江蘇鹽城沿海灘涂濕地放線菌多樣性及功能基因的篩選
2019-07-23 01:40:56保玉心
遵義醫科大學學報 2019年3期
鄭 潔,庹 利,2,李 偉,保玉心,2
(1.遵義醫科大學 研究生院,貴州 遵義 563099;2.遵義醫科大學 醫學與生物學研究中心 遵義市理化分析測試工程研究中心,貴州 遵義 563099)
放線菌因其次級代謝產物結構的復雜性和多樣性而具有抗感染、抗腫瘤等生物活性[1]。目前在臨床上應用的抗生素有3/4是由放線菌生產的[2]。如臨床廣泛應用的鏈霉素、阿奇霉素等[3]。隨著抗生素的廣泛使用,病原菌的耐藥性急劇增強,使得現有抗生素越來越不足以應對這種局面,因此迫切需要新型抗生素,以適應病原菌對抗生素敏感性改變的速度[4-6]。放線菌在自然界中分布廣泛,近年來,為了獲取新的放線菌資源,人們逐漸將目光投向一些特殊環境[7]。沿海灘涂是介于陸地與海洋的生態系統交錯區,受到海洋和陸地環境的雙重影響,因此灘涂土壤中可能擁有獨特的、既不同于陸地也不同于海洋環境的微生物資源[8-9]。有研究表明,灘涂沉積物中培養分離得到的細菌中有20%~30%屬于新菌種[10]。在同屬黃海區域的朝鮮半島潮間帶沉積物中,Lin等[11]分離到好氧菌株Marinimicrobiumkoreense和Marinimicrobiumagarilyticum;Bruns等[12]在德國北海潮間帶沉積物中分離到厭氧菌株Muricaudaruestringensis;Choi等[13]分離到從屬于Flavobacteriaceae的菌株Lutibacterlitoralis,這些結果表明灘涂土壤中可能具有獨特的微生物資源。江蘇鹽城沿海灘涂濕地位于我國沿海地區中東部,屬于從北亞熱帶向南暖溫帶過渡地帶,是太平洋西海岸亞洲大陸邊緣面積最大、原始生態保持最完好的海岸型天然濕地。目前,關于江蘇鹽城沿海濕地微生物特別是放線菌研究報道較少,紀元、張小蒙等只對江蘇鹽城沿海灘涂進行了簡單的放線菌菌落計數[14-15]。對該地區放線菌的多樣性及抗生素生物合成基因未見報道。本研究從江蘇鹽城沿海采集灘涂土壤樣品,通過純培養的方法對該地區的放線菌進行分離鑒定、多樣性分析及抗生素生物合成基因篩選,以了解該地區放線菌分布特點,同時期待發現放線菌新物種或稀有放線菌,為新抗生素發現提供菌種儲備,為進一步開發利用灘涂濕地放線菌資源開發和利用提供科學依據。
1 材料與方法
1.1 材料
1.1.1 土壤樣品 2018年2月,圍繞鹽城市射陽縣黃海海域每隔10~20 m設置采樣點一個(樣品采集地點見表1)。去除土壤表面覆蓋的泥沙后,用無菌鏟鏟取20 cm深度土樣后立即放入無菌封口袋中,放于4 ℃便攜冰箱中保存運回實驗室。共采集4個樣本,分別標記Y1、Y2、Y3、Y4。
表1采集的土壤樣品信息

樣品采集地點Y1N33°41′23″,E120°33′57″Y2N33°41′23″,E120°33′57″Y3N33°41′23″,E120°33′58″Y4N33°41′23″,E120°33′57″
1.1.2 培養基 ①分離培養基:A、基礎培養基:酸性酪蛋白5.0 g、檸檬酸鈉3.0 g、酵母浸粉 10.0 g、KCl 2.0 g、蛋白胨5.0 g、MgSO4·7H2O 2.0 g、NaCl 40.0 g、瓊脂20.0 g,加雙蒸水定容至1 L,pH7.2;B、Marine agar 2216E合成培養基(MA):Marine broth 2216E (MB;美國BD)37.5 g,瓊脂20.0 g,加雙蒸水定容至1 L,pH7.2;C、改良TSA合成培養基:tryptic soy broth (TSB;美國BD)30.0 g,NaCl 20.0 g,瓊脂20.0 g,加雙蒸水定容至1 L,pH7.2;D、TSA合成培養基:Tryptic Soy broth(TSB;美國BD)30 .0 g,瓊脂20.0 g,加雙蒸水定容至1 L,pH7.2;E、改良ISP2培養基:酵母粉4.0 g,麥芽浸粉10.0 g,葡萄糖4.0 g,瓊脂20.0 g,NaCl 20.0 g,加雙蒸水定容至1 L,pH7.2;F、ISP2培養基:酵母粉4.0 g,麥芽浸粉10.0 g,葡萄糖4.0 g,瓊脂20.0 g,加雙蒸水定容至1 L,pH7.2。②純化培養基:ISP2培養基:酵母粉4.0 g,麥芽浸粉4.0 g,葡萄糖4.0 g,瓊脂20.0 g,加雙蒸水定容至1 L,pH7.2。
1.1.3 抑制劑 萘啶酮酸20 mg/L,重鉻酸鉀60 mg/L[16],待培養基溫度降至40~50 ℃時加入。
1.1.4 主要試劑和儀器 2×Easy Taq SuperMix、High Fidelity PCR SuperMix、pEASY-T1 Cloning Kit、EasyPure Quick Gel Extraction Kit、Trans-T1感受態細胞均購自北京全式金生物技術有限公司;DNA Marker、X-gal均購自上海生工生物工程有限公司;2×Taq PCR Master Mix購自北京康為世紀生物科技有限公司;PCR引物由上海生工生物工程有限公司合成;Chelex-100樹脂,美國BioRad公司;Agarose、TAE緩沖液購自北京索萊寶公司;Tryptic Soy broth合成培養基和marine agar 2216E合成培養基購自美國BD公司。生物安全柜(BSC-1600IIA2),蘇州安泰空氣技術有限公司;生化培養箱(ZXSD-B1270),上海智城分析儀器制造有限公司;SHKE481HP型落地式恒溫制冷搖床,美國Thermo公司;高壓蒸汽滅菌鍋,日本Tomy;Microfuge20 系列臺式微量離心機(B31606),美國Beckman公司;電泳儀,北京市六一儀器廠;PCR擴增儀以及凝膠成像儀,美國BioRad公司。
1.2 方法
1.2.1 樣品處理 取土壤樣品置于無菌培養皿中,于菌種分離前2周自然風干研磨過篩,每個樣品取2 g置于裝有18 mL無菌水的離心管中,28℃、180rpm振蕩過夜。
1.2.2 菌株分離培養、純化及保藏 將預處理的土壤懸液進行梯度稀釋,取200 μL 10-3稀釋度樣品接種于上述分離培養基平皿中,用滅菌后的涂布棒涂勻。倒置放入28 ℃培養箱中培養4~8周,及時觀察分離培養基上的菌落形態,并挑取肉眼判斷為放線菌的單菌落進行菌株編號,采用四區劃線法接種于純化培養基平板上,重復這一過程,直到得到純菌株。得到的純培養物接種于20%甘油中-80 ℃保藏。
1.2.3 基于16S rRNA基因序列的系統發育分析 ①16S rRNA基因測序和分析:菌株DNA提取按Chelex-100法進行[17-18]。PCR擴增引物為通用引物27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和1492R(5′-GGTTACCTTGTTACGACTT-3′)。PCR反應體系:2(EasyTaqSuperMix 25 μL,27F引物2 μL,1492R引物2 μL,模板2 μL,無菌水19 μL。PCR反應條件:95 ℃ 5 min;94 ℃ 1 min,55 ℃ 1 min,72 ℃ 2 min,35個循環;72 ℃ 10 min。PCR擴增產物經瓊脂糖凝膠電泳檢測,將電泳檢測結果顯示為陽性的PCR擴增產物送交上海生工生物工程股份有限公司進行序列測定。潛在新物種的16S rRNA基因序列使用High Fidelity PCR SuperMix擴增,反應體系及條件同上;PCR擴增產物經瓊脂糖凝膠電泳檢測,目標陽性條帶切膠回收,將回收DNA片段連接到pEASY-1克隆載體后,轉化至Trans1-T1感受態細胞中;利用抗性和藍白斑篩選挑取陽性克隆子,接種于添加氨芐青霉素的LB液體培養基中,37 ℃、200 rpm培養10 h,菌液交由生工生物工程(上海)股份有限公司完成測序。②序列分析和系統發育樹構建:測序獲得的16SrRNA基因序列利用EzBioCloud(http://www.ezbiocloud.net)[19]數據庫中的EzTaxon在線比對服務進行相關標準菌株的相似性比對搜索,確定菌株的分類學地位,并從中調出相關的標準菌株的序列作為參比對象,用Clustal X[20]軟件進行多序列比對,采用MEGA 5.0[21]軟件以鄰接法(Neighbor-joining)[22]進行聚類分析并構建系統發育樹,系統進化矩陣根據Kimura-2-parameter模型估算,重復取樣1 000次進行自展值分析以評估進化樹拓撲結構的穩定性。
1.2.4 抗生素生物合成基因(NRPS、PKSI和PKSII)的PCR篩選 放線菌基因組DNA的提取方法同上,以基因組DNA作為模板PCR擴增非核糖體多肽合成酶(NRPS)基因A結構域(引物A3F:5′-GCSTACSYSATSTACACSTCSGG-3′;A7R:5′-SASGTCVCCSGTSCGGTAS-3′)[23]、擴增I型聚酮合酶(PKSI)基因KS結構域(引物K1F:5′-TSAAGTCSAACATCGGBCA-3′;M6R:5′-CGCAGGTTSCSGTACCAGTA-3′)[24]和擴增II型聚酮合酶(PKSII)基因KS結構域(引物KSα:5′-TSGCSTGCTTGGAYGCSATC-3′;KSβ:5′-TGGAANCCGCCGAABCCTCT-3′)[24],三者擴增反應條件:96 ℃ 2 min;96 ℃ 1 min,60 ℃ 2 min,73 ℃ 1.5 min,35個循環;72 ℃ 8.5 min。PCR反應體系都為50 μL,其中模板DNA 2 μL,A3F/ A7R (10 μM)、K1F/ M6R (10 μM)、KSα/ KSβ (10 μM)分別為2 μL,2(Taq PCR Master Mix為25 μL,用ddH2O補至50 μL。PCR擴增產物用1%瓊脂糖凝膠電泳進行檢測,凝膠成像儀觀察結果。
2 結果與分析
2.1 菌種分離結果 從江蘇鹽城沿海灘涂濕地收集的4份土壤樣品,采用6種分離培養基分離放線菌,根據菌落形態初步排重,選擇菌株進行16S rRNA基因序列擴增和序列分析,16S rRNA基因序列比對結果表明134株為放線菌,不同樣品中放線菌出菌數見圖1。

圖1 不同樣品中分離到的放線菌數量
不同培養基中分離的菌株具有特殊性,Curtobacterium為A培養基所分離,Microbacterium為B培養基所分離,Dietzia、Micromonospora、Paenarthrobacter、Pseudarthrobacter為C培養基所分離,Isoptericola為D培養基所分離,Mycetocola為E培養基所分離,Pseudarthrobacter為F培養基所分離。不同培養基中放線菌出菌數見表2。因此,根據分離到的菌株數目以及種屬數目,比較好的培養基依次為D培養基、C培養基、E培養基、B培養基、F培養基、A培養基。
表26種培養基分離放線菌的效果
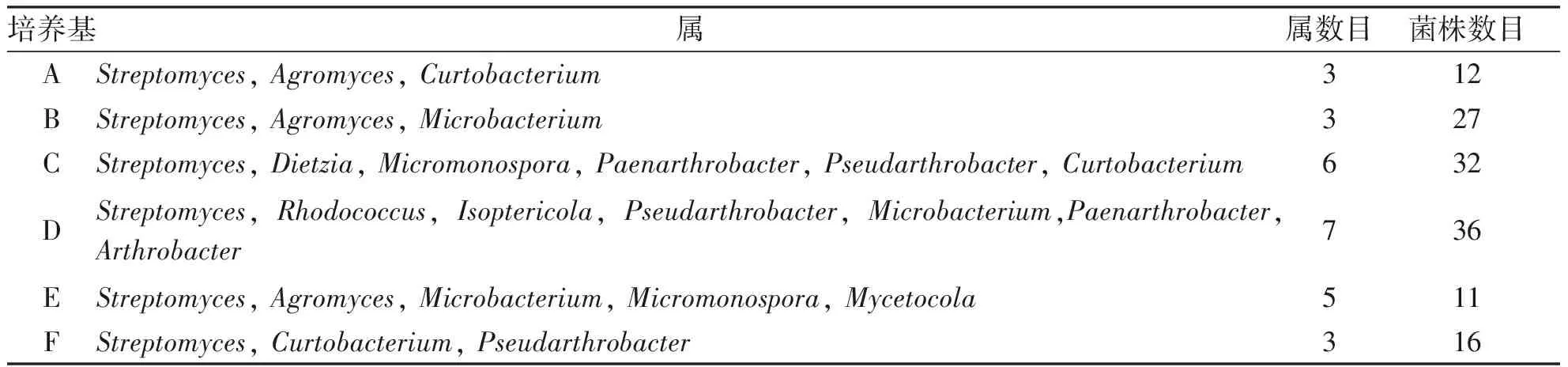
培養基屬屬數目菌株數目AStreptomyces, Agromyces, Curtobacterium312BStreptomyces, Agromyces, Microbacterium327CStreptomyces, Dietzia, Micromonospora, Paenarthrobacter, Pseudarthrobacter, Curtobacterium 632DStreptomyces, Rhodococcus, Isoptericola, Pseudarthrobacter, Microbacterium,Paenarthrobacter, Arthrobacter736EStreptomyces, Agromyces, Microbacterium, Micromonospora, Mycetocola511FStreptomyces, Curtobacterium, Pseudarthrobacter 316
2.2 土壤放線菌多樣性分析 分離到的134株放線菌分布于放線菌域的4個目7個科12個屬中,除優勢菌鏈霉菌屬(Streptomyces)外,還分離到微桿菌屬(Microbacterium)、節桿菌屬(Arthrobacter)、Paenarthrobacter、壤霉菌屬(Agromyces)、短小桿菌屬(Curtobacterium)、小單孢菌屬(Micromonospora)、迪茨氏菌屬(Dietzia)、紅球菌屬(Rhodococcus)、Pseudarthrobacter、白蟻菌屬 (Isoptericola)、棲霉菌屬(Mycetocola)稀有放線菌屬。其中,鏈霉菌屬為108株,占所分離菌株的80.6%,為優勢菌屬(見表3和圖2)。所獲得的稀有放線菌菌株以及種屬數相對較高。
表3134株放線菌的多樣性分布情況

目Order科Family屬Genus菌株數量NO. of strains鏈霉菌目Streptomycetales鏈霉菌科Sterptomycetaceae鏈霉菌屬Streptomyces108微球菌目Micrococcales微桿菌科Microbacteriaceae微桿菌屬Microbacterium4棲霉菌屬Mycetocola1壤霉菌屬 Agromyces3短小桿菌屬Curtobacterium4原小單孢菌科Promicromonosporaceae白蟻菌屬Isoptericola2微球菌科Micrococcaceae節桿菌屬Arthrobacter2Pseudarthrobacter3Paenarthrobacter3小單孢菌目Micromonosporales小單孢菌科Micromonosporaceae小單孢菌屬Micromonospora2棒桿菌目Corynebacteriales迪茨氏菌科Dietziaceae迪茨氏菌屬Dietzia1諾卡氏菌科Nocardiaceae紅球菌屬Rhodococcus1
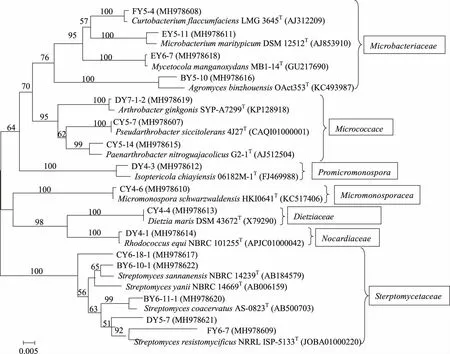
分支點上的數值為1 000次自舉值分析所得值,僅顯示大于50%的值;標尺0.005為進化距離。圖2 基于16S rRNA基因部分序列構建的部分菌株的Neighbour-Joining系統進化樹
2.3 放線菌新穎性分析 以98.65%作為區分兩個原核物種的“金標準”[25],即菌株16S rRNA基因序列最大相似率低于98.65%的視為潛在新物種,對江蘇鹽城沿海灘涂濕地放線菌進行新穎性分析,發現分離獲得的放線菌中,16S rRNA基因序列相似率低于98.65%的菌株共有1株,編號為BY5-10。對BY5-10進行16S rRNA基因克隆測序(16S rRNA基因序列長度為1490 bp),發現其與最近有效發表菌株AgromycesbinzhouensisOact353T(GenBank 登錄號:KC493987)的16S rRNA基因序列相似率為98.35%,在基于16S rRNA基因序列構建的Neighbour-Joining(NJ)系統進化樹中,菌株BY5-10在Agromyces屬中與AgromycesbinzhouensisOAct353T聚為一簇(見圖3)。基于相似率和進化樹分析,推測菌株BY5-10為微桿菌科壤霉菌屬的潛在新種。菌株EY6-7(16S rRNA基因序列長度為1483 bp)雖然與最近有效發表菌株MycetocolamanganoxydansMB1-14T(GenBank登錄號:GU217690)的16S rRNA基因序列相似率為99.14%(高于98.65%),在基于16S rRNA基因序列構建Neighbour-Joining(NJ)系統發育樹中,菌株EY6-7在Mycetocola屬中與MycetocolamiduiensisCGMCC 1.11101T聚為一簇(見圖4),但進化距離較遠。基于進化樹分析,推測菌株EY6-7為微桿菌科Mycetocola屬的潛在新種。其確切的分類地位需要進行DNA-DNA同源性分析、細胞化學成分分析、形態特征分析、生理生化特性分析等結果來判斷。
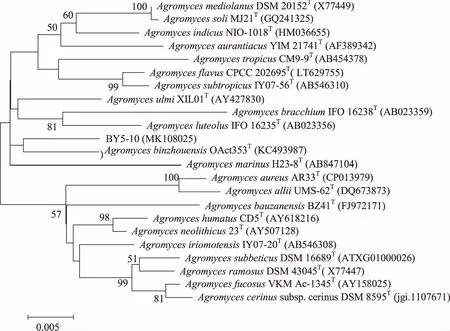
分支點上的數值為1 000次自展值分析所得值,僅顯示大于50 %的值;標尺0.005為進化距離。圖3 基于16S rRNA基因序列構建的菌株BY5-10與其相鄰菌株的Neighbour-joining系統進化樹
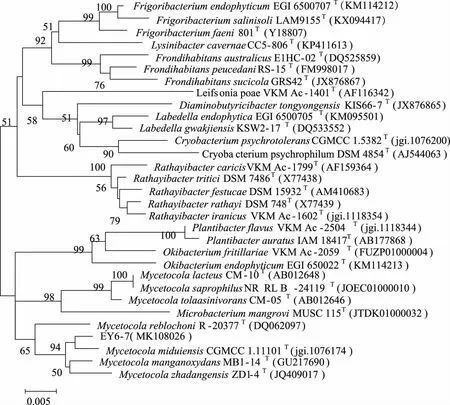
分支點上的數值為1 000次自展值分析所得值,僅顯示大于50 %的值;標尺0.005為進化距離。圖4 基于16S rRNA基因序列構建的菌株EY6-7與其相鄰菌株的Neighbour-joining系統進化樹
2.4 NRPS、PKS I和PKS II基因的PCR篩選結果 根據肉眼觀察形態不同菌株中選取63株放線菌進行NRPS、PKS I和PKS II生物合成基因的PCR篩選。3種基因片段長度不同,NRPS基因片段長度在700~800 bp之間,PKS I基因片段長度在1 200~1 400 bp之間,PKS II基因片度長度在600 bp左右。結果顯示其中55株放線菌含有至少1種生物合成基因,總陽性率為87.3%。如圖5所示,含有NRPS基因的陽性菌株數為36株,含有PKS I基因的陽性菌株數為22株,含有PKS II基因的陽性菌株數為42株,3種生物合成基因均為陽性的菌株13株。部分菌株的NRPS、PKS I和PKS II基因篩選凝膠電泳圖分別見圖6A、B、C所示。這說明江蘇鹽城灘涂地區土壤放線菌具有一定抗生素合成的潛力。

圖5 PCR篩選NRPS、PKS I和PKS II基因的陽性菌株數

圖6 功能基因篩選電泳圖
3 討論
經過多年的發掘,陸地放線菌的研究非常普遍,從陸地放線菌中發現新菌種、新活性物質的幾率正在逐漸下降[26]。而海洋放線菌由于生活在特殊的海洋環境中,其代謝途徑獨特復雜,形成的次級代謝產物在結構類型及生物活性方面都呈現出與陸地放線菌不同的特點和多樣性[27]。沿海灘涂是海陸結合帶的自然復合生態系統,兼具海洋與陸地生態系統的特點,是眾多生物的棲息地。同時,灘涂也是一種不可再生的、重要的綜合資源,具有巨大的經濟效益和生態、環境、水文、地質等價值,開發潛力很大[28]。
本研究的樣品采集自江蘇省鹽城射陽縣,射陽縣是江蘇省灘涂面積最大的縣,其沿海灘涂屬淤泥質型海岸,地勢平坦,自然屬性優越。由于本研究選取的實驗材料受沿海灘涂地區特有環境的影響,分離到的菌株具有一些海洋放線菌的特點。如分離到的鏈霉菌和小單孢菌屬是海洋沉積物中的優勢放線菌類群。
通常自然環境中只有不到1%的微生物能夠被純培養,雖然通過宏基因技術能夠獲得菌株群落結構和生物功能信息[29],但是純培養放線菌的獲得對研究了解放線菌次級代謝產物的生物合成途徑或代謝通路仍是關鍵[30]。通過預處理方式、培養基配方或共培養方式獲得難培養放線菌是當前微生物研究的重要內容之一[31]。本文通過綜合各研究人員所用培養基營養成分,設計6種分離培養基分離放線菌。共分離到到分布于3個目7個科12個屬的134株放線菌,除了優勢菌屬鏈霉菌外,還分離到微桿菌屬(Microbacterium)、節桿菌屬(Arthrobacter)、Paenarthrobacter、壤霉菌屬(Agromyces)、短小桿菌屬(Curtobacterium)、小單孢菌屬(Micromonospora)、迪茨氏菌屬(Dietzia)、Pseudarthrobacter、紅球菌屬(Rhodococcus)、Isoptericola、Mycetocola等稀有放線菌屬,所獲得的稀有放線菌菌株以及種屬數相對較高。
天然活性化合物的分離和結構解析需要花費大量的時間和資金,往往伴隨著極大的風險。因此,次級代謝產物生物合成基因篩選作為一種快速發現新型化合物的方法以及被廣泛使用,大大提高篩選效率[32-36]。因為放線菌的大部分次級代謝產物是通過NRPS和PKS途徑合成的[37],因此本研究對灘涂土壤可培養放線菌的NRPS、PKS I和PKS II等3種生物合成基因進行研究。從結果來看,63株放線菌中共有55株放線菌含有至少一種生物合成基因,總陽性率為87.3%,說明江蘇鹽城沿海灘涂濕地可培養放線菌具有較高的合成聚酮類化合物或非核糖體多肽類化合物的潛力,是后期放線菌分離方法研究的高質量材料,能為尋找新活性新結構化合物提供菌株,同時為新藥的創制奠定研究基礎。
江蘇鹽城沿海灘涂濕地中有著多樣性豐富和新穎性的放線菌資源,具有從中發現放線菌新物種及新抗生素的潛力。
