2017-2018年陜西省H7N9禽流感病毒流行特征及基因進化分析
2020-04-13 08:42:54張俊君焦亞東張少白余鵬博
中國人獸共患病學報 2020年2期
史 偉,李 慎,吳 瑞,張俊君,李 俊,高 濤,焦亞東,張 蕾,徐 藝,張少白,余鵬博
2013年長三角地區的上海市和安徽省先后報告H7N9流感病毒引起的人間感染病例,這也是全球首次報道的人類感染該亞型的流感病毒[1]。截至2017年4月,全球共報告1 344例人感染H7N9流感確診病例,其中死亡511人[2]。迄今為止,陜西省共報告7例人感染H7N9流感病例和2起禽間H7N9疫情。其中2017年5月,咸陽市一例重癥肺炎病例在西安被確診為H7N9病例,為陜西省首例H7N9流感病毒導致的人類感染病例[3],此后又發現6例人感染病例[4]。
為評估H7N9流感病毒暴露風險,為防控工作提供科學依據,本研究對2017年5月至2018年3月陜西省H7N9流行分布和基因特性進行整理和分析。
1 材料與方法
1.1樣本采集 陜西省在每個地市均設有監測點,長年開展外環境禽流感病原學監測任務[5]。采樣方法參考《職業暴露人群血清學和環境高致病性禽流感監測方案(2011版)》[6]的要求,采集禽類糞便、禽類飲水、禽籠具表面擦拭等類型環境標本。
H7N9報告病例檢測及禽間疫情暴露環境監測:按照人感染H7N9禽流感疫情防控方案(第3版)》[7]和《人感染H7N9禽流感病毒標本采集及實驗室檢測策略》[8]進行樣本采集和流行病學調查。
1.2病毒檢測與毒株分離 待檢樣本暫存于4 ℃并于24 h內送至當地流感網絡實驗室。核酸提取采用凱杰公司QIAamp Viral RNA Mini Kit (Cat No.52904)提取。所有樣本先開展甲型流感初篩,陽性樣本再進行H7亞型檢測。流感分型試劑由江蘇碩世生產。國家流感中心對陽性樣本進行復核和雞胚毒株分離,并對毒株利用Illumina平臺進行全基因組測序;PCR核酸檢測陽性但未分離出雞胚株的環境樣本直接進行測序[9]。測序結果上傳至中國流感病毒基因序列數據庫。
1.3病毒序列分析 采用MEGA X軟件[10]對核苷酸推導出的氨基酸序列進行位點變異比對并構建遺傳進化樹,進化樹采用Neighboring-joining法進行繪制,Bootstrap重復值為1000。導出結果用FigTree進行美化(http://tree.bio.ed.ac.uk/software/figtree)。參考毒株的基因序列來自GISAID(Global Initiative on Sharing Avian Influenza Data,全球共享禽流感數據倡議組織數據庫)。
2 結 果
2.12017-2018年外環境H7N9病原監測情況 2017年5月至2018年3月,陜西省142個監測點共采集活禽市場外環境標本2 442份,檢出H7N9陽性標本12份,陽性率為4.91‰。
2.2H7N9報告病例及暴露環境監測情況 陜西省共發現H7N9病例7例,其中包括:咸陽[3]和榆林的兩起聚集性病例,以及西安和寶雞報告的散發病例。其中5例發病前有活禽接觸史。
對所有本地病例暴露環境進行環境監測發現:咸陽采集環境樣本192份,結果均為陰性;西安采集環境樣本209份,H7N9陽性樣本11份;寶雞采集各類環境樣70份,H7N9陽性樣本3份。
另外,根據世界動物衛生組織報告,陜西省榆林市[11]和銅川市各發現1起禽間禽流感疫情。其中榆林市報告家禽死亡約2.2萬只,銅川市死亡蛋雞810只。具體信息和地理分布情況分別見表1和圖1。
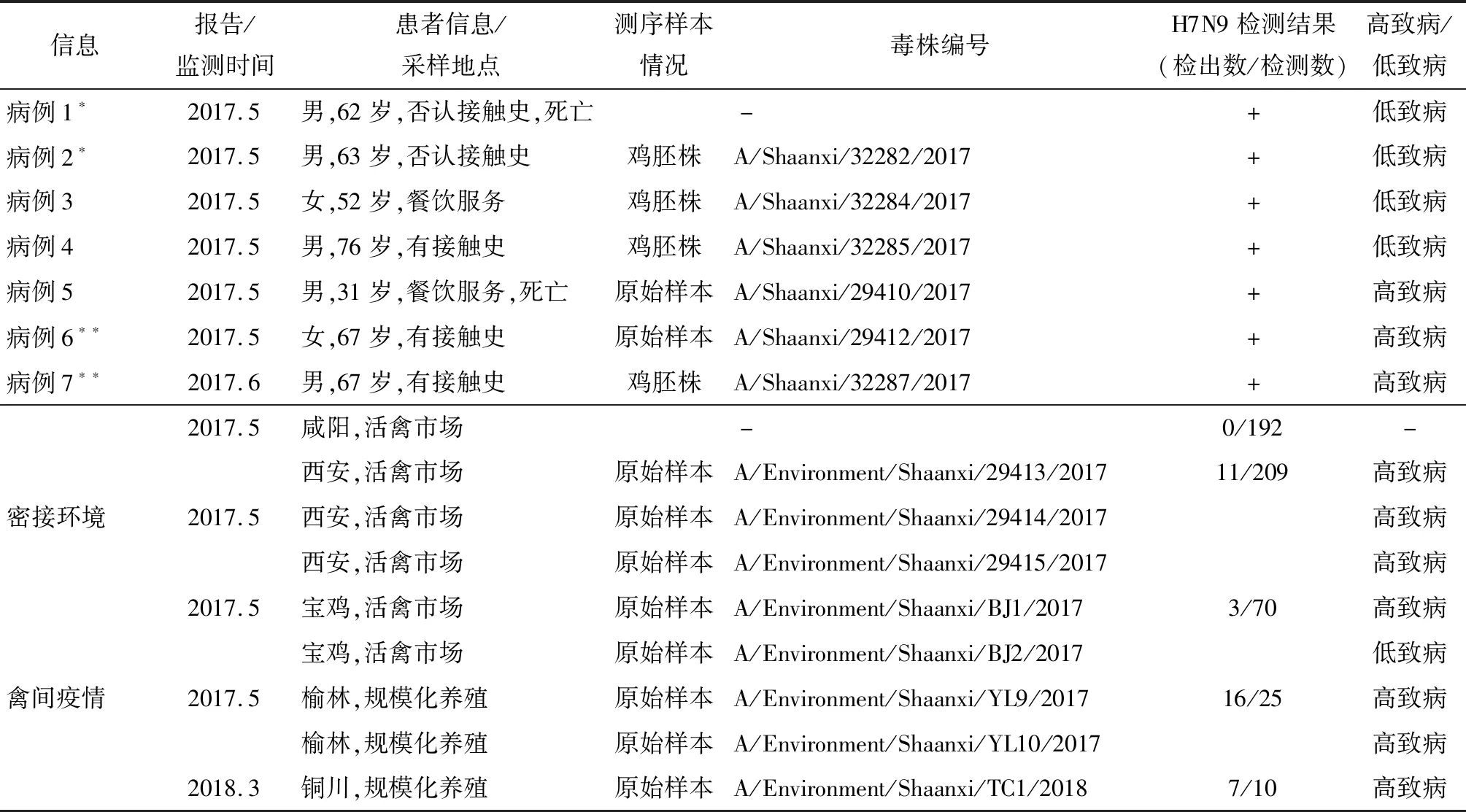
表1 2017年至2018年陜西省H7N9標本基本信息及檢測結果Tab.1 Informations and results of specimens from Shaanxi Province from 2017 to 2018
注:*和**為聚集病例
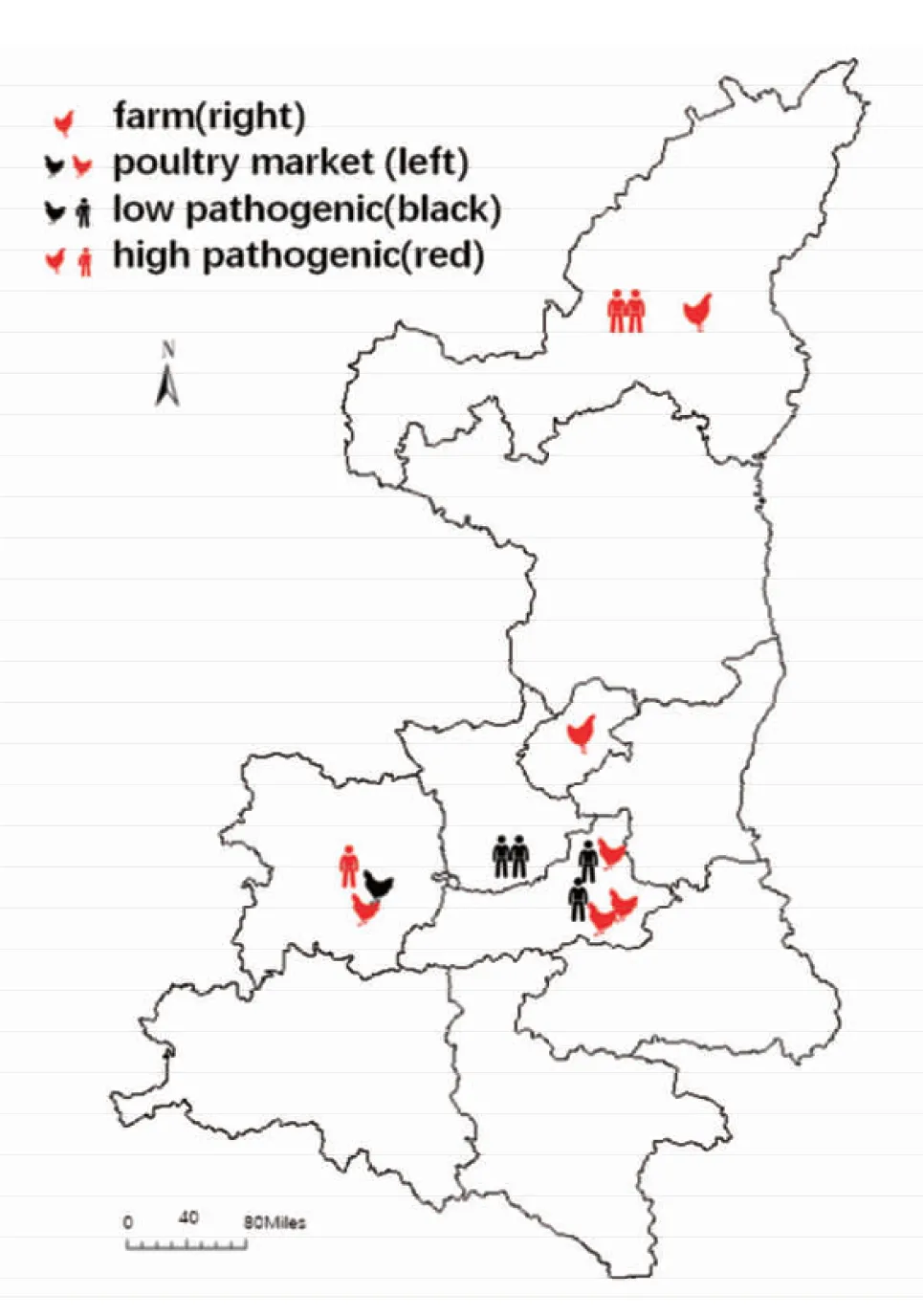
注:圖例中雞/人(紅)代表感染高致病性H7N9,雞/人(黑)代表感染低致病性H7N9;雞頭向右代表樣本來自規模化養殖場,向左代表樣本來自活禽市場圖1 2017-2018年陜西省H7N9病毒變異株分布情況Fig.1 Distribution of H7N9 virus mutants in Shaanxi Province from 2017 to 2018
2.3陜西省H7N9流感病毒HA和NA核苷酸序列同源性分析 選擇A/Anhui/2013和A/Guangdong/17SF062/2017、H7N9重組疫苗株A/duck/Fujian/S4170/2014為參考株,運用MEGA X對上述14株病毒HA基因和NA基因同源性進行分析。陜西省分離的H7N9 HA基因核苷酸的同源性分別為99.17%~99.29%、99.28%~99.89%和96.44%~96.58%;NA基因核苷酸與上述H7N9流感病毒代表株的核苷酸的同源性為99.09%~99.24%、98.51%~98.67%、95.89%~96.27%。
2.4H7N9流感病毒種系進化樹分析 寶雞、榆林、西安、銅川分離毒株HA氨基酸發生插入突變為PEVPKRKRTAR↓GLF,咸陽、西安、寶雞為PEIPKGR↓GLF。據報道,長江三角洲地區是公認的原始來源地,珠三角地區作為H7N9的額外暴發源[12]。從進化上將H7N9分為長江三角洲分支(Yangtze River Delta)和珠江三角洲分支(Pearl River Delta),而陜西省分離到的毒株均屬于長江三角洲分支。陜西省所發現的H7N9高致病株HA序列與2016年底在廣東最早發現的高致病株高度同源,見圖2左圖所示。NA遺傳進化特點與HA一致,即陜西高致病性H7N9變異株與廣東2016高致病株NA基因高度同源。見圖2右圖所示。
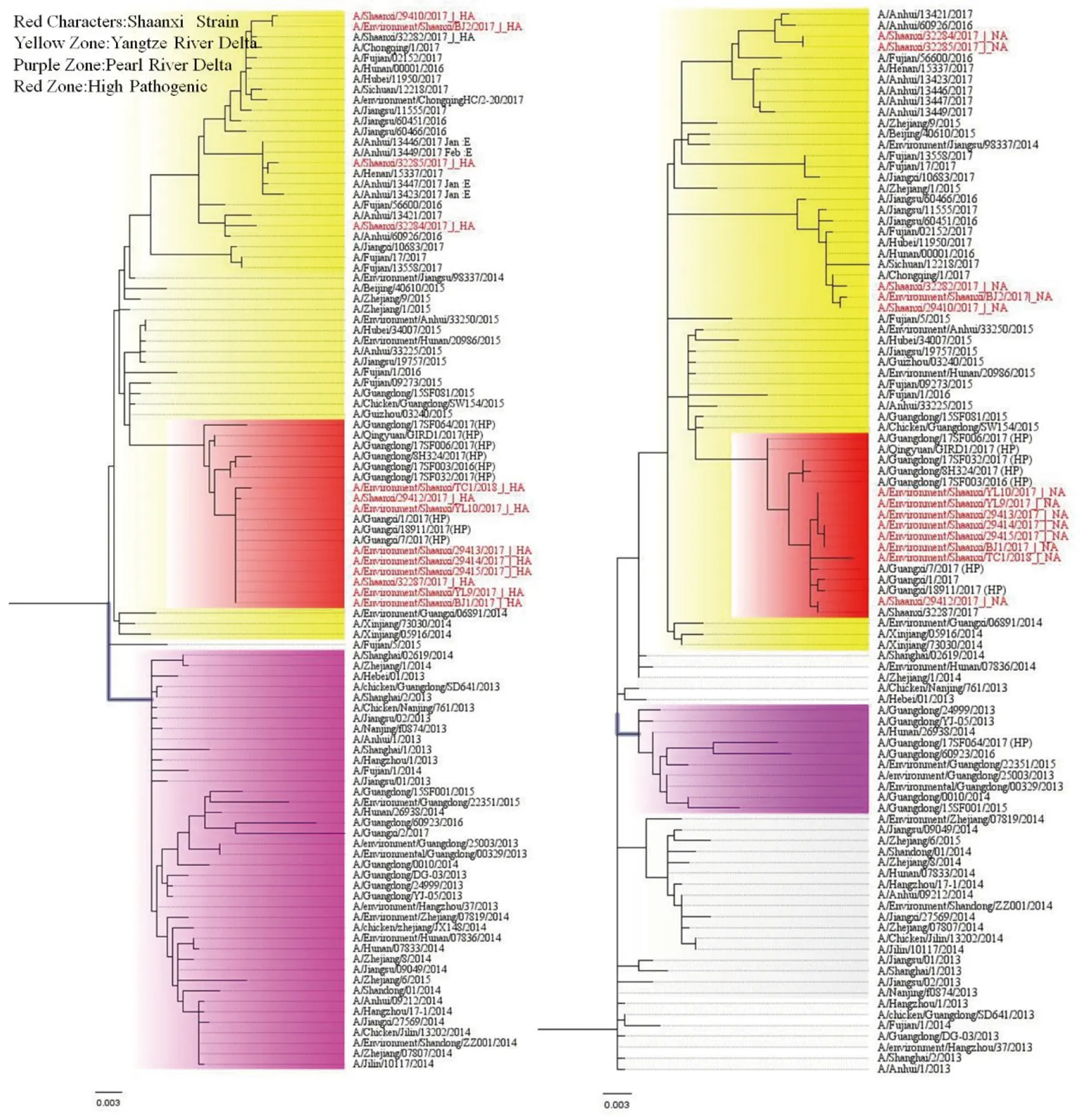
圖2 2017-2018年陜西省H7N9流感病毒HA(左)和NA(右)進化樹Fig.2 Phylogenetic trees for HA (left)and NA(right) genes of the influenza H7N9 viruses in Shaanxi Province
2.5H7N9陜西株編碼蛋白關鍵氨基酸位點分析 結合表1信息和所有毒株核酸序列信息,對H7N9關鍵氨基酸位點發現:寶雞、榆林、西安、銅川分離毒株HA氨基酸發生插入突變為PEVPKRKRTAR↓GLF,系高致病性H7N9流感病毒(High Pathogenic,HP);咸陽、西安、寶雞為PEIPKGR↓GLF,系低致病性H7N9流感病毒(Low Pathogenic,LP)。部分病毒HA蛋白出現Q226L改變,提示這幾株病毒具有結合人類和禽類的雙受體結合特性。3株毒株PB2蛋白分別出現E627V或E627K改變,提示病毒發生與毒力增強相關的突變[13-15]。所有毒株PB1蛋白473V改變,提示在哺乳動物中的復制能力增強[16];M1蛋白30和215位點均發生與毒力增強相關的突變[17];M2蛋白S31N改變,提示所有毒株對烷胺類藥物耐藥[18];NS1蛋白中80-84位點氨基酸未發生缺失[19]。詳見表2。

表2 2017年至2018年陜西省H7N9標本基本信息及檢測結果Tab.2 Information and results of specimens from Shaanxi Province from 2017 to 2018
3 討 論
人感染H7N9流感病毒是2013年首次被發現[1],在之后數年在我國安徽、上海、浙江、福建、廣東等多省均有報道[2, 20]。陜西作為內陸省份,外環境一直未監測到[5]。直到2017年5月才報道首例人感染H7N9病例[3]。據報道,陜西省多個縣區開展活禽養殖業務[21],人群暴露在病毒環境下的可能性較高。因此,評估人群暴露風險、研究本省H7N9禽流感病毒的分子特征至關重要。
2017年5月至2018年3月,陜西省142個活禽市場監測點H7N9核酸陽性率為4.91‰,提示陜西省活禽市場整體污染狀況較低。但從H7N9病例追溯其接觸的活禽市場中環境樣本有病毒檢出,提示從業人員應注意禽流感暴露的風險。
從進化來分析陜西14株H7N9流感病毒基因信息。它們與長三角地區的H7N9高度同源,但卻又分成低致病性和高致病性兩個分支,也反映了病毒來源的復雜性。這可能和兩方面原因有關:一方面陜西有豐富的自然資源,適宜鳥類的生存與遷徙[22-23];另一方面,陜西地處交通要道,周邊省份發現H7N9病例后關閉活禽市場[24],病禽很可能流通到本地。
編碼蛋白關鍵氨基酸位點序列分析發現:H7N9裂解位點的突變導致其對家禽致病力增強,給活禽養殖業帶來巨大威脅。同時,HA蛋白Q226L突變增加了病毒對人α-2,6唾液酸受體結合力增強;PB1蛋白473V改變、PB2蛋白E627V/K改變、M1蛋白30D和215A均提示病毒致病力和對哺乳動物的適應性在增強。M蛋白均發生S31N突變,提醒我們在臨床診療中選擇針對性的藥物[25]。
因此,為更好的防控H7N9流感,需要加強人H7N9流感疫情的檢測并密切關注病毒變異情況。規范活禽市場管理,加強禽類H7N9流感病毒的監測和控制。
利益沖突:無
引用本文格式:史偉,李慎,吳瑞,等.2017-2018年陜西省H7N9禽流感病毒流行特征及基因進化分析[J].中國人獸共患病學報,2020,36(2):124-129. DOI:10.3969/j.issn.1002-2694.2020.00.014
