱。所以糖尿病大鼠腦組織出現的糖酵解能力及三羧酸循環活性變化是引起能量變化的重要原因。
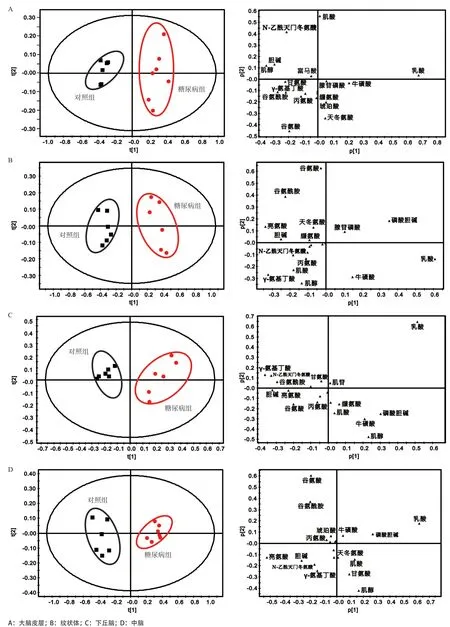
圖2 2組大鼠4個腦區代謝模式分析的PCA得分圖及其對應載荷圖

圖3 糖尿病大鼠大腦皮層的代謝通路分析圖
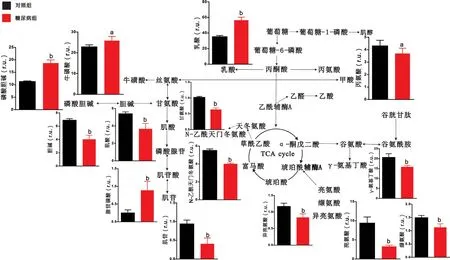
圖4 糖尿病大鼠紋狀體的代謝通路分析圖
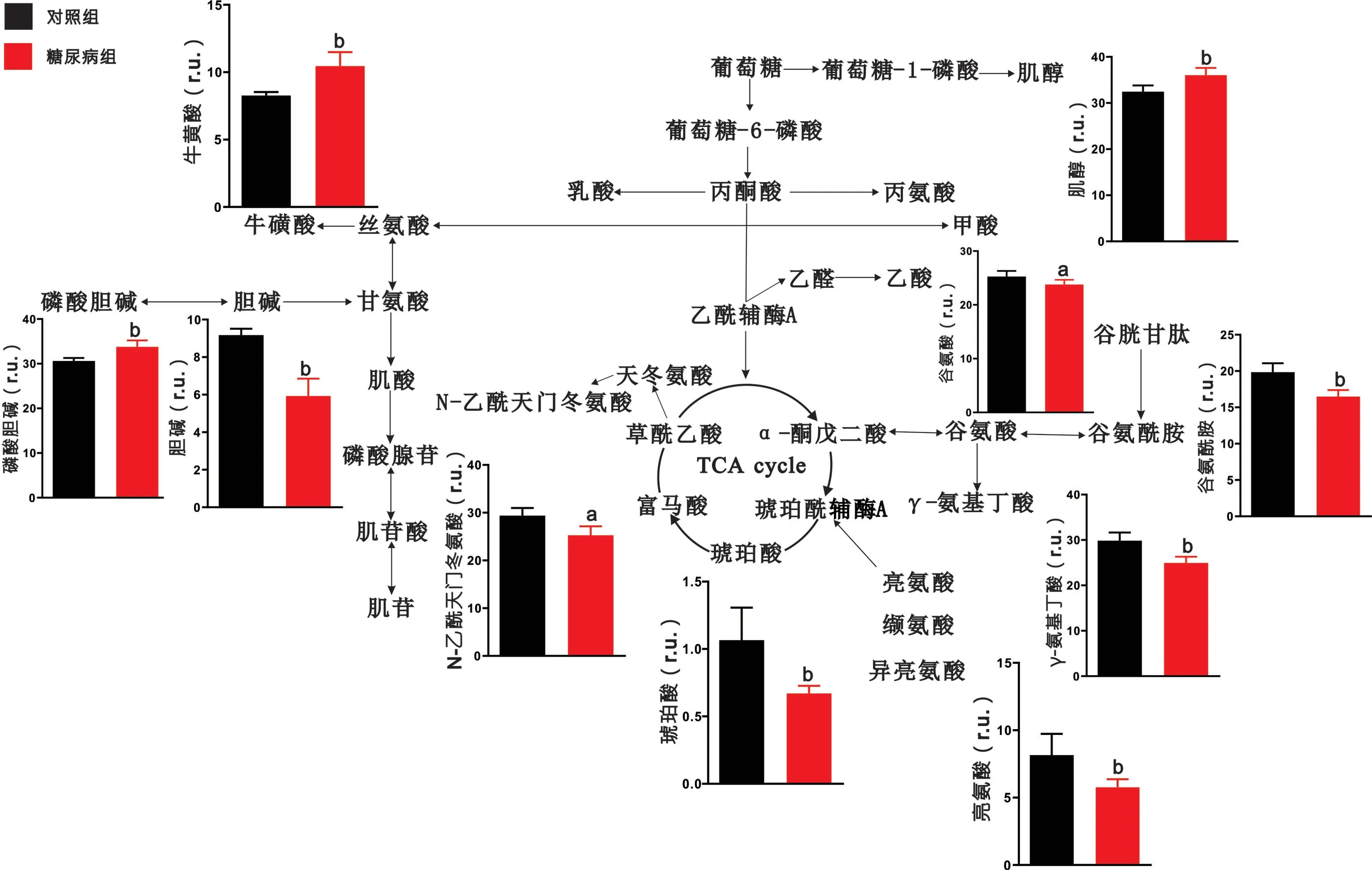
圖5 糖尿病大鼠下丘腦代謝通路分析圖
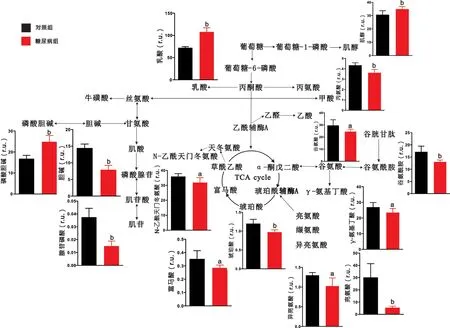
圖6 糖尿病大鼠中腦的代謝通路分析圖
3.2 糖尿病大鼠腦區特異性神經遞質代謝紊亂 谷氨酸-谷氨酰胺(Glu-Gln-GABA)循環維持著神經元和膠質細胞之間的神經遞質平衡[15]。星形膠質細胞攝取谷氨酸后,將其轉化為谷氨酰胺,隨后將合成的谷氨酰胺轉運至神經元,作為氨基酸類神經遞質谷氨酸和γ-氨基丁酸的主要前體物質,繼續參與神經沖動傳遞或其他代謝途徑[16]。本研究發現,與對照組相比,在糖尿病大鼠的大腦皮層中的谷氨酰胺和γ-氨基丁酸的含量出現降低,而在下丘腦、中腦兩個腦區中,谷氨酸、谷氨酰胺和γ-氨基丁酸的含量均出現顯著降低,說明神經元和膠質細胞之間的神經遞質循環被嚴重抑制。在2型糖尿病db/db小鼠的全腦代謝分析中同樣出現了類似變化,這提示神經遞質代謝紊亂無論是在1型還是2型糖尿病腦病中都至關重要[17]。此外紋狀體中γ-氨基丁酸的含量也出現了顯著降低。抑制性神經遞質γ-氨基丁酸作為腦內最重要的神經遞質之一,其含量降低也會嚴重影響正常腦功能。天冬氨酸作為興奮性氨基酸類神經遞質能夠參與到TCA循環的補給途徑中,我們發現大腦皮層中γ-氨基丁酸含量降低的同時天冬氨酸的含量卻顯著升高,這一結果進一步說明了神經遞質代謝出現紊亂。
3.3 糖尿病大鼠腦區特異性神經膠質細胞標志物變化 牛磺酸和肌醇都是神經膠質細胞的特征性代謝物[18-19],普遍認為牛磺酸可反映星形膠質細胞的活性。我們發現糖尿病大鼠的大腦皮層、紋狀體和下丘腦中的牛磺酸含量都出現了顯著提高,提示這三個腦區的膠質細胞出現增生,而膠質細胞的增生已被發現于糖尿病大鼠腦中[20]。此外有研究發現糖尿病大鼠海馬膠質細胞增生與海馬神經元的凋亡有著緊密聯系[21],因此我們推測上述三個腦區的神經元也出現了不同程度的損傷。同時牛磺酸也具有抗氧化作用,這提示糖尿病大鼠腦內可能出現了氧化應激損傷。肌醇是調節膠質細胞滲透壓平衡和內環境穩定的重要代謝物。本研究發現,與對照組相比,糖尿病大鼠大腦皮層中肌醇含量出現下降,這提示神經膠質細胞功能異常,膠質細胞滲透壓調節能力失效;而下丘腦和中腦卻出現完全相反的結果,膠質細胞的滲透壓調節顯著增強。
3.4 糖尿病大鼠腦區特異性膽堿代謝異常 膽堿和磷酸膽堿是生物膜的重要組成成分,膽堿和磷酸膽堿含量的變化與細胞膜損傷有關[22]。本研究發現,與對照組相比,在糖尿病大鼠大腦皮層中,磷酸膽堿的含量出現顯著的下降而膽堿含量無明顯變化。這一結果提示大腦皮層細胞膜的損傷較為嚴重,細胞的更替能力下降。
3.5 糖尿病大鼠腦區特異性神經元標志物變化 N-乙酰天門冬氨酸作為神經元活性的特征性代謝物,一般認為其含量的降低可反映神經元的損傷[23]。本研究結果發現,與對照組相比,N-乙酰天門冬氨酸在糖尿病大鼠的紋狀體、下丘腦和中腦中都出現顯著下降,說明糖尿病對大鼠腦神經元造成較大損傷。因此,高血糖所誘導的神經元損傷是廣泛涉及多個腦區的,而并不只存在于海馬區域[24]。
綜上所述,本研究應用基于1HNMR的代謝組學方法分析了1型糖尿病大鼠大腦皮層、紋狀體、下丘腦和中腦四個腦區的代謝水平變化,發現糖尿病大鼠出現腦區特異性代謝紊亂,主要包括能量代謝、神經遞質代謝和膽堿代謝。
猜你喜歡
中老年保健(2022年5期)2022-08-24 02:35:42
中老年保健(2022年1期)2022-08-17 06:14:56
中老年保健(2021年5期)2021-08-24 07:07:20
中老年保健(2021年9期)2021-08-24 03:51:04
中老年保健(2021年7期)2021-08-22 07:42:16
中老年保健(2021年3期)2021-08-22 06:49:56
中老年保健(2021年11期)2021-08-22 03:15:16
中國生殖健康(2020年2期)2021-01-18 02:51:44
中國生殖健康(2018年2期)2018-11-06 07:11:04
基層中醫藥(2018年2期)2018-05-31 08:45:04
