樹莓1-氨基環丙烷-1-羧酸合酶基因(RiACS)家族成員的克隆及生物信息學分析
2019-10-14 08:12:02周義杰閆希煥高亭豪楊明峰馬蘭青
北京農學院學報 2019年3期
周義杰,閆希煥,高亭豪,劉 芳,楊明峰,馬蘭青,3*
(1.北京農學院植物科學技術學院/農業農村部華北都市農業重點實驗室,北京 102206;2.生物與資源環境學院/農業農村部華北都市農業重點實驗室,北京 102206;3.北京農學院北京林果業生態環境功能提升協同創新中心,北京 102206)
樹莓(Raspberry)是薔薇科(Rusaceae)懸鉤子屬(RubusL.)小漿果類灌木的統稱,又名托盤、山莓、懸鉤子等,是世界公認的第三代新興水果,有“黃金水果”“生命之果”之稱。樹莓在栽培上主要有空心莓亞屬(SubgenusIdeobatus)和實心莓亞屬(SubgenusEubatus)兩個亞屬,其主要區別是空心莓亞屬果實成熟時與花托分離而實心莓亞屬果實成熟時與花托同落。空心莓亞屬有歐洲紅樹莓(Rubusidaeus)、黑樹莓(Rubusoccidentalis)和紫樹莓(Rubusneglectus)3個種群,實心莓亞屬有黑莓、無刺黑莓、匍匐形黑莓和黑莓雜交種4個種群[1]。樹莓果實柔嫩多汁,色澤鮮艷,香氣濃郁,風味獨特,富含多種對人體有益的活性成分,如超氧化物歧化酶、鞣花酸、黃酮、類黃酮、樹莓酮等,具有抗氧化、抗癌、抗菌消炎、抑制肥胖、預防心血管疾病等作用[2-5]。樹莓除了果實能夠鮮食外,還廣泛用于食品、制藥、保健品、化妝品生產等行業,發展前景廣闊[6]。近年來,中國樹莓產業推廣速度較快,種植的面積和產量逐漸增加,但樹莓果實采收時節氣溫較高,果實極不耐貯存,容易在運輸過程腐爛變質,導致樹莓鮮果上市期短,常溫下貨架期只有1~2 d[7],限制樹莓產業的快速發展。
乙烯是一種重要的內源性植物激素,存在于植物的各個器官和組織中,參與調控高等植物的許多生理過程,比如種子萌發,生根,葉片、花器官的衰老、果實成熟和逆境脅迫等[8]。乙烯在植物中合成途徑為:腺苷蛋氨酸合成酶催化蛋氨酸與腺苷酸(adenosine monophosphate, AMP)反應生成腺苷蛋氨酸(s-adenosvlmeIhionine, SAM);ACS催化SAM生成1-氨基環丙烷-1-羧酸(1-aminocyclopropane-1-carboxylate, ACC);ACC氧化酶(1-aminocyclopropane-1-carboxylate oxidase, ACO)催化ACC氧化生成乙烯,其中ACS是整個合成途徑的限速酶[9]。自1990年從番茄果實的cDNA文庫中分離得到第一個ACS基因序列以來[10],研究人員已經在很多植物中分離出ACS基因,所有這些植物中都含有一個以上的ACS基因[11-15],說明ACS是由多基因家族編碼的。
電子克隆(insilicocloning)是隨著基因組計劃和EST(Expressed Sequence Tag)計劃發展起來的基因克隆新技術。與傳統的圖位克隆、轉座子標簽技術、遞減雜交技術等方法相比具有成本低、速度快、操作簡單、目的性強的優勢[16-18]。利用電子克隆技術進行樹莓ACS基因家族成員的克隆,從中尋找與果實成熟相關的ACS基因,可以推進樹莓果實成熟衰老機制的研究和應用,改善樹莓果實不耐貯藏的特性,對于樹莓產業的發展具有重要意義。然而傳統的電子克隆技術依賴大量EST序列進行推測、組裝和拼接來獲得基因的電子序列[19],所以對于EST資料不夠豐富的物種無能為力。與一些模式植物和其他薔薇科經濟植物相比,樹莓的EST資料遠遠達不到傳統電子克隆技術的要求。本研究使用一種新的電子克隆方案,通過ACS家族蛋白質保守結構域與樹莓基因組比對獲得基因的電子序列,然后進行基因組PCR驗證,最終獲得6條ACS基因,并對基因序列和其編碼蛋白的理化性質、蛋白結構、同源性等進行預測和分析,為后續研究樹莓ACS基因家族成員的差異表達和樹莓果實成熟機制奠定基礎。
1 材料與方法
1.1 試驗材料
樹莓植物材料為本實驗室組織培養保存,品種為‘托拉蜜’(Tulameen)。主要試劑有:植物DNA提取試劑盒(Magen,Guangzhou,China),金牌Mix高保真PCR酶(Tsingke,Beijing,China),膠回收試劑盒(Magen,Guangzhou,China),pClone007克隆載體(Tsingke,Beijing,China),DH5-α感受態細胞(Tsingke,Beijing,China),質粒快速提取試劑盒(Magen,Guangzhou,China)。
1.2 試驗方法
1.2.1 樹莓ACS基因家族成員的電子克隆 以NCBI登錄的薔薇科ACS蛋白序列XP_004306735、XP_024176223、XP_021807957和XP_007219056與已知的擬南芥ACS蛋白家族序列[20]進行比對,得到薔薇科ACS蛋白的7個保守結構域序列,將之分別與GDR(Genome Database for Rosaceae)數據庫(https://www.rosaceae.org/)中樹莓基因組進行tblastn比對,得到樹莓ACS基因家族成員的電子序列及其所在染色體的位置信息。
1.2.2 樹莓ACS基因家族成員的分子克隆及序列分析 根據所得電子序列使用Primer Premier 5.0軟件設計引物(表1),由北京瑞博興科生物技術公司合成。取樹莓組培苗葉片200 mg使用植物DNA提取試劑盒提取基因組DNA,然后進行PCR擴增,PCR體系為:金牌mix 45 μL,上游引物(10 μmol/L)2 μL,下游引物(10 μmol/L) 2 μL,模板(100 ng/μL) 1 μL。PCR條件為:98 ℃,2 min;98 ℃ 10 s,Tm (RiACS1-6分別為:54、55、57、52、57、56 ℃) 15 s,72 ℃,45 s,35個循環;72 ℃,5 min。PCR產物經膠回收后連接到pClone007克隆載體并轉化大腸桿菌感受態細胞,經過夜培養后挑取單克隆搖菌并進行菌液PCR,使用PCR呈陽性的菌液提取質粒,提取的質粒送北京睿博興科生物技術有限公司測序。測序所得基因序列使用SnapGene軟件分析GC含量,使用Augustus程序(http://bioinf.uni-greifswald.de/augustus/)分析開放閱讀框(ORF,Open Reading Frame)和內含子,并獲得CDS(Coding Sequences)序列和其所編碼的蛋白序列[21],使用DNAMAN軟件比較基因序列和蛋白序列之間的相似性。

表1 樹莓ACS基因家族成員克隆引物Tab.1 Cloning primers of raspberry ACS family genes
1.2.3 樹莓ACS家族蛋白序列的生物信息學分析 使用ExpPASYProtParam程序(https://web.expasy.org/protparam/)分析蛋白的理化性質[22],使用SignalP 4.1 Server程序(http://www.cbs.dtu.dk/services/SignalP/)進行蛋白質信號肽預測[23],使用TMHMMServer 2.0程序(http://www.cbs.dtu.dk/services/TMHMM/)進行蛋白跨膜結構預測[24],使用PSORT程序(http://psort1.hgc.jp/form.html)進行蛋白亞細胞定位預測,使用SOPMA程序(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)進行蛋白二級結構預測[25],使用SWISS-MODEL軟件(https://swissmodel.expasy.org/)進行蛋白三維結構同源建模[26],使用DNAMAN軟件進行蛋白多序列比對,使用MEGA 7.0軟件生成系統進化樹[27]。
2 結果與分析
2.1 樹莓ACS家族成員的序列獲得及分析
由薔薇科ACS蛋白與擬南芥ACS蛋白序列比對,得到7個薔薇科蛋白保守結構域序列如下:IQMGLAEN、FQDY、FCLADPGDAFL、NPSNPLGT、YSLSKDMG、KMSSFGLVSSQTQ、PGWFRVCFA,將其分別與樹莓基因組進行比對后得到6條樹莓ACS基因家族成員的電子序列,將其按照染色體順序命名。根據電子序列設計引物進行基因組PCR(圖1),將PCR膠回收產物連接克隆載體測序,得到樹莓ACS家族成員的基因序列。所得基因序列已上傳至NCBI數據庫,序列號為 MK388675、
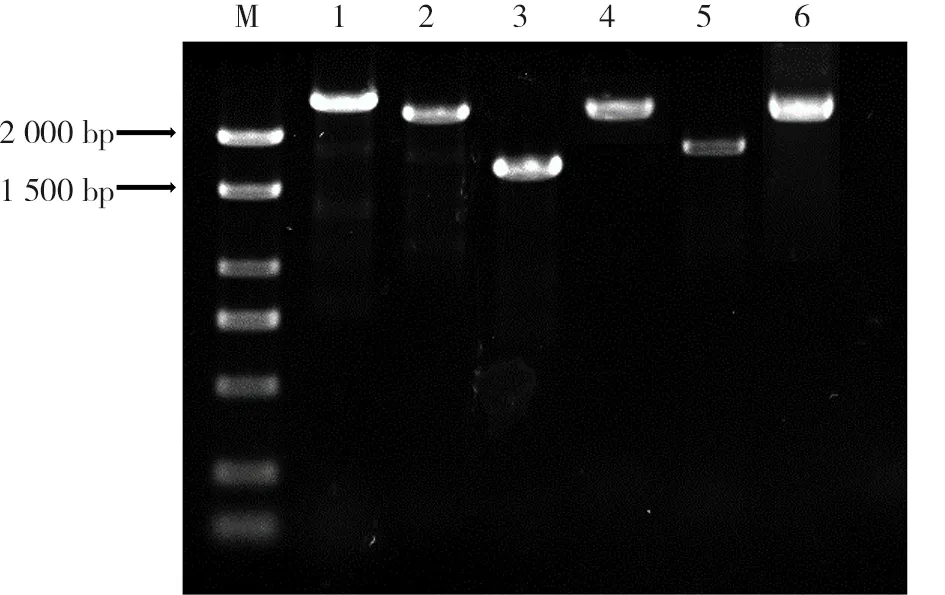
注:M. Marker;1. RiACS5;2. RiACS4;3. RiACS1;4. RiACS6;5. RiACS2;6. RiACS3。Note: M. Marker;1. RiACS5;2. RiACS4;3. RiACS1;4. RiACS6;5. RiACS2;6. RiACS3.圖1 樹莓ACS基因家族成員PCR擴增Fig.1 PCR amplification of raspberry ACS family genes
MK388676、MK388677、MK388678、MK388679、MK388680。對其進行序列分析(表2)發現,RiACS1-6的GC含量均小于50%,與全基因組GC含量吻合[28],ORF長度在1 657~2 432 bp之間,除了RiACS1和RiACS2外其余均包含部分5′非翻譯區和3′非翻譯區序列,RiACS1含有2個內含子而其余均含有3個內含子,內含子的長度不等,最短的僅85 bp而最長的達713 bp,去除內含子后得到的CDS序列長度在1 392~1 608 bp之間。
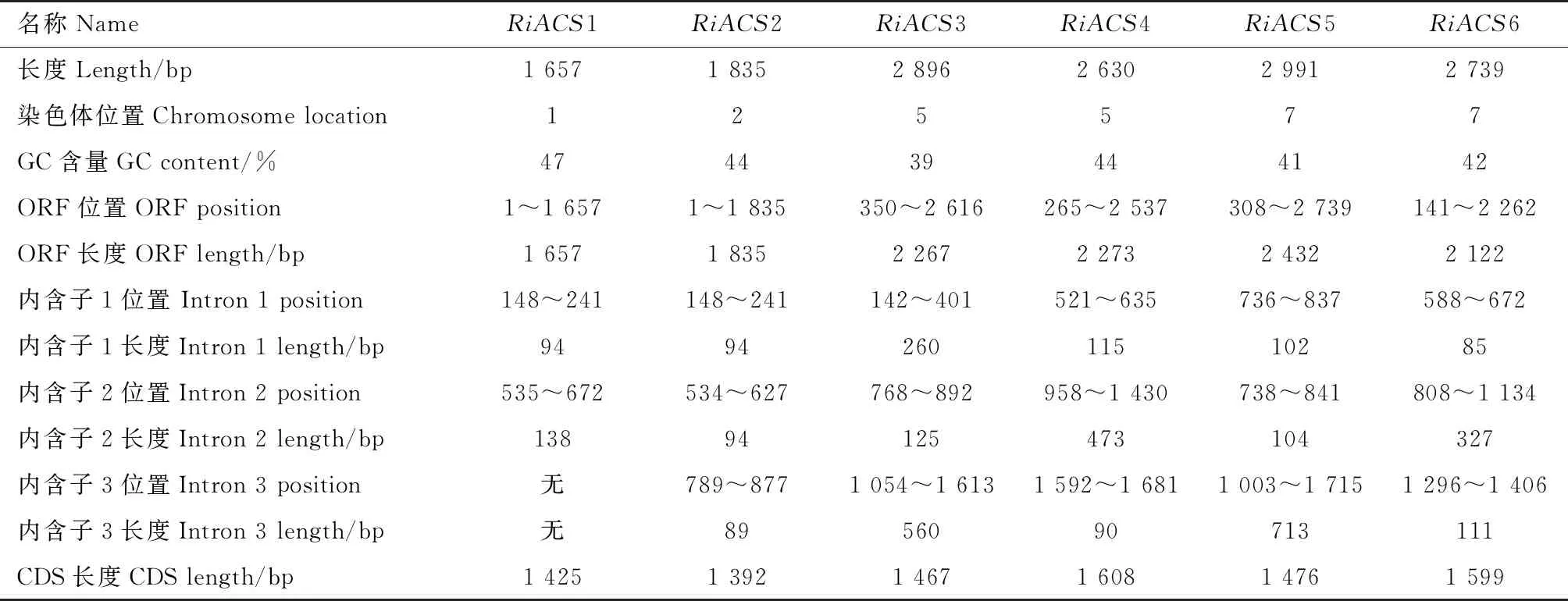
表2 樹莓ACS基因家族成員序列分析Tab.2 Sequence analysis of raspberry ACS family genes
使用DNAMAN軟件對所得的基因序列和翻譯而成的蛋白序列進行比對,結果顯示樹莓ACS家族成員之間基因序列相似性為39%~70%,蛋白序列相似性為29%~68%,這一結果與擬南芥中的研究結果一致[20],說明ACS家族成員的序列之間具有較大的差異性。
2.2 樹莓ACS家族蛋白理化性質分析
ExpPASYProtParam的蛋白理化性質分析結果表明,RiACS1-6的理論等電點集中在6.05~8.95;RiACS3和RiACS6正電荷殘基數小于負電荷殘基數,整體帶負電,其余整體帶正電;RiACS1-6不穩定系數均大于40,可能不穩定,總平均疏水性均小于0,可能是親水性蛋白[22]。
2.3 樹莓ACS家族蛋白信號肽、跨膜結構及亞細胞定位預測
SignalP的信號肽預測結果顯示樹莓ACS家族蛋白均不含有信號肽,為非分泌蛋白。TMHMMServer的跨膜結構預測結果顯示RiACS4在第33—57個氨基酸位置具有一個跨膜結構域,預測其為跨膜蛋白,其余蛋白都不含跨膜結構域,為非跨膜蛋白,但RiACS6有部分氨基酸在膜內。PSORT的亞細胞定位預測結果表明RiACS1定位于細胞核的概率最大,為76.0%,RiACS2定位于細胞核的概率最大,為88.0%,RiACS3定位于細胞核的概率最大,為60.0%,RiACS4定位于內質網膜的概率最大,為60.0%,RiACS5定位于細胞質的概率最大,為65.0%,RiACS6定位于高爾基體的概率最大,為90.0%。
2.4 樹莓ACS家族蛋白二級結構預測
SOPMA的蛋白二級結構預測結果(表3)表明樹莓ACS家族蛋白的二級結構以α-螺旋和無規則卷曲為主,分別占到36.26%~43.25%和33.76%~39.63%,各蛋白成員之間二級結構組成區別不是很大。
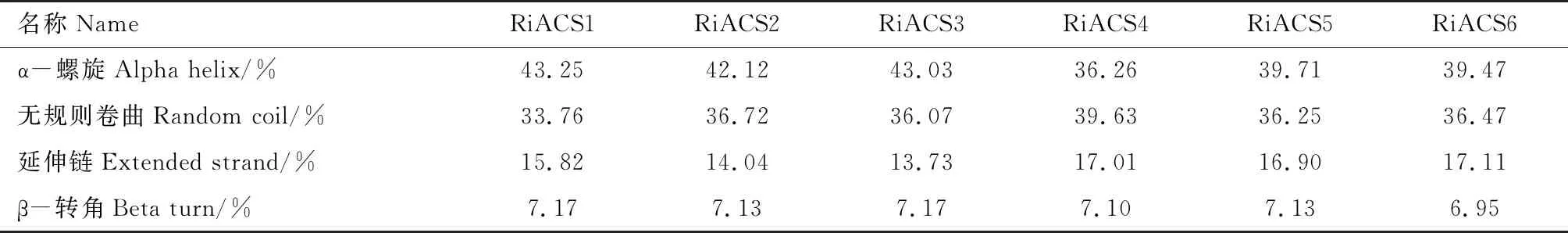
表3 樹莓ACS家族蛋白的二級結構預測Tab.3 Secondary structure prediction of raspberry ACS family proteins
2.5 樹莓ACS家族蛋白三級結構預測
將樹莓ACS家族蛋白輸入SWISS-MODEL在線建模程序,以自動匹配模板方式進行同源建模,結果顯示自動匹配到的模板均為ACS蛋白,GMQE值均在0.6~1.0之間,QMEAN值均大于0,覆蓋度均較高,說明建模結果較為可靠。ACS蛋白以同源二聚體或異源二聚體的形式行使蛋白功能[29],建模結果預測RiACS1-6蛋白均以同源二聚體的形式存在,符合預期。由三維結構圖(圖2)可以看出,RiACS1-6蛋白均以α-螺旋和無規則卷曲為主,與二級結構預測結果一致;RiACS1、RiACS2、RiACS4、RiACS6三級結構相近,而RiACS3和RiACS5三級結構相近。
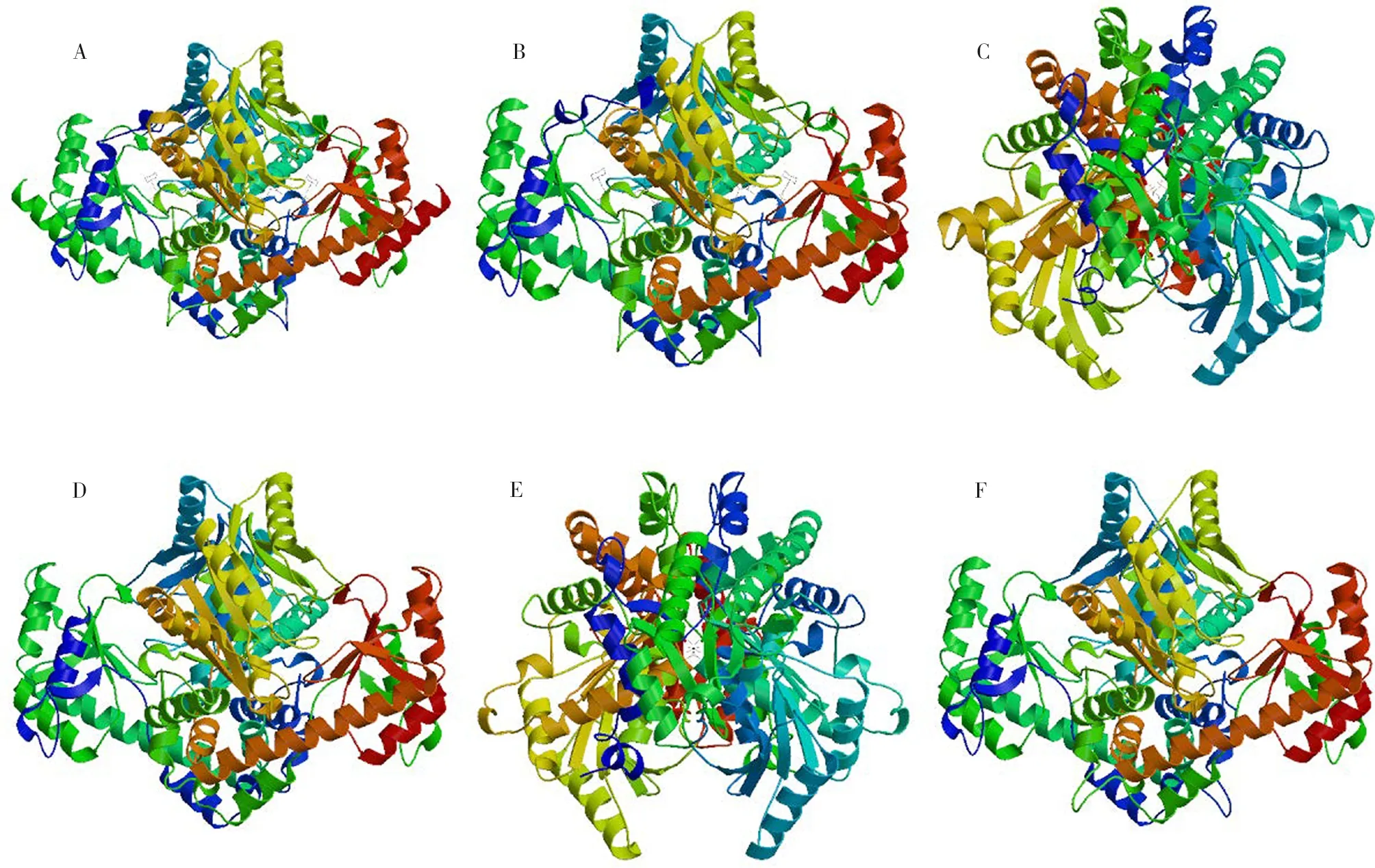
注:A-F RiACS1-6三維結構預測。Note: A-F three-dimensional structure prediction of RiACS1-6.圖2 樹莓ACS家族蛋白的三維構預測Fig.2 Three-dimession prediction of raspberry ACS family proteins
2.6 樹莓ACS家族蛋白同源性分析及進化樹構建
使用NCBI的blastp工具對樹莓ACS家族蛋白進行同源性分析,發現其與其他薔薇科植物的ACS蛋白序列均具有較高的相似性,其中與月季ACS蛋白序列相似性最高,RiACS1與月季ACS3序列相似性為91%,RiACS2與月季ACS1序列相似性為92%,RiACS5與月季ACS2序列相似性為89%。使用DNAMAN軟件對樹莓ACS家族蛋白和月季ACS家族蛋白進行多序列比對發現,RiACS1-6均含有ACS蛋白典型的7個保守結構域,而RiACS4和RiACS6在第6個結構域處與其他蛋白成員差異較大,這一結果與擬南芥中ACS10和ACS12的特點類似[20](圖3)。
將樹莓ACS家族蛋白序列和擬南芥ACS家族蛋白序列導入MEGA軟件,進行Clustalw 比對后使用鄰接法(NJ)生成系統進化樹,bootstrap值設為1000(圖4)。擬南芥ACS家族蛋白根據C末端序列差異可以分為三個類型:Type I是ACS1、ACS2和ACS6;Type II是ACS4、ACS5、ACS8和ACS9;Type III是ACS7、ACS11,此外AtACS10和AtACS12執行氨基轉移酶功能[12,20,30-31]。從進化樹可以看出,樹莓ACS家族蛋白可分為3組,RiACS3、RiACS5與AtACS1、AtACS2、AtACS6聚集于同一分支,可能是Type Ⅰ型ACS蛋白;RiACS1、RiACS2與AtACS4、AtACS5、AtACS8、AtACS9聚集于同一分支,可能是Type Ⅱ型ACS蛋白;RiACS4、RiACS6與AtACS10、AtACS12聚集于同一分支,可能執行氨基轉移酶功能。
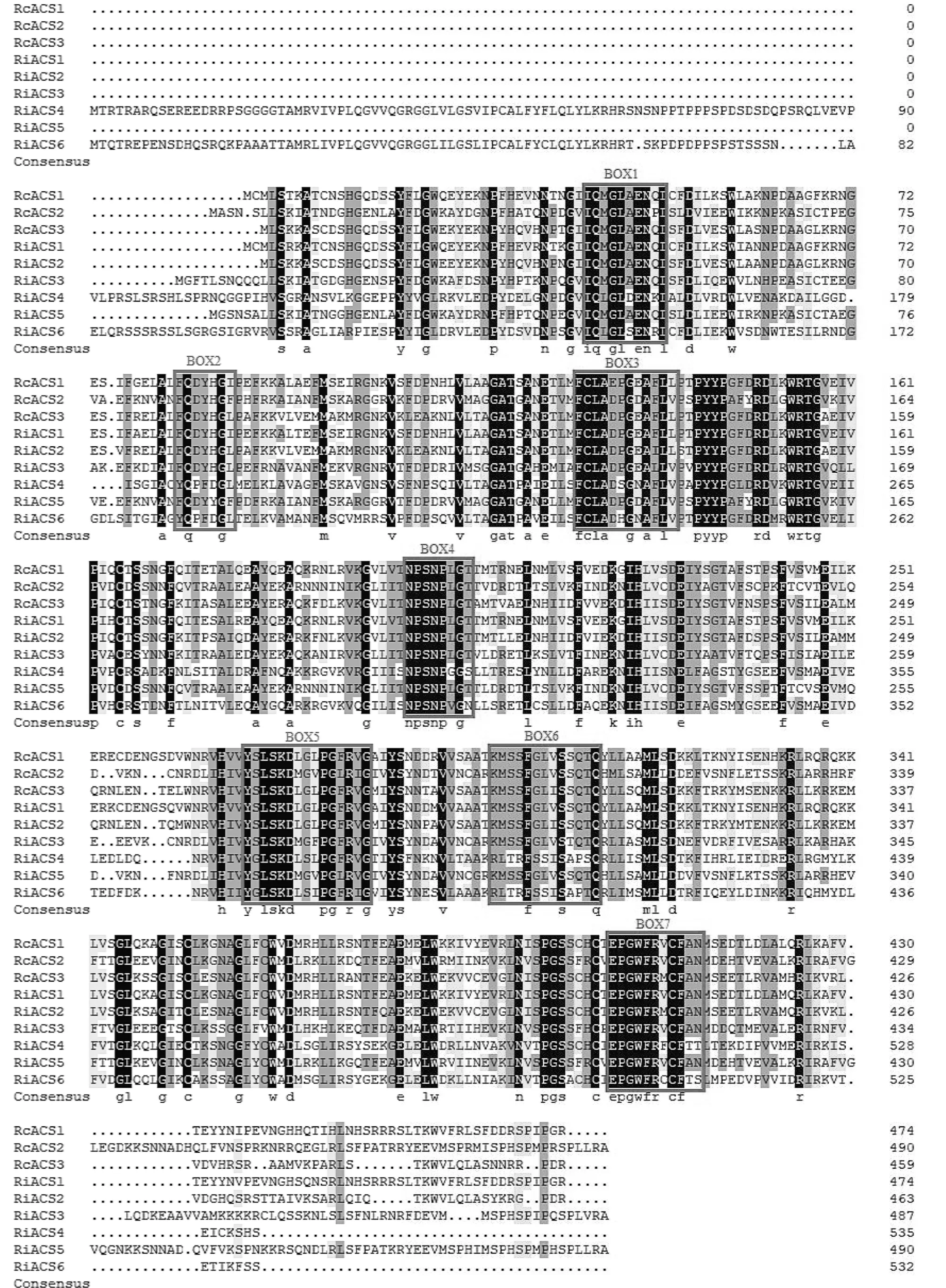
圖3 樹莓ACS家族蛋白與月季ACS蛋白多序列比對Fig.3 Multiple sequence alignment of raspberry ACS family proteins with rose ACS protein
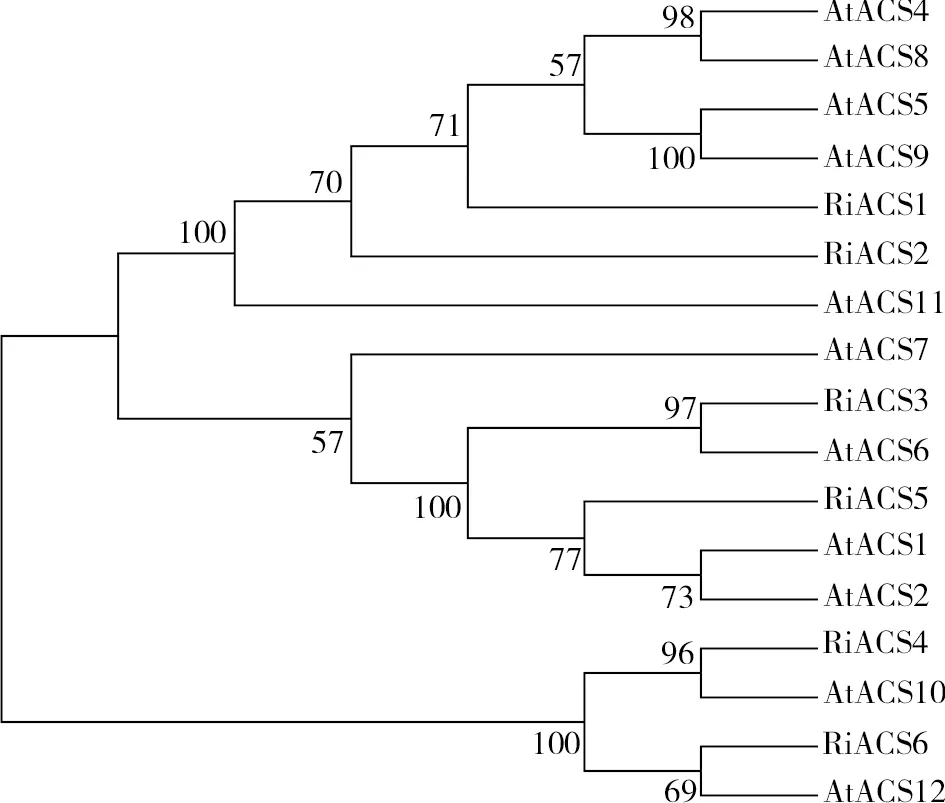
圖4 樹莓ACS家族蛋白進化分析Fig.4 Phylogenetic analysis of raspberry ACS family proteins
3 討 論
樹莓是一種廣受歡迎的新興水果,具有較高的營養價值和經濟價值,然而樹莓果實極不耐貯存的特點阻礙了樹莓產業的快速發展。乙烯在植物生長發育過程中起重要調控作用,根據成熟衰老過程乙烯釋放和呼吸強度變化的不同,果實可分為躍變型和非躍變型兩類,樹莓果實屬于非躍變型。20世紀乙烯被認為在非躍變型果實成熟衰老中發揮的作用非常有限,然而近年來越來越多的研究結果表明,非躍變型果實的成熟衰老過程也受乙烯調控[32-34]。目前有研究表明乙烯參與樹莓果實成熟進程[35],然而其分子機理仍不清楚。乙烯合成的關鍵酶是ACS,因此,進行樹莓ACS基因的克隆和分析有助于推進樹莓果實成熟機制的研究,為改善樹莓果實不耐貯存的特性提供理論基礎。
隨著基因組計劃的進行,越來越多的物種完成了全基因組測序。與此同時,其中很多物種的EST資料卻沒有大規模的增長。傳統的電子克隆利用EST數據庫來獲取電子序列,在EST資料不夠豐富的物種中無法應用。本試驗使用蛋白保守結構域序列與基因組數據庫比對的方法進行電子克隆,避開EST數據庫的限制。試驗結果獲得6個樹莓ACS家族成員的基因序列和其蛋白序列,并進行生物信息學分析。值得一提的是,在電子克隆過程中,發現推測的第7條ACS基因(數據未顯示),其特點是基因分為前后兩段,兩段分別比對都具有ACS基因特征。在基因兩端設計特異性引物以基因組DNA為模板進行PCR擴增,得到1條長度遠小于原本電子序列的基因片段,使用Augustus程序進行內含子分析發現其不含內含子,翻譯后的蛋白序列經同源性分析屬于ACS蛋白片段。由此可以推測樹莓中存在第7條ACS基因,其電子序列由前后兩段ACS基因構成的特點有可能是測序錯誤導致的。一般意義上的基因全長是指包含5′非翻譯區、ORF和3′非翻譯區的cDNA序列,現在的生物信息學手段無法通過基因組序列準確預測mRNA的5′非翻譯區和3′非翻譯區序列,所以雖然從基因組克隆到RiACS3、RiACS4、RiACS5、RiACS6的ORF區兩端非翻譯區序列,但仍無法確定其是否為基因全長,后續試驗可以借助RT-PCR技術和RACE(rapid-amplification of cDNA ends)技術來獲取包括第7條ACS基因在內的樹莓ACS多基因家族成員的cDNA全長序列。
對本試驗獲得的樹莓ACS家族基因序列和蛋白序列進行相似性比較,發現其成員之間具有較大的序列差異性。此外,盡管屬于同家族基因,但其成員之間內含子數目和長度都不盡相同,其中RiACS1包含2個較短的內含子,RiACS3和RiACS5則含有兩短一長的3個內含子,而RiACS4和RiACS6含有短-長-短3個內含子,這一發現也從側面證明ACS多基因家族成員之間的序列差異性。樹莓ACS家族蛋白的生物信息學預測和分析表明其均為不穩定的親水性蛋白,均不含信號肽。RiACS1、RiACS2和RiACS3可能是位于細胞核的非跨膜蛋白,RiACS4可能是位于內質網膜的跨膜蛋白,RiACS5可能是位于細胞質的非跨膜蛋白,RiACS6可能是位于高爾基體的非跨膜蛋白。一般定位于內膜系統的蛋白和跨膜蛋白均含有信號肽,但RiACS4和RiACS6的預測結果卻不含信號肽,可能是由于信號肽或亞細胞定位預測軟件結果不準確導致的。RiACS1-6的二級結構較為相似,均以α-螺旋和無規則卷曲為主,三級結構均以同源二聚體形式存在,分為RiACS3、RiACS5和RiACS1、RiACS2、RiACS4、RiACS6兩組。它們均含有7個ACS蛋白保守結構域,但RiACS4和RiACS6在第6個結構域處與其他蛋白有所差異,并且在系統進化樹中,RiACS4和RiACS6與擬南芥ACS10、ACS12聚集于同一分支,而擬南芥ACS10、ACS12是氨基轉移酶,所以推測RiACS4和RiACS6有可能是氨基轉移酶的一種而不是ACS。
研究表明,植物ACS基因的表達具有組織、器官特異性,時空特異性,且受外界環境的影響[36]。本試驗克隆到樹莓6條ACS基因家族成員,后續可以開展RT-qPCR等試驗來研究不同ACS基因家族成員在樹莓中的表達差異性,找到在樹莓果實成熟過程中特異性表達的ACS基因,為進一步研究樹莓果實成熟機制奠定基礎。
